Relevant Spatial Scales of Chemical Variation in Aplysina aerophoba

Abstract
:
1. Introduction
2. Results and Discussion
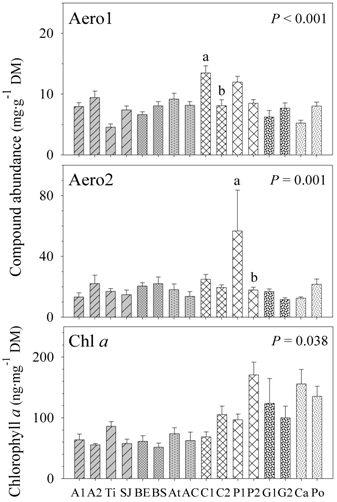
2.1. Natural Product and Chlorophyll a Quantification


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth | Chl a | All | Aero1 | Aero2 | Aply1 | Iso3 | |
| Depth | <0.001 * | 0.001 * | 0.008 | 0.276 | 0.494 | <0.001 * | |
| Chl a | −0.391 * | 0.001 * | 0.059 | 0.459 | 0.796 | <0.001 * | |
| All | 0.294 * | −0.283 * | --- | --- | --- | --- | |
| Aero1 | 0.236 | −0.168 | --- | <0.001 * | <0.001 * | <0.001 * | |
| Aero2 | 0.098 | −0.067 | --- | 0.403 * | <0.001 * | 0.001 * | |
| Aply1 | 0.062 | −0.023 | --- | 0.543 * | −0.466 * | 0.508 | |
| Iso3 | 0.349 * | −0.430 * | --- | 0.385 * | 0.294 * | 0.060 |
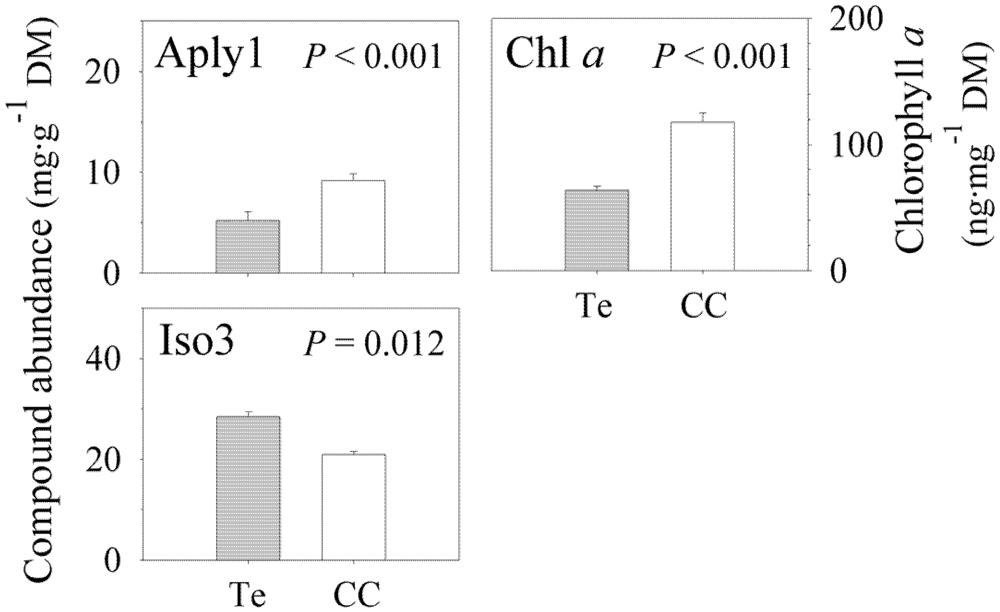
2.2. Secondary Metabolite Variation
| Compound | Depth | Region | Zone | Location | Site | Error |
|---|---|---|---|---|---|---|
| Chlorophyll a | 0.069 | <0.001/31.76% | 0.905 | 0.236 | 0.038/9.46% | 59.33% |
| Total BAs | 0.275 | 0.722 | 0.086 | 0.650 | <0.001/28.41% | 68.58% |
| Aerophobin-1 | 0.382 | 0.762 | 0.054 | 0.694 | <0.001/21.95% | 53.40% |
| Aerophobin-2 | 0.064 | 0.945 | 0.060 | 0.736 | 0.001/28.47% | 72.27% |
| Aplysinamisin-1 | 0.595 | <0.001/18.34% | 0.955 | 0.102 | 0.273 | 76.26% |
| Isofistularin-3 | 0.937 | 0.012/46.27% | 0.555 | 0.388 | 0.110 | 49.20% |


2.3. Chlorophyll a Variation
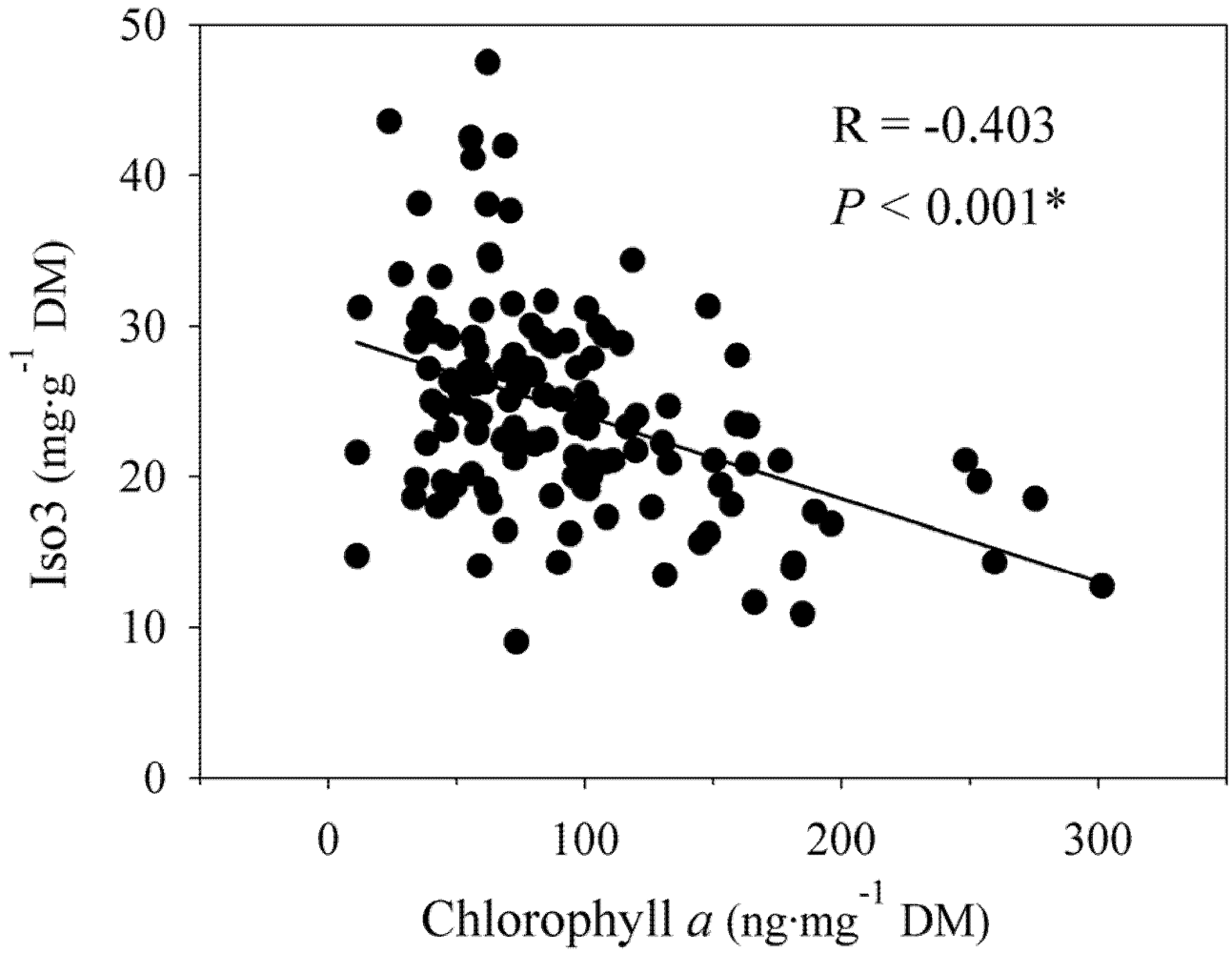
2.4. Relationship Between Natural Products and Chlorophyll a

3. Experimental Section

4. Conclusions
Acknowledgments
References
- Levin, S.A. The problem of pattern and scale in ecology. Ecology 1992, 73, 1943–1967. [Google Scholar]
- Becerro, M.A.; Turon, X.; Uriz, M.J. Multiple functions for secondary metabolites in encrusting marine invertebrates. J. Chem. Ecol. 1997, 23, 1527–1547. [Google Scholar]
- Schmitt, T.M.; Hay, M.E.; Lindquist, N. Constraints on chemically mediated coevolution—Multiple functions for seaweed secondary metabolites. Ecology 1995, 76, 107–123. [Google Scholar]
- Paul, V.J.; Vanalstyne, K.L. Activation of chemical defenses in the tropical green-algae Halimeda spp. J. Exp. Mar. Biol. Ecol. 1992, 160, 191–203. [Google Scholar]
- Matlock, D.B.; Ginsburg, D.W.; Paul, V.J. Spatial variability in secondary metabolite production by the tropical red alga Portieria hornemannii. Hydrobiologia 1999, 399, 267–273. [Google Scholar]
- Pennings, S.C.; Siska, E.L.; Bertness, M.D. Latitudinal differences in plant palatability in Atlantic coast salt marshes. Ecology 2001, 82, 1344–1359. [Google Scholar]
- Wright, J.T.; de Nys, R.; Steinberg, P.D. Geographic variation in halogenated furanones from the red alga Delisea pulchra and associated herbivores and epiphytes. Mar. Ecol. Prog. Ser. 2000, 207, 227–241. [Google Scholar]
- Hay, M.E. Marine chemical ecology: What’s known and what’s next? J. Exp. Mar. Biol. Ecol. 1996, 200, 103–134. [Google Scholar] [CrossRef]
- Paul, V.J.; Puglisi, M.P.; Ritson-Williams, R. Marine chemical ecology. Nat. Prod. Rep. 2006, 23, 153–180. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2011, 28, 196–268. [Google Scholar]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.G.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2006, 23, 26–78. [Google Scholar]
- Becerro, M.A. Quantitative trends in sponge ecology research. Mar. Ecol. Evol. Persp. 2008, 29, 167–177. [Google Scholar]
- Abdo, D.A.; Motti, C.A.; Battershill, C.N.; Harvey, E.S. Temperature and spatiotemporal variability of salicylihalamide A in the sponge Haliclona sp. J. Chem. Ecol. 2007, 33, 1635–1645. [Google Scholar]
- Duckworth, A.R.; Battershill, C.N. Population dynamics and chemical ecology of New Zealand demospongiae Latrunculia sp. nov. and Polymastia croceus (Poecilosclerida: Latrunculiidae: Polymastiidae). N. Z. J. Mar. Freshwat. Res. 2001, 35, 935–949. [Google Scholar] [CrossRef]
- Page, M.; West, L.; Northcote, P.; Battershill, C.; Kelly, M. Spatial and temporal variability of cytotoxic metabolites in populations of the New Zealand sponge Mycale hentscheli. J. Chem. Ecol. 2005, 31, 1161–1174. [Google Scholar]
- Turon, X.; Becerro, M.A.; Uriz, M.J. Seasonal patterns of toxicity in benthic invertebrates: The encrusting sponge Crambe crambe (Poecilosclerida). Oikos 1996, 75, 33–40. [Google Scholar]
- Noyer, C.; Thomas, O.P.; Becerro, M.A. Patterns of chemical diversity in the Mediterranean sponge Spongia lamella. PLoS One 2011, 6. [Google Scholar]
- Becerro, M.A.; Paul, V.J. Effects of depth and light on secondary metabolites and cyanobacterial symbionts of the sponge Dysidea granulosa. Mar. Ecol. Prog. Ser. 2004, 280, 115–128. [Google Scholar]
- Becerro, M.A.; Turon, X.; Uriz, M.J. Natural variation of toxicity in encrusting sponge Crambe crambe (Schmidt) in relation to size and environment. J. Chem. Ecol. 1995, 21, 1931–1946. [Google Scholar]
- Thompson, J.E. Exudation of biologically-active metabolites in the sponge Aplysina fistularis.1. Biological evidence. Mar. Biol. 1985, 88, 23–26. [Google Scholar]
- Ettinger-Epstein, P.; Tapiolas, D.M.; Motti, C.A.; Wright, A.D.; Battershill, C.N.; de Nys, R. Production of manoalide and its analogues by the sponge Luffariella variabilis is hardwired. Mar. Biotechnol. 2008, 10, 64–74. [Google Scholar]
- Puyana, M.; Fenical, W.; Pawlik, J.R. Are there activated chemical defenses in sponges of the genus Aplysina from the Caribbean? Mar. Ecol. Prog. Ser. 2003, 246, 127–135. [Google Scholar] [CrossRef]
- Becerro, M.A.; Paul, V.J.; Starmer, J. Intracolonial variation in chemical defenses of the sponge Cacospongia sp. and its consequences on generalist fish predators and the specialist nudibranch predator Glossodoris pallida. Mar. Ecol. Prog. Ser. 1998, 168, 187–196. [Google Scholar]
- Sacristán-Soriano, O.; Banaigs, B.; Casamayor, E.O.; Becerro, M.A. Exploring the links between natural products and bacterial assemblages in the sponge Aplysina aerophoba. Appl. Environ. Microbiol. 2011, 77, 862–870. [Google Scholar]
- Unson, M.D.; Holland, N.D.; Faulkner, D.J. A brominated secondary metabolite synthesized by the cyanobacterial symbiont of a marine sponge and accumulation of a crystalline metabolite in the sponge tissue. Mar. Biol. 1994, 119, 1–11. [Google Scholar]
- Bewley, C.A.; Holland, N.D.; Faulkner, D.J. Two classes of metabolites from Theonella swinhoei are localized in distinct populations of bacterial symbionts. Experientia 1996, 52, 716–722. [Google Scholar]
- Turon, X.; Becerro, M.A.; Uriz, M.J. Distribution of brominated compounds within the sponge Aplysina aerophoba: Coupling of X-ray microanalysis with cryofixation techniques. Cell Tissue Res. 2000, 301, 311–322. [Google Scholar]
- Bergquist, P.R.; Wells, R. Chemotaxonomy of the Porifera: The Development and Current Status of the Field. In Chemical and Biological Perspectives; Scheuer, P., Ed.; Academic Press: New York, NY, USA, 1983; Volume 5, pp. 1–50. [Google Scholar]
- Ciminiello, P.; Costantino, V.; Fattorusso, E.; Magno, S.; Mangoni, A.; Pansini, M. Chemistry of Verongida sponges. 2. Constituents of the Caribean sponge Aplysina fistularis forma fulva. J. Nat. Prod. 1994, 57, 705–712. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Magno, S.; Pansini, M. Chemistry of verongida sponges. 8. Bromocompounds from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. Tetrahedron 1997, 53, 6565–6572. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Magno, S.; Pansini, M. Chemistry of Verongida sponges. 6. Comparison of the secondary metabolic composition of Aplysina insularis and Aplysina fulva. Biochem. Syst. Ecol. 1996, 24, 105–113. [Google Scholar]
- Tymiak, A.A.; Rinehart, K.L. Biosynthesis of dibromotyrosine-derived antimicrobial compounds by the marine sponge Aplysina fistularis (Verongia aurea). J. Am. Chem. Soc. 1981, 103, 6763–6765. [Google Scholar]
- Teeyapant, R.; Kreis, P.; Wray, V.; Witte, L.; Proksch, P. Brominated secondary compounds from the marine sponge Verongia aerophoba and the sponge feeding gastropod Tylodina perversa. Z. Naturforsch. C Biosci. 1993, 48, 640–644. [Google Scholar]
- Encarnacion-Dimayuga, R.; Ramirez, M.R.; Luna-Herrera, J. Aerothionin, a bromotyrosine derivative with antimycobacterial activity from the marine sponge Aplysina gerardogreeni (Demospongia). Pharm. Biol. 2003, 41, 384–387. [Google Scholar]
- Koulman, A.; Proksch, P.; Ebel, R.; Beekman, A.C.; van Uden, W.; Konings, A.W.T.; Pedersen, J.A.; Pras, N.; Woerdenbag, H.J. Cytotoxicity and mode of action of aeroplysinin-1 and a related dienone from the sponge Aplysina aerophoba. J. Nat. Prod. 1996, 59, 591–594. [Google Scholar]
- Teeyapant, R.; Woerdenbag, H.J.; Kreis, P.; Hacker, J.; Wray, V.; Witte, L.; Proksch, P. Antibiotic and cytotoxic activity of brominated compounds from the marine sponge Verongia aerophoba. Z. Naturforsch. C Biosci. 1993, 48, 939–945. [Google Scholar]
- Weiss, B.; Ebel, R.; Elbrächter, M.; Kirchner, M.; Proksch, P. Defense metabolites from the marine sponge Verongia aerophoba. Biochem. Syst. Ecol. 1996, 24, 1–12. [Google Scholar]
- Kelly, S.R.; Jensen, P.R.; Henkel, T.P.; Fenical, W.; Pawlik, J.R. Effects of Caribbean sponge extracts on bacterial attachment. Aquat. Microb. Ecol. 2003, 31, 175–182. [Google Scholar]
- Thoms, C.; Ebel, R.; Proksch, P. Activated chemical defense in Aplysina sponges revisited. J. Chem. Ecol. 2006, 32, 97–123. [Google Scholar]
- Thoms, C.; Wolff, M.; Padmakumar, K.; Ebel, R.; Proksch, P. Chemical defense of Mediterranean sponges Aplysina cavernicola and Aplysina aerophoba. Z. Naturforsch. C Biosci. 2004, 59, 113–122. [Google Scholar]
- Nunez, C.V.; de Almelda, E.V.R.; Granato, A.C.; Marques, S.O.; Santos, K.O.; Pereira, F.R.; Macedo, M.L.; Ferreira, A.G.; Hajdu, E.; Pinheiro, U.S.; et al. Chemical variability within the marine sponge Aplysina fulva. Biochem. Syst. Ecol. 2008, 36, 283–296. [Google Scholar]
- Becerro, M.A.; Turon, X.; Uriz, M.J.; Templado, J. Can a sponge feeder be a herbivore? Tylodina perversa (Gastropoda) feeding on Aplysina aerophoba (Demospongiae). Biol. J. Linn. Soc. 2003, 78, 429–438. [Google Scholar]
- Kreuter, M.H.; Robitzki, A.; Chang, S.; Steffen, R.; Michaelis, M.; Kljajic, Z.; Bachmann, M.; Schröder, H.C.; Müller, W.E.G. Production of the cytostatic agent aeroplysinin by the sponge Verongia aerophoba in in vitro culture. Comp. Biochem. Phys. C 1992, 101, 183–187. [Google Scholar]
- Putz, A.; Kloeppel, A.; Pfannkuchen, M.; Brummer, F.; Proksch, P. Depth-related alkaloid variation in Mediterranean Aplysina sponges. Z. Naturforsch. C Biosci. 2009, 64, 279–287. [Google Scholar]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2004, 21, 519–538. [Google Scholar]
- Unson, M.D.; Faulkner, D.J. Cyanobacterial symbiont biosynthesis of chlorinated metabolites from Dysidea herbacea (Porifera). Experientia 1993, 49, 349–353. [Google Scholar]
- Siegl, A.; Hentschel, U. PKS and NRPS gene clusters from microbial symbiont cells of marine sponges by whole genome amplification. Environ. Microbiol. Rep. 2010, 2, 507–513. [Google Scholar]
- Erwin, P.M.; Thacker, R.W. Incidence and identity of photosynthetic symbionts in Caribbean coral reef sponge assemblages. J. Mar. Biol. Assoc. UK 2007, 87, 1683–1692. [Google Scholar]
- Freeman, C.J.; Thacker, R.W. Complex interactions between marine sponges and their symbiotic microbial communities. Limnol. Oceanogr. 2011, 56, 1577–1586. [Google Scholar]
- Ebel, R.; Brenzinger, M.; Kunze, A.; Gross, H.J.; Proksch, P. Wound activation of protoxins in marine sponge Aplysina aerophoba. J. Chem. Ecol. 1997, 23, 1451–1462. [Google Scholar]
- Teeyapant, R.; Proksch, P. Biotransformation of brominated compounds in the marine sponge Verongia aerophoba—Evidence for an induced chemical defense. Naturwissenschaften 1993, 80, 369–370. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research; Freeman,W. H. and Co.: New York, NY, USA, 1995; p. 887. [Google Scholar]
- Coykendall, D.K.; Johnson, S.B.; Karl, S.A.; Lutz, R.A.; Vrijenhoek, R.C. Genetic diversity and demographic instability in Riftia pachyptila tubeworms from eastern Pacific hydrothermal vents. BMC Evol. Biol. 2011, 11, 96–106. [Google Scholar]
- Noyer, C.; Becerro, M.A. Relationship between genetic, chemical, and bacterial diversity in the Atlanto-Mediterranean bath sponge Spongia lamella. Hidrobiologia 2011, in press. [Google Scholar]
- Poulin, R.; Blanar, C.A.; Thieltges, D.W.; Marcogliese, D.J. The biogeography of parasitism in sticklebacks: Distance, habitat differences and the similarity in parasite occurrence and abundance. Ecography 2011, 34, 540–551. [Google Scholar]
- Swearingen, D.C.; Pawlik, J.R. Variability in the chemical defense of the sponge Chondrilla nucula against predatory reef fishes. Mar. Biol. 1998, 131, 619–627. [Google Scholar]
- Ding, C.M.; Cantor, C.R. Quantitative analysis of nucleic acids—The last few years of progress. J. Biochem. Mol. Biol. 2004, 37, 1–10. [Google Scholar]
- Donia, M.S.; Schmidt, E.W. Linking chemistry and genetics in the growing cyanobactin natural products family. Chem. Biol. 2011, 18, 508–519. [Google Scholar]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae, and natural phytoplankton. Biochem. Physiol. Pflanz 1975, 167, 191–194. [Google Scholar]
- SPSS, Systat 9: Statistics I, 9.0 ed; SPSS Inc.: Chicago, IL, USA, 1999.
- SPSS, Systat 9: Statistics II, 9.0 ed; SPSS Inc.: Chicago, IL, USA, 1999.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sacristan-Soriano, O.; Banaigs, B.; Becerro, M.A. Relevant Spatial Scales of Chemical Variation in Aplysina aerophoba. Mar. Drugs 2011, 9, 2499-2513. https://doi.org/10.3390/md9122499
Sacristan-Soriano O, Banaigs B, Becerro MA. Relevant Spatial Scales of Chemical Variation in Aplysina aerophoba. Marine Drugs. 2011; 9(12):2499-2513. https://doi.org/10.3390/md9122499
Chicago/Turabian StyleSacristan-Soriano, Oriol, Bernard Banaigs, and Mikel A. Becerro. 2011. "Relevant Spatial Scales of Chemical Variation in Aplysina aerophoba" Marine Drugs 9, no. 12: 2499-2513. https://doi.org/10.3390/md9122499
APA StyleSacristan-Soriano, O., Banaigs, B., & Becerro, M. A. (2011). Relevant Spatial Scales of Chemical Variation in Aplysina aerophoba. Marine Drugs, 9(12), 2499-2513. https://doi.org/10.3390/md9122499