Natural Products from the Lithistida: A Review of the Literature since 2000

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction

2. Cyclic Peptides











3. Linear Peptides



4. Polyketides and Macrolides











5. Sterols, Lipids, and Fatty Acids







6. Alkaloids

7. Recent Total Synthesis of Lithistid Natural Products







8. Conclusion
Acknowledgments
References
- Bewley, C.A.; Faulkner, D.J. Lithistid sponges: Star performers or hosts to the stars. Angew. Chem. Int. Ed. 1998, 37, 2162–2178. [Google Scholar]
- Hooper, J.N.A.; van Soest, R.W.M. Systema Porifera: A Guide to the Classification of Sponges; Kluwer Academic: New York, NY, USA, 2002; Volume 1. [Google Scholar]
- van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez, B.; Hajdu, E.; Pisera, A.B.; Vacelet, J.; Manconi, R.; et al. World Porifera Database. Available online: http://www.marinespecies.org/porifera (accessed on 22 August 2011).
- Catalogue of Life: 2007 Annual Checklist: Species 2000 & ITIS Catalogue of Life Hierarchy. Available online: http://www.gbif.net (accessed on 18 August 2011).
- D’Auria, M.V.; Zampella, A.; Zollo, F. The Chemistry of Lithistid Sponge: A Spectacular Source of New Metabolites. In Bioactive Natural Products; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2002; Volume 26, Part 7, pp. 1175–1258. [Google Scholar]
- Fusetani, N.; Matsunaga, S. Bioactive sponge peptides. Chem. Rev. 1993, 93, 1793–1806. [Google Scholar]
- Andrianasolo, E.H.; Gross, H.; Goeger, D.; Musafija-Girt, M.; McPhail, K.; Leal, R.M.; Mooberry, S.L.; Gerwick, W.H. Isolation of swinholide A and related glycosylated derivatives from two field collections of marine cyanobacteria. Org. Lett. 2005, 7, 1375–1378. [Google Scholar]
- Piel, J. Metabolites from symbiotic bacteria. Nat. Prod. Rep. 2009, 26, 338–362. [Google Scholar]
- Piel, J.; Hui, D.; Wen, G.; Butzke, D.; Platzer, M.; Fusetani, N.; Matsunaga, S. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc. Natl. Acad. Sci. USA 2004, 101, 16222–16227. [Google Scholar]
- Wright, A.E. The Lithistida: Important sources of compounds useful in biomedical research. Curr. Opin. Biotechnol. 2010, 21, 801–807. [Google Scholar]
- Dictionary of Natural Products on DVDBuckingham, J. (Ed.) CRC Press Taylor and Francis Group: Boca Raton, FL USA, 2011, v. 20:1.
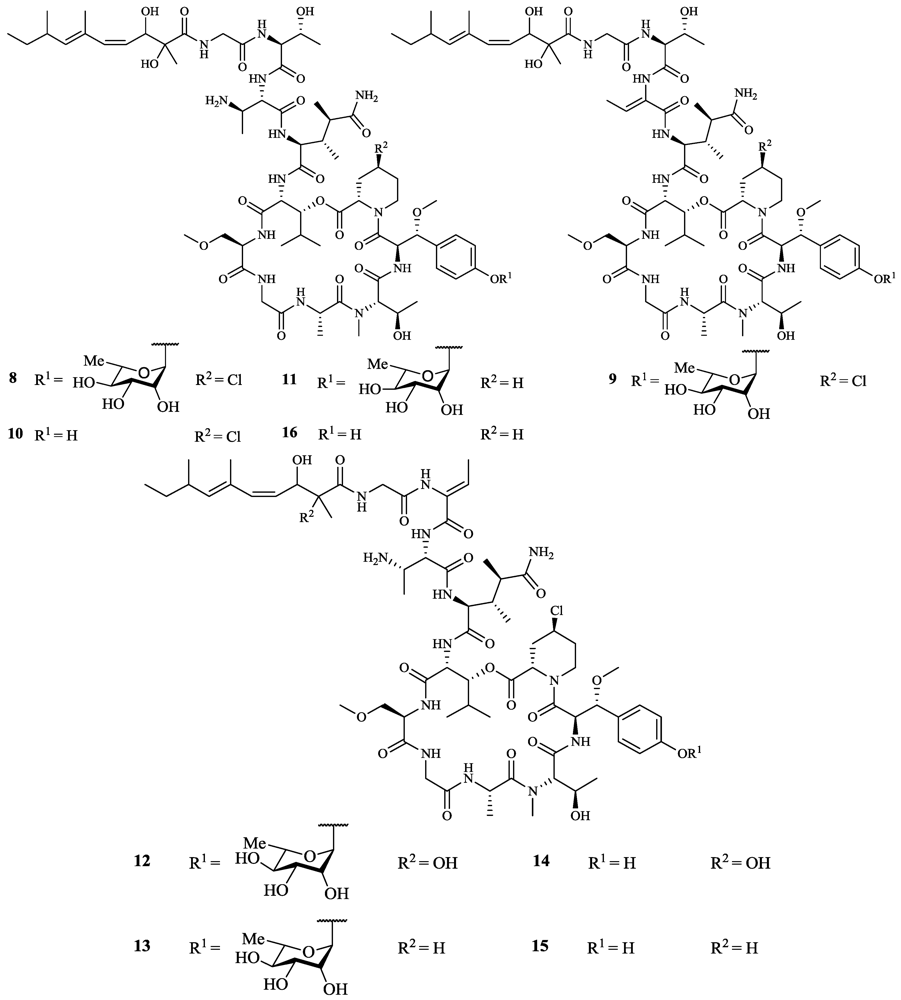
- Ratnayake, A.S.; Bugni, T.S.; Feng, X.; Harper, M.K.; Skalicky, J.J.; Mohammed, K.A.; Andjelic, C.D.; Barrows, L.R.; Ireland, C.M. Theopapuamide, a cyclic depsipeptide from a Papua New Guinea Lithistid sponge Theonella swinhoei. J. Nat. Prod. 2006, 69, 1582–1586. [Google Scholar]
- Plaza, A.; Bifulco, G.; Keffer, J.L.; Lloyd, J.R.; Baker, H.L.; Bewley, C.A. Celebesides A–C and theopapuamides B–D, depsipeptides from an Indonesian sponge that inhibit HIV-1 entry. J. Org. Chem. 2008, 74, 504–512. [Google Scholar]
- Plaza, A.; Gustchina, E.; Baker, H.L.; Kelly, M.; Bewley, C.A. Mirabamides A–D, depsipeptides from the sponge Siliquariaspongia mirabilis that inhibit HIV-1 fusion. J. Nat. Prod. 2007, 70, 1753–1760. [Google Scholar]
- Lu, Z.; van Wagoner, R.M.; Harper, M.K.; Baker, H.L.; Hooper, J.N.A.; Bewley, C.A.; Ireland, C.M. Mirabamides E–H, HIV-inhibitory depsipeptides from the sponge Stelletta clavosa. J. Nat. Prod. 2011, 74, 185–193. [Google Scholar]
- Ford, P.W.; Gustafson, K.R.; McKee, T.C.; Shigematsu, N.; Maurizi, L.K.; Pannell, L.K.; Williams, D.E.; Dilip de Silva, E.; Lassota, P.; Allen, T.M.; et al. Papuamides A–D, HIV-inhibitory and cytotoxic depsipeptides from the sponges Theonella mirabilis and Theonella swinhoei collected in Papua New Guinea. J. Am. Chem. Soc. 1999, 121, 5899–5909. [Google Scholar]
- Andjelic, C.; Planelles, V.; Barrows, L. Characterizing the anti-HIV activity of papuamide A. Mar. Drugs 2008, 6, 528–549. [Google Scholar]
- Roy, M.C.; Ohtani, I.I.; Ichiba, T.; Tanaka, J.; Satari, R.; Higa, T. New cyclic peptides from the Indonesian sponge Theonella swinhoei. Tetrahedron 2000, 56, 9079–9092. [Google Scholar]
- Roy, M.C.; Ohtani, I.I.; Tanaka, J.; Higa, T.; Satari, R. Barangamide A, a new cyclic peptide from the Indonesian sponge Theonella swinhoei. Tetrahedron Lett. 1999, 40, 5373–5376. [Google Scholar]
- Kobayashi, M.; Lee, N.K.; Shibuya, H.; Momose, T.; Kitagawa, I. Marine natural products. XXVI. Biologically active tridecapeptide lactones from the Okinawan marine sponge Theonella swinhoei (Theonellidae). Structures of theonellapeptolides Ia, Ib, Ic, and Ie. Chem. Pharm. Bull. 1991, 39, 1177–1184. [Google Scholar]
- Green, C.J. Immunosuppression with cyclosporin A: A review. Diagn. Histopathol. 1981, 4, 157–174. [Google Scholar]
- Okada, Y.; Matsunaga, S.; van Soest, R.W.M.; Fusetani, N. Nagahamide A, an antibacterial depsipeptide from the marine sponge Theonella swinhoei. Org. Lett. 2002, 4, 3039–3042. [Google Scholar]
- Bewley, C.A.; Debitus, C.; Faulkner, D.J. Microsclerodermins A and B. Antifungal cyclic peptides from the Lithistid sponge Microscleroderma sp. J. Am. Chem. Soc. 1994, 116, 7631–7636. [Google Scholar]
- Schmidt, E.W.; Faulkner, D.J. Microsclerodermins C–E, antifungal cyclic peptides from the lithistid marine sponges Theonella sp. and Microscleroderma sp. Tetrahedron 1998, 54, 3043–3056. [Google Scholar]
- Qureshi, A.; Colin, P.L.; Faulkner, D.J. Microsclerodermins F–I, antitumor and antifungal cyclic peptides from the Lithistid sponge Microscleroderma sp. Tetrahedron 2000, 56, 3679–3685. [Google Scholar]
- Erdogan, I.; Tanaka, J.I.; Higa, T. Two cyclic hexapeptides from the marine sponge Theonella cupola. FABAD J. Pharm. Sci. 2000, 25, 7–10. [Google Scholar]
- Schmidt, E.W.; Raventos-Suarez, C.; Bifano, M.; Menendez, A.T.; Fairchild, C.R.; Faulkner, D.J. Scleritodermin A, a cytotoxic cyclic peptide from the Lithistid sponge Scleritoderma nodosum. J. Nat. Prod. 2004, 67, 475–478. [Google Scholar]
- Liu, S.; Cui, Y.; Nan, F. Total synthesis of the originally proposed and revised structures of scleritodermin A. Org. Lett. 2008, 10, 3765–3768. [Google Scholar]
- Sellanes, D.; Manta, E.; Serra, G. Toward the total synthesis of scleritodermin A: Preparation of the C1-N15 fragment. Tetrahedron Lett. 2007, 48, 1827–1830. [Google Scholar]
- Angawi, R.F.; Calcinai, B.; Cerrano, C.; Dien, H.A.; Fattorusso, E.; Scala, F.; Taglialatela-Scafati, O. Dehydroconicasterol and aurantoic acid, a chlorinated polyene derivative, from the Indonesian sponge Theonella swinhoei. J. Nat. Prod. 2009, 72, 2195–2198. [Google Scholar]
- Carmely, S.; Kashman, Y. Structure of swinholide A, a new macrolide from the marine sponge. Tetrahedron Lett. 1985, 26, 511–514. [Google Scholar]
- De Silva, E.D.; Williams, D.E.; Andersen, R.J.; Klix, H.; Holmes, C.F.B.; Allen, T.M. Motuporin, a potent protein phosphatase inhibitor isolated from the Papua New Guinea sponge Theonella swinhoei. Tetrahedron Lett. 1992, 33, 1561–1564. [Google Scholar]
- Klenchin, V.A.; King, R.; Tanaka, J.; Marriott, G.; Rayment, I. Structural basis of swinholide A binding to actin. Chem. Biol. 2005, 12, 287–291. [Google Scholar]
- Maynes, J.T.; Luu, H.A.; Cherney, M.M.; Andersen, R.J.; Williams, D.; Holmes, C.F.B.; James, M.N.G. Crystal structures of protein phosphatase-1 bound to motuporin and dihydromicrocystin-LA: Elucidation of the mechanism of enzyme inhibition by cyanobacterial toxins. J. Mol. Biol. 2006, 356, 111–120. [Google Scholar]
- Wegerski, C.J.; Hammond, J.; Tenney, K.; Matainaho, T.; Crews, P. A serendipitous discovery of isomotuporin-containing sponge populations of Theonella swinhoei. J. Nat. Prod. 2006, 70, 89–94. [Google Scholar]
- Zampella, A.; Sepe, V.; Bellotta, F.; Luciano, P.; D’Auria, M.V.; Cresteil, T.; Debitus, C.; Petek, S.; Poupat, C.; Ahond, A. Homophymines B–E and A1–E1, a family of bioactive cyclodepsipeptides from the sponge Homophymia sp. Org. Biomol. Chem. 2009, 7, 4037–4044. [Google Scholar]
- Zampella, A.; Sepe, V.; Luciano, P.; Bellotta, F.; Monti, M.C.; D’Auria, M.V.; Jepsen, T.; Petek, S.; Adeline, M.-T.; Laprevote, O.; et al. Homophymine A, an anti-HIV cyclodepsipeptide from the sponge Homophymia sp. J. Org. Chem. 2008, 73, 5319–5327. [Google Scholar]
- Oku, N.; Gustafson, K.R.; Cartner, L.K.; Wilson, J.A.; Shigematsu, N.; Hess, S.; Pannell, L.K.; Boyd, M.R.; McMahon, J.B. Neamphamide A, a new HIV-inhibitory depsipeptide from the Papua New Guinea marine sponge Neamphius huxleyi. J. Nat. Prod. 2004, 67, 1407–1411. [Google Scholar]
- Zampella, A.; D’Auria, M.V.; Paloma, L.G.; Casapullo, A.; Minale, L.; Debitus, C.; Henin, Y. Callipeltin A, an anti-HIV cyclic depsipeptide from the New Caledonian Lithistida sponge Callipelta sp. J. Am. Chem. Soc. 1996, 118, 6202–6209. [Google Scholar]
- Plaza, A.; Keffer, J.L.; Lloyd, J.R.; Colin, P.L.; Bewley, C.A. Paltolides A–C, anabaenopeptin-type peptides from the Palau sponge Theonella swinhoei. J. Nat. Prod. 2010, 73, 485–488. [Google Scholar]
- Bjoerquist, P.; Buchanan, M.; Campitelli, M.; Carroll, A.; Hyde, E.; Neve, J.; Polla, M.; Quinn, R. Use of cyclic anabaenopeptin-type peptides for the treatment of a condition wherein inhibition of carboxypeptidase U is beneficial, novel anabaenopeptin derivatives and intermediates thereof. PCT Int. Appl. WO2005039617, 14 February 2008. [Google Scholar]
- Plaza, A.; Bifulco, G.; Masullo, M.; Lloyd, J.R.; Keffer, J.L.; Colin, P.L.; Hooper, J.N.A.; Bell, L.J.; Bewley, C.A. Mutremdamide A and koshikamides C–H, peptide inhibitors of HIV-1 entry from different Theonella species. J. Org. Chem. 2010, 75, 4344–4355. [Google Scholar]
- Gulavita, N.K.; Pomponi, S.A.; Wright, A.E.; Yarwood, D.; Sills, M.A. Isolation and structure elucidation of perthamide B, a novel peptide from the sponge Theonella sp. Tetrahedron Lett. 1994, 35, 6815–6818. [Google Scholar]
- Festa, C.; de Marino, S.; Sepe, V.; Monti, M.C.; Luciano, P.; D’Auria, M.V.; Débitus, C.; Bucci, M.; Vellecco, V.; Zampella, A. Perthamides C and D, two new potent anti-inflammatory cyclopeptides from a Solomon Lithistid sponge Theonella swinhoei. Tetrahedron 2009, 65, 10424–10429. [Google Scholar]
- Sepe, V.; D’Auria, M.V.; Bifulco, G.; Ummarino, R.; Zampella, A. Concise synthesis of AHMHA unit in perthamide C. Structural and stereochemical revision of perthamide C. Tetrahedron 2010, 66, 7520–7526. [Google Scholar]
- Festa, C.; de Marino, S.; Sepe, V.; D’Auria, M.V.; Bifulco, G.; Andrés, R.; Terencio, M.C.; Payá, M.; Debitus, C.; Zampella, A. Perthamides C–F, potent human antipsoriatic cyclopeptides. Tetrahedron 2011, 67, 7780–7786. [Google Scholar]
- Araki, T.; Matsunaga, S.; Nakao, Y.; Furihata, K.; West, L.; Faulkner, D.J.; Fusetani, N. Koshikamide B, a cytotoxic peptide lactone from a marine sponge Theonella sp. J. Org. Chem. 2008, 73, 7889–7894. [Google Scholar]
- Festa, C.; de Marino, S.; Sepe, V.; D’Auria, M.V.; Bifulco, G.; Debitus, C.; Bucci, M.; Vellecco, V.; Zampella, A. Solomonamides A and B, new anti-inflammatory peptides from Theonella swinhoei. Org. Lett. 2011, 13, 1532–1535. [Google Scholar]
- Nakao, Y.; Fujita, M.; Warabi, K.; Matsunaga, S.; Fusetani, N. Miraziridine A, a novel cysteine protease inhibitor from the marine sponge Theonella aff. mirabilis. J. Am. Chem. Soc. 2000, 122, 10462–10463. [Google Scholar]
- Konno, H.; Kubo, K.; Makabe, H.; Toshiro, E.; Hinoda, N.; Nosaka, K.; Akaji, K. Total synthesis of miraziridine A and identification of its major reaction site for cathepsin B. Tetrahedron 2007, 63, 9502–9513. [Google Scholar]
- Schaschke, N. Miraziridine A: Natures blueprint towards protease class-spanning inhibitors. Bioorg. Med. Chem. Lett. 2004, 14, 855–857. [Google Scholar]
- Araki, T.; Matsunaga, S.; Fusetani, N. Koshikamide A2, a cytotoxic linear undecapeptide isolated from a marine sponge of Theonella sp. Biosci. Biotechnol. Biochem. 2005, 69, 1318–1322. [Google Scholar]
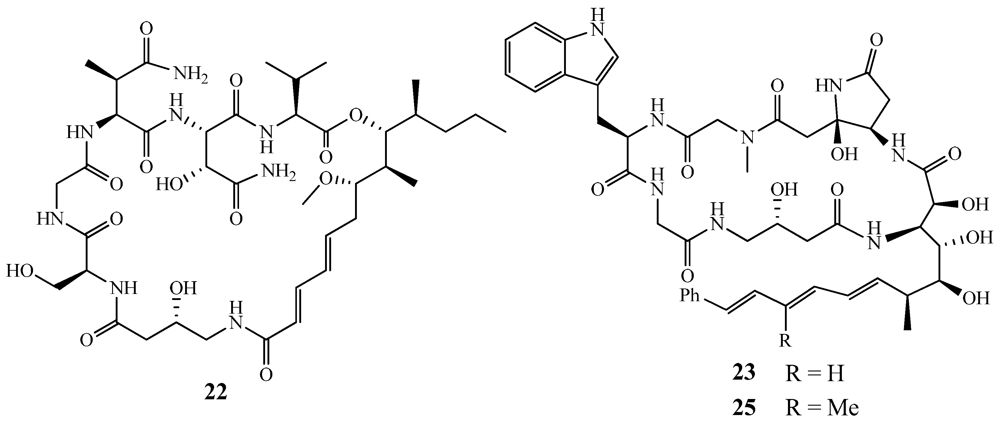
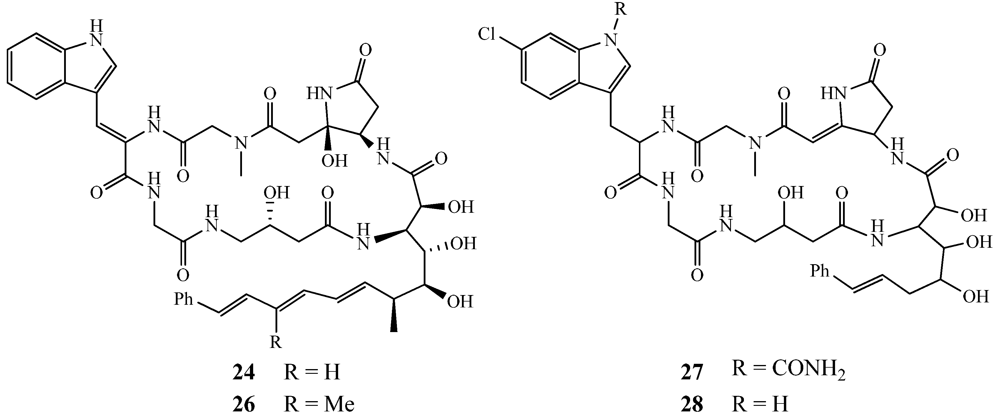
- Hamada, T.; Matsunaga, S.; Yano, G.; Fusetani, N. Polytheonamides A and B, highly cytotoxic, linear polypeptides with unprecedented structural features, from the marine sponge, Theonella swinhoei. J. Am. Chem. Soc. 2004, 127, 110–118. [Google Scholar]
- Hamada, T.; Sugawara, T.; Matsunaga, S.; Fusetani, N. Polytheonamides, unprecedented highly cytotoxic polypeptides from the marine sponge Theonella swinhoei. Structure elucidation. Tetrahedron Lett. 1994, 35, 609–612. [Google Scholar]
- Ketchem, R.R.; Lee, K.C.; Huo, S.; Cross, T.A. Macromolecular structural elucidation with solid-state NMR-derived orientational constraints. J. Biomol. NMR 1996, 8, 1–14. [Google Scholar]
- Youssef, D.T.A.; Mooberry, S.L. Hurghadolide A and swinholide I, potent actin-microfilament disrupters from the Red Sea sponge Theonella swinhoei. J. Nat. Prod. 2006, 69, 154–157. [Google Scholar]
- Kobayashi, M.; Tanaka, J.; Katori, T.; Yamashita, M.; Kitagawa, I. Marine natural products. XXII.: The absolute stereostructure of swinholide A, a potent cytotoxic dimeric macrolide from the Okinawan marine sponge Theonella swinhoei. Chem. Pharm. Bull. 1990, 38, 2409–2418. [Google Scholar]
- Marino, S.D.; Festa, C.; D’Auria, M.V.; Cresteil, T.; Debitus, C.; Zampella, A. Swinholide J, a potent cytotoxin from the marine sponge Theonella swinhoei. Mar. Drugs 2011, 9, 1133–1141. [Google Scholar]
- Edrada, R.A.; Ebel, R.; Supriyono, A.; Wray, V.; Schupp, P.; Steube, K.; van Soest, R.; Proksch, P. Swinhoeiamide A, a new highly active calyculin derivative from the marine sponge Theonella swinhoei. J. Nat. Prod. 2002, 65, 1168–1172. [Google Scholar]
- Matsunaga, S.; Wakimoto, T.; Fusetani, N. Isolation of four new calyculins from the marine sponge Discodermia calyx. J. Org. Chem. 1997, 62, 2640–2642. [Google Scholar]
- Wakimoto, T.; Matsunaga, S.; Takai, A.; Fusetani, N. Insight into binding of calyculin A to protein phosphatase 1: Isolation of hemicalyculin A and chemical transformation of calyculin A. Chem. Biol. 2002, 9, 309–319. [Google Scholar]
- Bagu, J.R.; Sykes, B.D.; Craig, M.M.; Holmes, C.F.B. A molecular basis for different interactions of marine toxins with protein phosphatase-1. J. Biol. Chem. 1997, 272, 5087–5097. [Google Scholar]
- Kita, A.; Matsunaga, S.; Takai, A.; Kataiwa, H.; Wakimoto, T.; Fusetani, N.; Isobe, M.; Miki, K. Crystal structure of the complex between calyculin A and the catalytic subunit of protein phosphatase 1. Structure 2002, 10, 715–724. [Google Scholar]
- Lindvall, M.K.; Pihko, P.M.; Koskinen, A.M.P. The binding mode of calyculin A to protein phosphatase-1. J. Biol. Chem. 1997, 272, 23312–23316. [Google Scholar]
- Sirirath, S.; Tanaka, J.; Ohtani, I.I.; Ichiba, T.; Rachmat, R.; Ueda, K.; Usui, T.; Osada, H.; Higa, T. Bitungolides A–F, new polyketides from the Indonesian sponge Theonella cf. swinhoei. J. Nat. Prod. 2002, 65, 1820–1823. [Google Scholar]
- Kobayashi, S.; Tsuchiya, K.; Kurokawa, T.; Nakagawa, T.; Shimada, N.; Iitaka, Y. Pironetin, a novel plant growth regulator produced by Streptomyces sp. NK10958. II. Structure elucidation. J. Antibiot. 1994, 47, 703–707. [Google Scholar]
- Kondoh, M.; Usui, T.; Kobayashi, S.; Tsuchiya, K.; Nishikawa, K.; Nishikiori, T.; Mayumi, T.; Osada, H. Cell cycle arrest and antitumor activity of pironetin and its derivatives. Cancer Lett. 1998, 126, 29–32. [Google Scholar]
- Su, Y.; Xu, Y.; Han, J.; Zheng, J.; Qi, J.; Jiang, T.; Pan, X.; She, X. Total Synthesis of (−)-Bitungolide F. J. Org. Chem. 2009, 74, 2743–2749. [Google Scholar]
- Sandler, J.S.; Colin, P.L.; Kelly, M.; Fenical, W. Cytotoxic macrolides from a new species of the deep-water marine sponge Leiodermatium. J. Org. Chem. 2006, 71, 7245–7251. [Google Scholar]
- Larivee, A.; Unger, J.B.; Thomas, M.; Wirtz, C.; Dubost, C.; Handa, S.; Fürstner, A. The leiodolide B puzzle. Angew. Chem. Int. Ed. 2011, 50, 304–309. [Google Scholar]
- Paterson, I.; Dalby, S.M.; Roberts, J.C.; Naylor, G.J.; Guzman, E.A.; Isbrucker, R.; Pitts, T.P.; Linley, P.; Divlianska, D.; Reed, J.K.; et al. Leiodermatolide, a potent antimitotic macrolide from the marine sponge Leiodermatium sp. Angew. Chem. Int. Ed. 2011, 50, 3219–3223. [Google Scholar]
- Gunasekera, S.P.; Paul, G.K.; Longley, R.E.; Isbrucker, R.A.; Pomponi, S.A. Five new discodermolide analogues from the marine sponge Discodermia sp. J. Nat. Prod. 2002, 65, 1643–1648. [Google Scholar]
- Gunasekera, S.P.; Gunasekera, M.; Longley, R.E.; Schulte, G.K. Discodermolide: A new bioactive polyhydroxylated lactone from the marine sponge Discodermia dissoluta. J. Org. Chem. 1990, 55, 4912–4915. [Google Scholar]
- Gunasekera, S.P.; Longley, R.E.; Isbrucker, R.A. Acetylated analogues of the microtubule-stabilizing agent discodermolide: Preparation and biological activity. J. Nat. Prod. 2001, 64, 171–174. [Google Scholar]
- Isbrucker, R.; Gunasekera, S.; Longley, R. Structure-activity relationship studies of discodermolide and its semisynthetic acetylated analogs on microtubule function and cytotoxicity. Cancer Chemother. Pharmacol. 2001, 48, 29–36. [Google Scholar]
- Wright, A.E.; Botelho, J.C.; Guzman, E.; Harmody, D.; Linley, P.; McCarthy, P.J.; Pitts, T.P.; Pomponi, S.A.; Reed, J.K. Neopeltolide, a macrolide from a Lithistid sponge of the Family Neopeltidae. J. Nat. Prod. 2007, 70, 412–416. [Google Scholar]
- Ulanovskaya, O.A.; Janjic, J.; Suzuki, M.; Sabharwal, S.S.; Schumacker, P.T.; Kron, S.J.; Kozmin, S.A. Synthesis enables identification of the cellular target of leucascandrolide A and neopeltolide. Nat Chem. Biol. 2008, 4, 418–424. [Google Scholar]
- Youngsaye, W.; Lowe, J.T.; Pohlki, F.; Ralifo, P.; Panek, J.S. Total synthesis and stereochemical reassignment of (+)-neopeltolide. Angew. Chem. Int. Ed. 2007, 46, 9211–9214. [Google Scholar]
- Custar, D.W.; Zabawa, T.P.; Scheidt, K.A. Total synthesis and structural revision of the marine macrolide neopeltolide. J. Am. Chem. Soc. 2008, 130, 804–805. [Google Scholar]
- Plaza, A.; Baker, H.L.; Bewley, C.A. Mirabilin, an antitumor macrolide lactam from the marine sponge Siliquariaspongia mirabilis. J. Nat. Prod. 2008, 71, 473–477. [Google Scholar]
- Plaza, A.; Baker, H.L.; Bewley, C.A. Mirabalin, an antitumor macrolide lactam from the marine sponge Siliquariaspongia mirabilis. J. Nat. Prod. 2009, 72, 324–324. [Google Scholar]
- Isbrucker, R.A.; Cummins, J.; Pomponi, S.A.; Longley, R.E.; Wright, A.E. Tubulin polymerizing activity of dictyostatin-1, a polyketide of marine sponge origin. Biochem. Pharmacol. 2003, 66, 75–82. [Google Scholar]
- Pettit, G.R.; Cichacz, Z.A.; Gao, F.; Boyd, M.R.; Schmidt, J.M. Isolation and structure of the cancer cell growth inhibitor dictyostatin-1. Chem. Commun. 1994, 1111–1112. [Google Scholar]
- Paterson, I.; Gardner, N.M.; Guzmán, E.; Wright, A.E. Total synthesis and biological evaluation of novel C2-C6 region analogues of dictyostatin. Bioorg. Med. Chem. 2009, 17, 2282–2289. [Google Scholar]
- Florence, G.J.; Gardner, N.M.; Paterson, I. Development of practical syntheses of the marine anticancer agents discodermolide and dictyostatin. Nat. Prod. Rep. 2008, 25, 342–375. [Google Scholar]
- Paterson, I.; Britton, R.; Delgado, O.; Meyer, A.; Poullennec, K.G. Total synthesis and configurational assignment of (−)-dictyostatin, a microtubule-stabilizing macrolide of marine sponge origin. Angew. Chem. Int. Ed. 2004, 43, 4629–4633. [Google Scholar]
- Paterson, I.; Naylor, G.J.; Fujita, T.; Guzman, E.; Wright, A.E. Total synthesis of a library of designed hybrids of the microtubule-stabilising anticancer agents taxol, discodermolide and dictyostatin. Chem. Commun. (Camb.) 2010, 46, 261–263. [Google Scholar]
- Paterson, I.; Naylor, G.J.; Gardner, N.M.; Guzman, E.; Wright, A.E. Total synthesis and biological evaluation of a series of macrocyclic hybrids and analogues of the antimitotic natural products dictyostatin, discodermolide, and taxol. Chem. Asian J. 2011, 6, 459–473. [Google Scholar]
- Paterson, I.; Naylor, G.J.; Wright, A.E. Total synthesis of a potent hybrid of the anticancer natural products dictyostatin and discodermolide. Chem. Commun. (Camb.) 2008, 4628–4630. [Google Scholar]
- Shin, Y.; Choy, N.; Balachandran, R.; Madiraju, C.; Day, B.W.; Curran, D.P. Discodermolide/dictyostatin hybrids: Synthesis and biological evaluation. Org. Lett. 2002, 4, 4443–4446. [Google Scholar]
- Shin, Y.; Fournier, J.-H.; Fukui, Y.; Brückner, A.M.; Curran, D.P. Total synthesis of (−)-dictyostatin: Confirmation of relative and absolute configurations. Angew. Chem. Int. Ed. 2004, 43, 4634–4637. [Google Scholar]
- Tsukamoto, S.; Matsunaga, S.; Fusetani, N.; Toh-e, A. Theopederins F–J: Five new antifungal and cytotoxic metabolites from the marine sponge, Theonella swinhoei. Tetrahedron 1999, 55, 13697–13702. [Google Scholar]
- Paul, G.K.; Gunasekera, S.P.; Longley, R.E.; Pomponi, S.A. Theopederins K and L. Highly potent cytotoxic metabolites from a marine sponge Discodermia sp. J. Nat. Prod. 2001, 65, 59–61. [Google Scholar]
- Fusetani, N.; Sugawara, T.; Matsunaga, S. Bioactive marine metabolites. 41. Theopederins A–E, potent antitumor metabolites from a marine sponge, Theonella sp. J. Org. Chem. 1992, 57, 3828–3832. [Google Scholar]
- Perry, N.B.; Blunt, J.W.; Munro, M.H.G.; Pannell, L.K. Mycalamide A, an antiviral compound from a New Zealand sponge of the genus Mycale. J. Am. Chem. Soc. 1988, 110, 4850–4851. [Google Scholar]
- Miyata, Y.; Matsunaga, S. Structure elucidation of 21,22-dihydroxyonnamides A1–A4 from the marine sponge Theonella swinhoei: An empirical rule to assign the relative stereochemistry of linear 1,5-diols. Tetrahedron Lett. 2008, 49, 6334–6336. [Google Scholar]
- Matsunaga, S.; Nishimura, S.; Fusetani, N. Two new antimicrobial Lysoplasmanylinositols from the marine sponge Theonella swinhoei. J. Nat. Prod. 2001, 64, 816–818. [Google Scholar]
- Tsukamoto, S.; Hirota, H.; Kato, H.; Fusetani, N. Isolation of eicosapentaenoyl and arachidonoyl lysophosphatidylinositols from the ascidian Halocynthia roretzi. Comp. Biochem. Physiol. Part C 1993, 106, 151–153. [Google Scholar]
- Nakao, Y.; Takada, K.; Matsunaga, S.; Fusetani, N. Calyceramides A–C: Neuraminidase inhibitory sulfated ceramides from the marine sponge Discodermia calyx. Tetrahedron 2001, 57, 3013–3017. [Google Scholar]
- Ojika, M.; Yoshino, G.; Sakagami, Y. Novel ceramide 1-sulfates, potent DNA topoisomerase I inhibitors isolated from the bryozoa Watersipora cucullata. Tetrahedron Lett. 1997, 38, 4235–4238. [Google Scholar]
- Barbieri, L.; Costantino, V.; Fattorusso, E.; Mangoni, A. Glycolipids from sponges. Part 16. Discoside, a rare myo-inositol-containing glycolipid from the Caribbean sponge Discodermia dissoluta. J. Nat. Prod. 2005, 68, 1527–1530. [Google Scholar]
- Prottey, C.; Ballou, C.E. Diacyl myoinositol monomannoside from Propionibacterium shermanii. J. Biol. Chem. 1968, 243, 6196–6201. [Google Scholar]
- Gross, H.; Reitner, J.; Koenig, G.M. Isolation and structure elucidation of azoricasterol, a new sterol of the deepwater sponge Macandrewia azorica. Naturwissenschaften 2004, 91, 441–446. [Google Scholar]
- Keffer, J.L.; Plaza, A.; Bewley, C.A. Motualevic acids A–F, antimicrobial acids from the sponge Siliquariaspongia sp. Org. Lett. 2009, 11, 1087–1090. [Google Scholar]
- Ratnayake, A.S.; Davis, R.A.; Harper, M.K.; Veltri, C.A.; Andjelic, C.D.; Barrows, L.R.; Ireland, C.M. Aurantosides G, H, and I: Three new tetramic acid glycosides from a Papua New Guinea Theonella swinhoei. J. Nat. Prod. 2004, 68, 104–107. [Google Scholar]
- Matsunaga, S.; Fusetani, N.; Kato, Y.; Hirota, H. Aurantosides A and B: Cytotoxic tetramic acid glycosides from the marine sponge Theonella sp. J. Am. Chem. Soc. 1991, 113, 9690–9692. [Google Scholar]
- Zhang, H.J.; Yi, Y.H.; Lin, H.W. Oxygenated 4-methylidene sterols from the South China Sea sponge Theonella swinhoei. Helv. Chim. Acta 2010, 93, 1120–1126. [Google Scholar]
- De Marino, S.; Ummarino, R.; D’Auria, M.V.; Chini, M.G.; Bifulco, G.; Renga, B.; D’Amore, C.; Fiorucci, S.; Debitus, C.; Zampella, A. Theonellasterols and conicasterols from Theonella swinhoei. Novel marine natural ligands for human nuclear receptors. J. Med. Chem. 2011, 54, 3065–3075. [Google Scholar]
- Qureshi, A.; Faulkner, D.J. 7α-Hydroxytheonellasterol, a cytotoxic 4-methylene sterol from the Philippines sponge Theonella swinhoei. J. Nat. Prod. 2000, 63, 841–842. [Google Scholar]
- Kho, E.; Imagawa, D.K.; Rohmer, M.; Kashman, Y.; Djerassi, C. Sterols in marine invertebrates. 22. Isolation and structure elucidation of conicasterol and theonellasterol, two new 4-methylene sterols from the Red Sea sponges Theonella conica and Theonella swinhoei. J. Org. Chem. 1981, 46, 1836–1839. [Google Scholar]
- Umeyama, A.; Shoji, N.; Enoki, M.; Arihara, S. Swinhosterols A–C, 4-methylene secosteroids from the marine sponge Theonella swinhoei. J. Nat. Prod. 1997, 60, 296–298. [Google Scholar]
- de Marino, S.; Sepe, V.; D’Auria, M.V.; Bifulco, G.; Renga, B.; Petek, S.; Fiorucci, S.; Zampella, A. Towards new ligands of nuclear receptors. Discovery of malaitasterol A, an unique bis-secosterol from marine sponge Theonella swinhoei. Org. Biomol. Chem. 2011, 9, 4856–4862. [Google Scholar]
- Cohen, J.; Paul, G.; Gunasekera, S.; Longley, R.; Pomponi, S. 6-Hydroxydiscodermindole, A new discodermindole from the marine sponge Discodermia polydiscus. Pharmaceut. Biol. 2004, 42, 59–61. [Google Scholar]
- Mickel, S.J. Total Synthesis of the marine natural product (+)-discodermolide in multigram quantities. Pure Appl. Chem. 2007, 79, 685–700. [Google Scholar]
- Smith, A.B.; Kaufman, M.D.; Beauchamp, T.J.; LaMarche, M.J.; Arimoto, H. Gram-scale synthesis of (+)-discodermolide. Org. Lett. 1999, 1, 1823–1826. [Google Scholar]
- Smith, A.B., III; Freeze, B.S. (+)-Discodermolide: Total synthesis, construction of novel analogues, and biological evaluation. Tetrahedron 2008, 64, 261–298. [Google Scholar]
- Buey, R.M.; Barasoain, I.; Jackson, E.; Meyer, A.; Giannakakou, P.; Paterson, I.; Mooberry, S.; Andreu, J.M.; Díaz, J.F. Microtubule interactions with chemically diverse stabilizing agents: Thermodynamics of binding to the paclitaxel site predicts cytotoxicity. Chem. Biol. 2005, 12, 1269–1279. [Google Scholar]
- Martello, L.; McDaid, H.; Regl, D.; Yang, C.; Meng, D.; Pettus, T.; Kaufman, M.; Arimoto, H.; Danishefsky, S.; Smith, A., III; et al. Taxol and discodermolide represent a synergistic drug combination in human carcinoma cell lines. Clin. Cancer Res. 2000, 6, 1978–1987. [Google Scholar]
- Shaw, S.J. The structure activity relationship of discodermolide analogues. Mini Rev. Med. Chem. 2008, 8, 276–284. [Google Scholar]
- O’Neil, G.W.; Phillips, A.J. Total synthesis of (−)-dictyostatin. J. Am. Chem. Soc. 2006, 128, 5340–5341. [Google Scholar]
- Ramachandran, P.V.; Srivastava, A.; Hazra, D. Total synthesis of potential antitumor agent, (−)-dictyostatin. Org. Lett. 2006, 9, 157–160. [Google Scholar]
- Paterson, I.; Gardner, N.M. Design, synthesis and biological evaluation of a macrocyclic discodermolide/dictyostatin hybrid. Chem. Commun. (Camb.) 2007, 49–51. [Google Scholar]
- Paterson, I.; Gardner, N.M.; Guzman, E.; Wright, A.E. Total synthesis and biological evaluation of potent analogues of dictyostatin: Modification of the C2-C6 dienoate region. Bioorg. Med. Chem. Lett. 2008, 18, 6268–6272. [Google Scholar]
- Paterson, I.; Gardner, N.M.; Poullennec, K.G.; Wright, A.E. Synthesis and biological evaluation of novel analogues of dictyostatin. Bioorg. Med. Chem. Lett. 2007, 17, 2443–2447. [Google Scholar]
- Paterson, I.; Gardner, N.M.; Poullennec, K.G.; Wright, A.E. Synthesis and biological evaluation of 10,11-dihydrodictyostatin, a potent analogue of the marine anticancer agent dictyostatin. J. Nat. Prod. 2008, 71, 364–369. [Google Scholar]
- Shin, Y.; Fournier, J.H.; Bruckner, A.; Madiraju, C.; Balachandran, R.; Raccor, B.S.; Edler, M.C.; Hamel, E.; Sikorski, R.P.; Vogt, A.; et al. Synthesis and biological evaluation of (−)-dictyostatin and stereoisomers. Tetrahedron 2007, 63, 8537–8562. [Google Scholar]
- Shin, Y.; Fournier, J.-H.; Balachandran, R.; Madiraju, C.; Raccor, B.S.; Zhu, G.; Edler, M.C.; Hamel, E.; Day, B.W.; Curran, D.P. Synthesis and biological evaluation of (−)-16-normethyldictyostatin: A potent analogue of (−)-dictyostatin. Org. Lett. 2005, 7, 2873–2876. [Google Scholar]
- Zampella, A.; D’Auria, M.V.; Minale, L.; Debitus, C.; Roussakis, C. Callipeltoside A: A cytotoxic aminodeoxy sugar-containing macrolide of a new type from the marine Lithistida sponge Callipelta sp. J. Am. Chem. Soc. 1996, 118, 11085–11088. [Google Scholar]
- Trost, B.M.; Gunzner, J.L.; Dirat, O.; Rhee, Y.H. Callipeltoside A: Total synthesis, assignment of the absolute and relative configuration, and evaluation of synthetic analogues. J. Am. Chem. Soc. 2002, 124, 10396–10415. [Google Scholar]
- Evans, D.A.; Burch, J.D.; Hu, E.; Jaeschke, G. Enantioselective total synthesis of callipeltoside A: Two approaches to the macrolactone fragment. Tetrahedron 2008, 64, 4671–4699. [Google Scholar]
- Huang, H.; Panek, J.S. Total synthesis of callipeltoside A. Org. Lett. 2004, 6, 4383–4385. [Google Scholar]
- Paterson, I.; Davies, R.D.M.; Heimann, A.C.; Marquez, R.; Meyer, A. Stereocontrolled total synthesis of (−)-callipeltoside A. Org. Lett. 2003, 5, 4477–4480. [Google Scholar]
- Carpenter, J.; Northrup, A.B.; Chung, D.; Wiener, J.J.M.; Kim, S.; MacMillan, D.W.C. Total synthesis and structural revision of callipeltoside C. Angew. Chem. Int. Ed. 2008, 47, 3568–3572. [Google Scholar]
- Zampella, A.; D’Auria, M.V.; Minale, L.; Debitus, C. Callipeltosides B and C, two novel cytotoxic glycoside macrolides from a marine lithistida sponge Callipelta sp. Tetrahedron 1997, 53, 3243–3248. [Google Scholar]
- D’Auria, M.V.; Zampella, A.; Paloma, L.G.; Minale, L.; Debitus, C.; Roussakis, C.; le Bert, V. Callipeltins B and C; bioactive peptides from a marine Lithistida sponge Callipelta sp. Tetrahedron 1996, 52, 9589–9596. [Google Scholar]
- Bassarello, C.; Zampella, A.; Monti, M.C.; Gomez-Paloma, L.; D’Auria, M.V.; Riccio, R.; Bifulco, G. Quantum mechanical calculation of coupling constants in the configurational analysis of flexible systems: Determination of the configuration of callipeltin A. Eur J. Org. Chem. 2006, 604–609. [Google Scholar]
- Zampella, A.; D’Orsi, R.; Sepe, V.; Casapullo, A.; Monti, M.C.; D’Auria, M.V. Concise synthesis of all stereoisomers of β-methoxytyrosine and determination of the absolute configuration of the residue in callipeltin A. Org. Lett. 2005, 7, 3585–3588. [Google Scholar]
- Kikuchi, M.; Watanabe, Y.; Tanaka, M.; Akaji, K.; Konno, H. Synthesis and cytotoxicity of the depsipeptides analogues of callipeltin B. Bioorg. Med. Chem. Lett. 2011, 21, 4865–4868. [Google Scholar]
- Allingham, J.S.; Zampella, A.; D’Auria, M.V.; Rayment, I. Structures of microfilament destabilizing toxins bound to actin provide insight into toxin design and activity. Proc. Natl. Acad. Sci. USA 2005, 102, 14527–14532. [Google Scholar]
- D’Auria, M.V.; Paloma, L.G.; Minale, L.; Zampella, A.; Verbist, J.-F.; Roussakis, C.; Debitus, C.; Patissou, J. Reidispongiolide A and B, two new potent cytotoxic macrolides from the New Caledonian sponge Reidispongia coerulea. Tetrahedron 1994, 50, 4829–4834. [Google Scholar]
- Paterson, I.; Ashton, K.; Britton, R.; Cecere, G.; Chouraqui, G.; Florence, G.J.; Knust, H.; Stafford, J. Total synthesis of (−)-reidispongiolide A, an actin-targeting macrolide isolated from the marine sponge Reidispongia coerulea. Chem. Asian J. 2008, 3, 367–387. [Google Scholar]
- Fuwa, H.; Naito, S.; Goto, T.; Sasaki, M. Total synthesis of (+)-neopeltolide. Angew. Chem. Int. Ed. 2008, 47, 4737–4739. [Google Scholar]
- Kartika, R.; Gruffi, T.R.; Taylor, R.E. Concise enantioselective total synthesis of neopeltolide macrolactone highlighted by ether transfer. Org. Lett. 2008, 10, 5047–5050. [Google Scholar]
- Paterson, I.; Miller, N.A. Total synthesis of the marine macrolide (+)-neopeltolide. Chem. Commun. 2008, 4708–4710. [Google Scholar]
- Custar, D.W.; Zabawa, T.P.; Hines, J.; Crews, C.M.; Scheidt, K.A. Total synthesis and structure activity investigation of the marine natural product neopeltolide. J. Am. Chem. Soc. 2009, 131, 12406–12414. [Google Scholar]
- Xie, W.; Ding, D.; Zi, W.; Li, G.; Ma, D. Total synthesis and structure assignment of papuamide B, a potent marine cyclodepsipeptide with anti-HIV properties. Angew. Chem. Int. Ed. 2008, 47, 2844–2848. [Google Scholar]
- D’Auria, M.V.; Debitus, C.; Paloma, L.G.; Minale, L.; Zampella, A. Superstolide A: A potent cytotoxic macrolide of a new type from the New Caledonian deep water marine sponge Neosiphonia superstes. J. Am. Chem. Soc. 2002, 116, 6658–6663. [Google Scholar]
- Tortosa, M.; Yakelis, N.A.; Roush, W.R. Total synthesis of (+)-superstolide A. J. Org. Chem. 2008, 73, 9657–9667. [Google Scholar]
- Zhu, J.; Ma, D. Total synthesis of microsclerodermin E. Angew. Chem. Int. Ed. 2003, 42, 5348–5351. [Google Scholar]
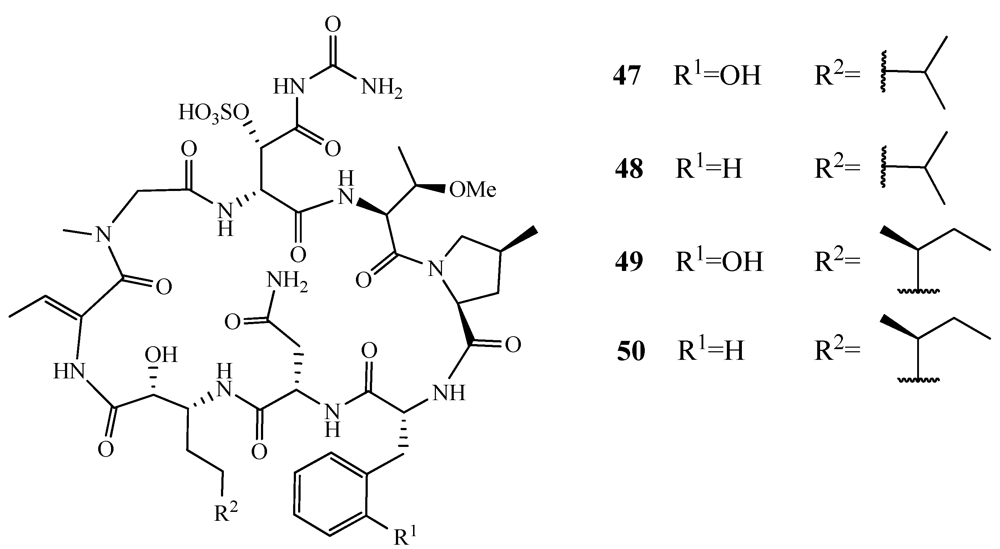
- Nakao, Y.; Oku, N.; Matsunaga, S.; Fusetani, N. Cyclotheonamides E2 and E3, new potent serine protease inhibitors from the marine sponge of the genus Theonella. J. Nat. Prod. 1998, 61, 667–670. [Google Scholar]
- Fusetani, N.; Matsunaga, S.; Matsumoto, H.; Takebayashi, Y. Bioactive marine metabolites. 33. Cyclotheonamides, potent thrombin inhibitors, from a marine sponge Theonella sp. J. Am. Chem. Soc. 1990, 112, 7053–7054. [Google Scholar]
- Hagihara, M.; Schreiber, S.L. Reassignment of stereochemistry and total synthesis of the thrombin inhibitor cyclotheonamide B. J. Am. Chem. Soc. 1992, 114, 6570–6571. [Google Scholar]
- Wipf, P.; Kim, H. Total synthesis of cyclotheonamide A. J. Org. Chem. 1993, 58, 5592–5594. [Google Scholar]
- Wasserman, H.H.; Zhang, R. Application of cyano ylide methodology to the synthesis of cyclotheonamides E2 and E3. Tetrahedron 2002, 58, 6277–6283. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Winder, P.L.; Pomponi, S.A.; Wright, A.E. Natural Products from the Lithistida: A Review of the Literature since 2000. Mar. Drugs 2011, 9, 2643-2682. https://doi.org/10.3390/md9122643
Winder PL, Pomponi SA, Wright AE. Natural Products from the Lithistida: A Review of the Literature since 2000. Marine Drugs. 2011; 9(12):2643-2682. https://doi.org/10.3390/md9122643
Chicago/Turabian StyleWinder, Priscilla L., Shirley A. Pomponi, and Amy E. Wright. 2011. "Natural Products from the Lithistida: A Review of the Literature since 2000" Marine Drugs 9, no. 12: 2643-2682. https://doi.org/10.3390/md9122643
APA StyleWinder, P. L., Pomponi, S. A., & Wright, A. E. (2011). Natural Products from the Lithistida: A Review of the Literature since 2000. Marine Drugs, 9(12), 2643-2682. https://doi.org/10.3390/md9122643