An Investigation into the Cytotoxic Effects of 13-Acetoxysarcocrassolide from the Soft Coral Sarcophyton crassocaule on Bladder Cancer Cells

Abstract
:1. Introduction

2 Materials and Methods
2.1. Materials
2.2. Cell Culture and the Treatment with 13-Acetoxysarcocrassolide
2.3. MTT Assay
2.4. Wound-Healing Assay
2.5. Analysis of Cell Cycle and Apoptosis
2.6. Protein Extraction and Preparation
2.7. Two-Dimensional Gel Electrophoresis
2.8. Protein Identification by LC-MS/MS
2.9. Western Blot Analysis
3 Results
3.1. The Anti-Proliferative Effect of 13-Acetoxysarcocrassolide on BFTC Cells


3.2. The Anti-Migratory Effect of 13-Acetoxysarcocrassolide on BFTC Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 h | 6 h | 12 h | 24 h | |
|---|---|---|---|---|
| control | 8.333 ± 3.215 | 46 ± 8 | 158.67 ± 18.175 | 344 ± 14.177 |
| 1 μg 13-acetoxysarcocrassolide | 10.67 ± 3.512 | 17.67 ± 6.506 | 107.31 ± 5.275 * | 219.67 ± 21.197 * |
| 1.5 μg 13-acetoxysarcocrassolide | 5.33 ± 2.309 | 14 ± 5 | 79 ± 10.44 * | 155.33 ± 12.22 * |
| 3 μg 13-acetoxysarcocrassolide | 4.67 ± 3.055 | 11.33 ± 5.508 | 53.33 ± 15.503 * | 105.33 ± 12.662 * |
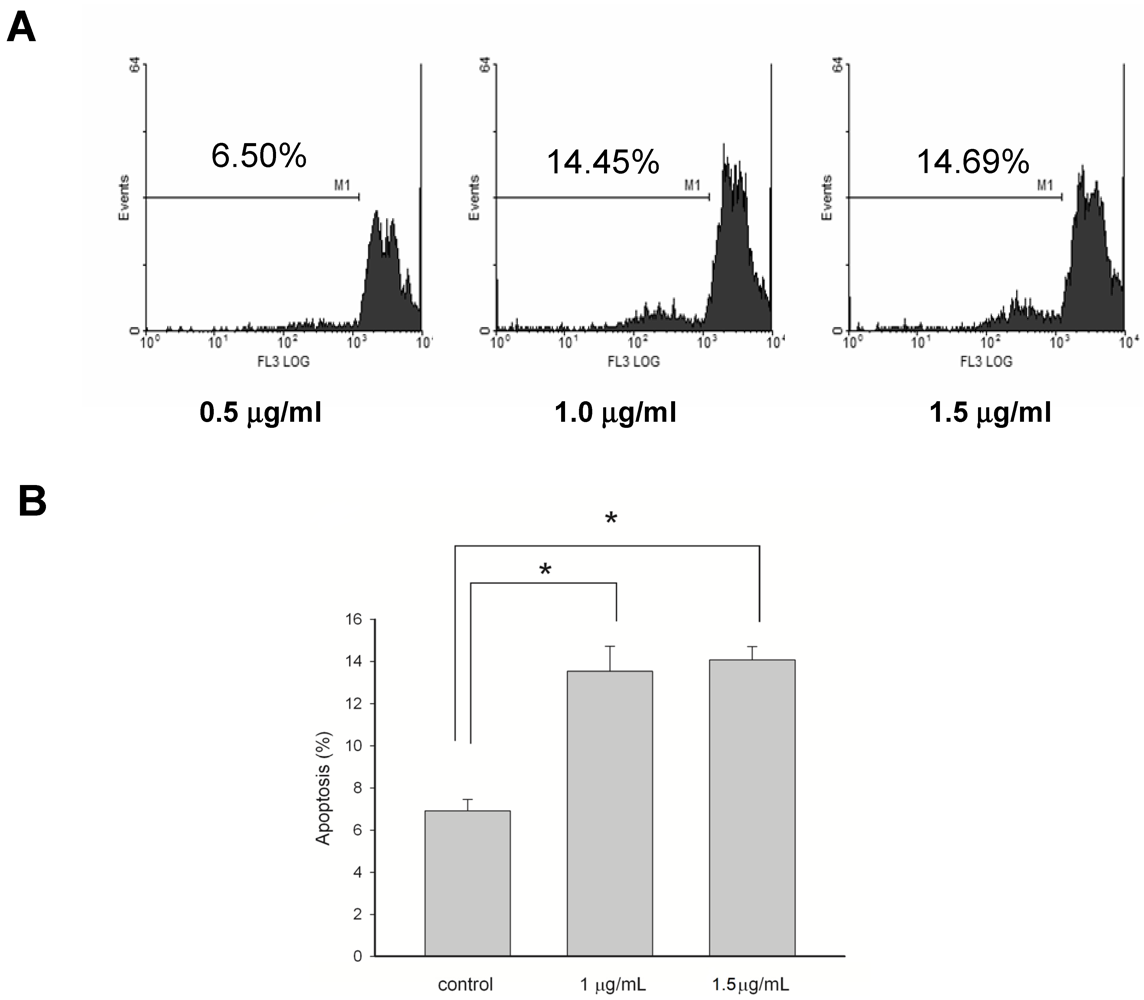
3.3. Apoptosis-Induced Effects of 13-Acetoxysarcocrassolide on BFTC Cells

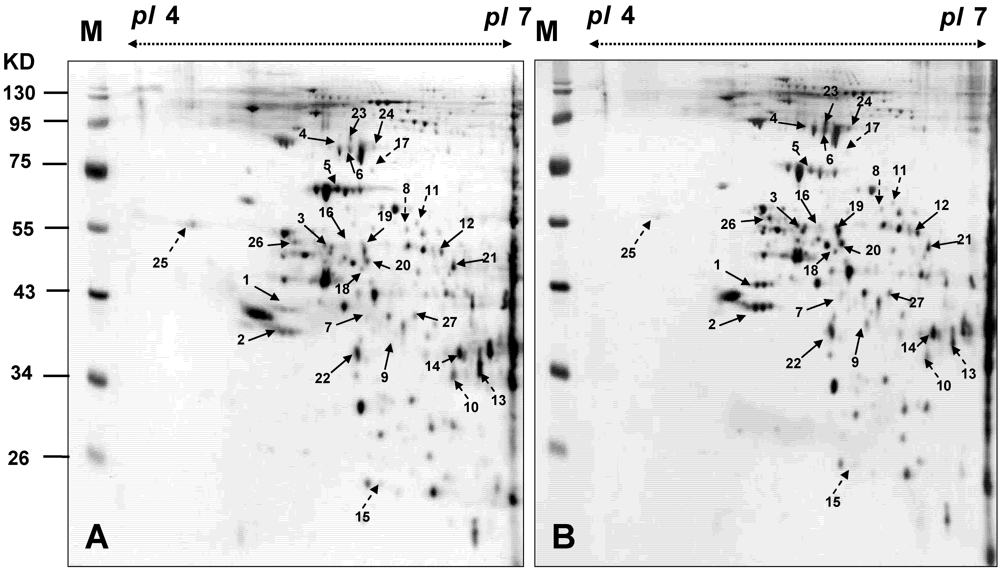
3.4. Proteomic Analysis of BFTC Cells Treated with 13-Acetoxysarcocrassolide

| Spot no. | Protein name | Accession no. | Calculate Mr/pI | Peptide matched | Sequence covered % | MASCOT score | Regulation (fold-change) * |
|---|---|---|---|---|---|---|---|
| 1 | Heterogeneous nuclear ribonucleoproteins C1/C2 | P07910 | 33.65/4.95 | 10 | 21 | 99 | +7.0 |
| 2 | Heterogeneous nuclear ribonucleoproteins C1/C2 | P07910 | 33.65/4.95 | 14 | 35 | 163 | +8.3 |
| 3 | Heterogeneous nuclear ribonucleoprotein F | P52597 | 45.64/5.38 | 20 | 31 | 201 | +2.0 |
| 4 | Heat shock cognate 71 kDa protein | P11142 | 70.85/5.37 | 7 | 14 | 93 | +3.7 |
| 5 | 60 kDa heat shock protein | P10809 | 61.01/5.7 | 35 | 39 | 686 | −2.0 |
| 6 | Stress-70 protein | P38646 | 73.63/5.87 | 27 | 37 | 333 | +2.2 |
| 7 | Arginase-2 | P78540 | 38.55/6.0 | 2 | 8 | 70 | +2.0 |
| 8 | Protein disulfide-isomerase A3 precursor | P30101 | 54.74/5.98 | 26 | 41 | 387 | −2.3 |
| 9 | l-lactate dehydrogenase B chain | P07195 | 36.61/5.71 | 7 | 16 | 63 | +2.6 |
| 10 | Isocitrate dehydrogenase [NAD] subunit alpha | P50213 | 39.56/6.47 | 7 | 15 | 127 | −3.2 |
| 11 | Protein disulfide-isomerase A3 precursor | P30101 | 54.74/5.98 | 14 | 29 | 211 | −1.8 |
| 12 | Mitochondrial-processing peptidase subunit beta | O75439 | 54.33/6.38 | 11 | 23 | 126 | +2.8 |
| 13 | Heterogeneous nuclear ribonucleoprotein H3 (hnRNP H3) | P31942 | 36.9/6.37 | 3 | 10 | 74 | −2.1 |
| 14 | Palmitoyl-protein thioesterase 1 precursor | P50897 | 34.17/6.07 | 5 | 10 | 83 | +1.8 |
| 15 | Proteasome subunit beta type 4 precursor | P28070 | 29.18/5.72 | 6 | 24 | 92 | −3.0 |
| 16 | Thioredoxin domain-containing protein 5 precursor | Q8NBS9 | 47.59/5.63 | 4 | 11 | 69 | +2.2 |
| 17 | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex | P10515 | 65.73/5.79 | 5 | 2 | 63 | −2.0 |
| 18 | Protein NDRG1 | Q92597 | 42.8/5.49 | 9 | 25 | 129 | +2.3 |
| 19 | Ubiquinol-cytochrome-c reductase complex core protein 1 | P31930 | 52.61/5.94 | 9 | 15 | 156 | +2.3 |
| 20 | Ubiquinol-cytochrome-c reductase complex core protein 1 | P31930 | 52.61/5.94 | 2 | 3 | 67 | +2.2 |
| 21 | Ornithine aminotransferase | P04181 | 48.5/6.57 | 23 | 30 | 296 | +1.5 |
| 22 | Guanine nucleotide-binding protein G(I)/G(S)/G(T) | P62873 | 37.35/5.6 | 12 | 20 | 151 | +1.9 |
| 23 | Stress-70 protein | P38646 | 73.63/5.87 | 17 | 21 | 190 | +2.7 |
| 24 | Heat shock cognate 71 kDa protein | P11142 | 70.85/5.37 | 14 | 17 | 137 | +2.2 |
| 25 | Calreticulin precursor | P27797 | 48.11/4.29 | 13 | 17 | 99 | −3.2 |
| 26 | ATP synthase subunit beta | P06576 | 56.52/5.26 | 20 | 34 | 256 | +2.8 |
| 27 | Glial fibrillary acidic protein | P14136 | 49.85/5.42 | 3 | 4 | 53 | +2.4 |
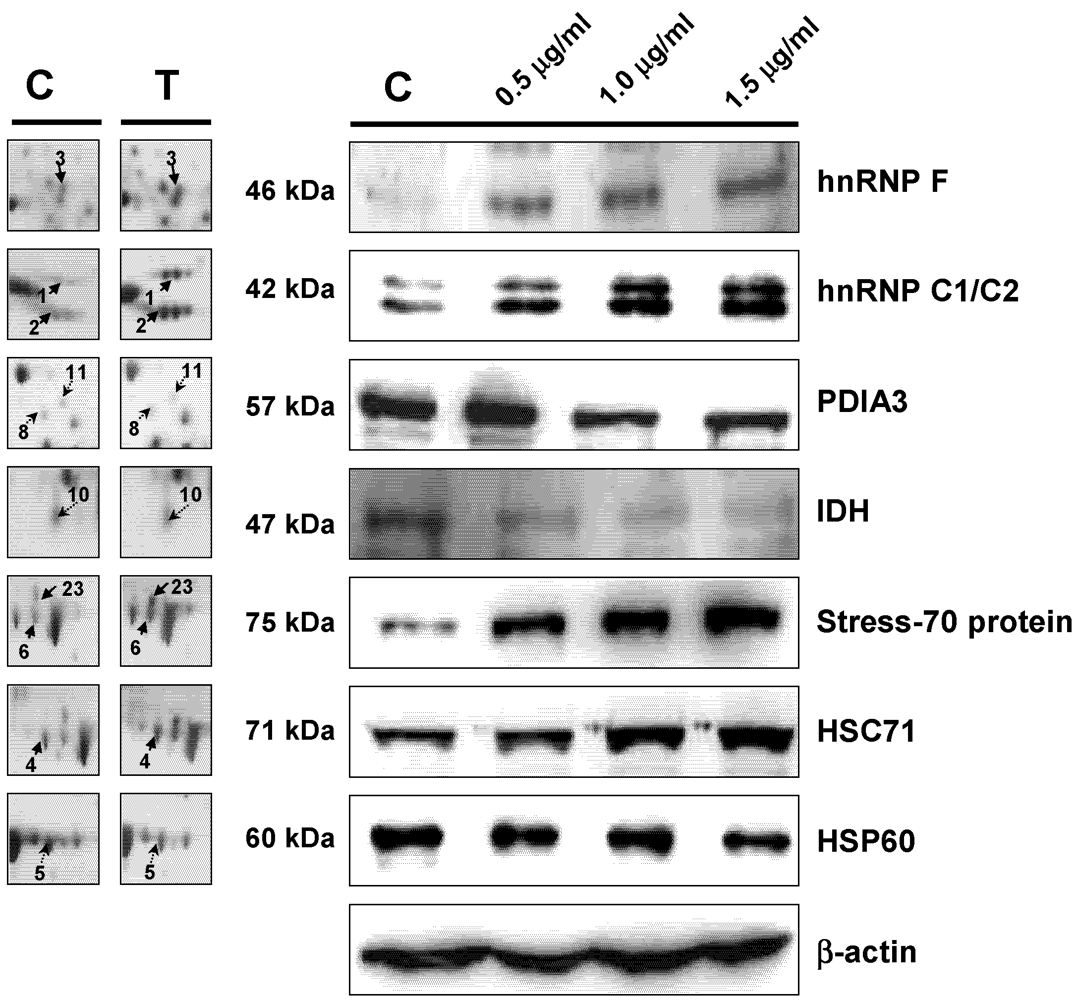
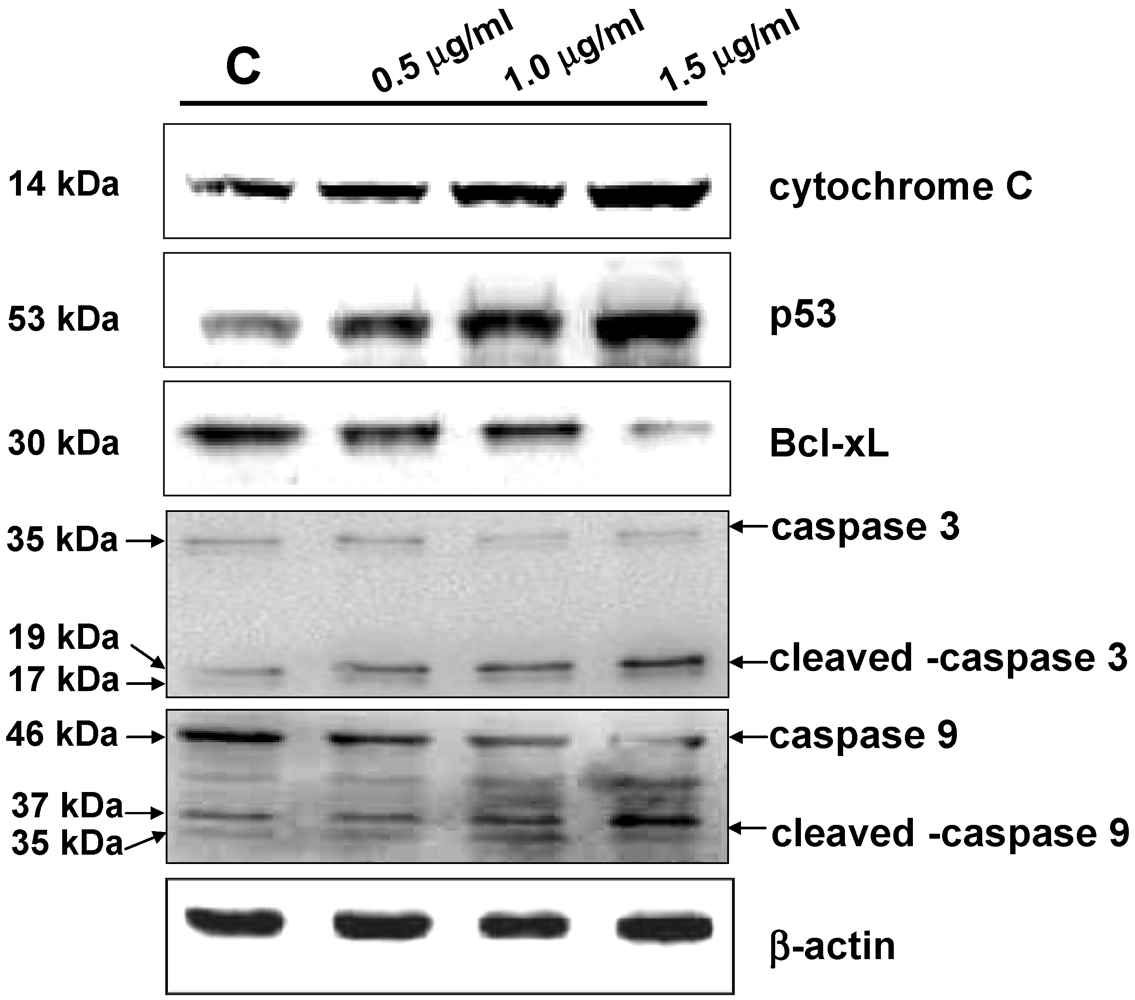
3.5. Western Blot Analysis


4. Discussion
5. Conclusion
Acknowledgments
Conflict of Interest
- Samples Availability: Available from the authors.
References
- Azemar, M.D.; Audouin, M.; Revaux, A.; Misrai, V.; Comperat, E.; Bitker, M.O.; Chartier-Kastler, E.; Richard, F.; Cussenot, O.; Roupret, M. Primary upper urinary tract tumors and subsequent location in the bladder. Prog. Urol. 2009, 19, 583–588. [Google Scholar]
- Dzombeta, T.; Krajacic-Jagarcec, G.; Tomas, D.; Kraus, O.; Ruzic, B.; Kruslin, B. Urothelial carcinoma with an inverted growth pattern: a report of 4 cases. Acta Med. Croatica 2010, 64, 47–50. [Google Scholar]
- Ruiz, E.; Alarcon Caba, M.; Toselli, L.; Moldes, J.; Ormaechea, M.; de Badiola, F.; Christiansen, S. Transitional cell carcinoma of the bladder in adolescents: A diagnosis to bear in mind. Arch. Argent. Pediatr. 2009, 107, 49–52. [Google Scholar]
- Hsieh, J.L.; Wu, C.L.; Lai, M.D.; Lee, C.H.; Tsai, C.S.; Shiau, A.L. Gene therapy for bladder cancer using E1B-55 kD-deleted adenovirus in combination with adenoviral vector encoding plasminogen kringles 1–5. Br. J. Cancer 2003, 88, 1492–1499. [Google Scholar]
- Goebell, P.J.; Legal, W.; Weiss, C.; Fietkau, R.; Wullich, B.; Krause, S. Multimodal therapy for bladder sparing with high grade bladder tumors. Urol. A 2008, 47, 838, 840–842, 844–845. [Google Scholar]
- Shi, B.; Zhang, K.; Zhang, J.; Chen, J.; Zhang, N.; Xu, Z. Relationship between patient age and superficial transitional cell carcinoma characteristics. Urology 2008, 71, 1186–1190. [Google Scholar]
- Amling, C.L. Diagnosis and management of superficial bladder cancer. Curr. Probl. Cancer 2001, 25, 219–278. [Google Scholar]
- Babjuk, M.; Dvoracek, J. Diagnosis and therapy of superficial tumors of the urinary bladder. Cas. Lek. Cesk. 2002, 141, 723–728. [Google Scholar]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 2001, 18, 1–49. [Google Scholar]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 2002, 19, 1–48. [Google Scholar]
- Ojika, M.; Islam, M.K.; Shintani, T.; Zhang, Y.; Okamoto, T.; Sakagami, Y. Three new cytotoxic acylspermidines from the soft coral, Sinularia sp. Biosci. Biotechnol. Biochem. 2003, 67, 1410–1412. [Google Scholar]
- Poza, J.J.; Fernandez, R.; Reyes, F.; Rodriguez, J.; Jimenez, C. Isolation, biological significance, synthesis, and cytotoxic evaluation of new natural parathiosteroids A–C and analogues from the soft coral Paragorgia sp. J. Org. Chem. 2008, 73, 7978–7984. [Google Scholar]
- Kamel, H.N.; Ferreira, D.; Garcia-Fernandez, L.F.; Slattery, M. Cytotoxic diterpenoids from the hybrid soft coral Sinularia maxima × Sinularia polydactyla. J. Nat. Prod. 2007, 70, 1223–1227. [Google Scholar]
- Hassan, H.M.; Khanfar, M.A.; Elnagar, A.Y.; Mohammed, R.; Shaala, L.A.; Youssef, D.T.; Hifnawy, M.S.; El Sayed, K.A. Pachycladins A–E, prostate cancer invasion and migration inhibitory Eunicellin-based diterpenoids from the red sea soft coral Cladiella pachyclados. J. Nat. Prod. 2010, 73, 848–853. [Google Scholar]
- Chiang, P.C.; Chien, C.L.; Pan, S.L.; Chen, W.P.; Teng, C.M.; Shen, Y.C.; Guh, J.H. Induction of endoplasmic reticulum stress and apoptosis by a marine prostanoid in human hepatocellular carcinoma. J. Hepatol. 2005, 43, 679–686. [Google Scholar]
- Chiang, P.C.; Kung, F.L.; Huang, D.M.; Li, T.K.; Fan, J.R.; Pan, S.L.; Shen, Y.C.; Guh, J.H. Induction of Fas clustering and apoptosis by coral prostanoid in human hormone-resistant prostate cancer cells. Eur. J. Pharmacol. 2006, 542, 22–30. [Google Scholar]
- Liu, C.-I.; Chen, C.-C.; Chen, J.-C.; Su, J.-H.; Huang, H.H.; Chen, J.Y.-F.; Wu, Y.-J. Proteomic Analysis of Anti-Tumor Effects of 11-Dehydrosinulariolide on CAL-27 Cells. Mar. Drugs 2011, 9, 1254–1272. [Google Scholar]
- Ahmed, A.; Shiue, R.; Wang, G.; Dai, C.; Kuo, Y.; Sheu, J. Five novel norcembranoids from Sinularia leptoclados and S. parva. Tetrahedron 2003, 59, 7337–7344. [Google Scholar]
- Chang, H.T.; Huang, J.K.; Wang, J.L.; Cheng, J.S.; Lee, K.C.; Lo, Y.K.; Lin, M.C.; Tang, K.Y.; Jan, C.R. Tamoxifen-induced Ca2+ mobilization in bladder female transitional carcinoma cells. Arch. Toxicol. 2001, 75, 184–188. [Google Scholar]
- Cheng, Y.T.; Li, Y.L.; Wu, J.D.; Long, S.B.; Tzai, T.S.; Tzeng, C.C.; Lai, M.D. Overexpression of MDM-2 mRNA and mutation of the p53 tumor suppressor gene in bladder carcinoma cell lines. Mol. Carcinog. 1995, 13, 173–181. [Google Scholar]
- Bowden, B.F.; Coll, J.C.; Mitchell, S.J. Studies of Australian soft corals. XVIII* Further cembranoid diterpenes from soft corals of the genus Sarcophyton. Aust. J. Chem. 1980, 33, 879–884. [Google Scholar]
- Duh, C.-Y.; Wang, S.-K.; Chung, S.-G.; Chou, G.-C.; Dai, C.-F. Cytotoxic cembrenolides and steroids from the Formosan soft coral Sarcophyton crassocaule. J. Nat. Prod. 2000, 63, 1634–1637. [Google Scholar]
- Xu, X.-H.; Kong, C.-H.; Lin, C.-J.; Wang, X.; Zhu, Y.-D.; Yang, H.-S. A novel diterpenoid from the soft coral Sarcophyton crassocaule. Chin. J. Chem. 2003, 21, 1506–1509. [Google Scholar]
- Zhang, C.; Li, J.; Su, J.; Liang, Y.-J.; Yang, X.-P.; Zheng, K.-C.; Zeng, L.-M. Cytotoxic diterpenoids from the soft coral Sarcophyton crassocaule. J. Nat. Prod. 2006, 69, 1476–1480. [Google Scholar]
- Huang, H.-C.; Ahmed, A.F.; Su, J.-H.; Wu, Y.-C.; Chiang, M.Y.; Sheu, J.-H. Crassocolides A–F, new cembranoids with a trans-fused lactone from the soft coral Sarcophyton crassocaule. J. Nat. Prod. 2006, 69, 1554–1559. [Google Scholar]
- Huang, H.-C.; Chao, C.-H.; Kuo, Y.-H.; Sheu, J.-H. Crassocolides G–M, cembranoids from a Formosan soft coral Sarcophyton crassocaule. Chem. Biodivers. 2009, 6, 1232–1242. [Google Scholar]
- Lin, W.-Y.; Su, J.-H.; Lu, Y.; Wen, Z.-H.; Dai, C.-F.; Kuo, Y.-H.; Sheu, J.-H. Cytotoxic and anti-inflammatory cembranoids from the Dongsha Atoll soft coral Sarcophyton crassocaule. Bioorg. Med. Chem. 2010, 18, 1936–1941. [Google Scholar]
- Lin, W.-Y.; Lu, Y.; Su, J.-H.; Wen, Z.-H.; Dai, C.-F.; Kuo, Y.-H.; Sheu, J.-H. Bioactive cembranoids from the Dongsha Atoll soft coral Sarcophyton crassocaule. Mar. Drugs 2011, 9, 994–1006. [Google Scholar]
- Zhang, C.-X.; Yan, S.-J.; Zhang, G.-W.; Su, J.-Y.; Zeng, L.-M. Isolation of new polyhydroxylated sterol from soft coral Sarcophyton crassocaule Mosre. Chem. J. Chin. Univ. 2007, 28, 686–688. [Google Scholar]
- Anjaneyulu, A.S.R.; Murthy, M.V.R.K.; Gowri, P.M. Novel epoxy steroids from the Indian ocean soft coral Sarcophyton crassocaule. J. Nat. Prod. 2000, 63, 112–118. [Google Scholar]
- Anjaneyulu, A.S.R.; Murthy, M.V.R.K.; Rao, N.S.K. New hippurins from the Indian Ocean soft coral Sarcophyton crassocaule. J. Chem. Res. (S) 1997, 450–451. [Google Scholar]
- Anjaneyulu, A.S.R.; Murthy, M.V.R.K.; Gowri, P.M.; Venugopal, M.J.R.V.; Laatsch, H. A rare prostaglandin from the soft coral Sarcophyton crassocaule of the Indian Ocean. J. Nat. Prod. 2000, 63, 1425–1426. [Google Scholar]
- Li, L.; Wang, C.-Y.; Shao, C.-L.; Han, L.; Sun, X.-P.; Zhao, J.; Guo, Y.-W.; Huang, H.; Guan, H.-S. Two new metabolites from the Hainan soft coral Sarcophyton crassocaule. J. Asian Nat. Prod. Res. 2009, 11, 851–855. [Google Scholar]
- Hsieh, P.W.; Chang, F.R.; McPhail, A.T.; Lee, K.H.; Wu, Y.C. New cembranolide analogues from the formosan soft coral Sinularia flexibilis and their cytotoxicity. Nat. Prod. Res. 2003, 17, 409–418. [Google Scholar]
- Huang, H.H.; Brennan, T.C.; Muir, M.M.; Mason, R.S. Functional alpha1- and beta2-adrenergic receptors in human osteoblasts. J. Cell. Physiol. 2009, 220, 267–275. [Google Scholar]
- Wu, Y.J.; Chen, H.M.; Wu, T.T.; Wu, J.S.; Chu, R.M.; Juang, R.H. Preparation of monoclonal antibody bank against whole water-soluble proteins from rapid-growing bamboo shoots. Proteomics 2006, 6, 5898–5902. [Google Scholar]
- Lu, C.M.; Wu, Y.J.; Chen, C.C.; Hsu, J.L.; Chen, J.C.; Chen, J.Y.; Huang, C.H.; Ko, Y.C. Identification of low-abundance proteins via fractionation of the urine proteome with weak anion exchange chromatography. Proteome Sci. 2011, 9, 17. [Google Scholar]
- Xanthoudakis, S.; Roy, S.; Rasper, D.; Hennessey, T.; Aubin, Y.; Cassady, R.; Tawa, P.; Ruel, R.; Rosen, A.; Nicholson, D.W. Hsp60 accelerates the maturation of pro-caspase-3 by upstream activator proteases during apoptosis. EMBO J. 1999, 18, 2049–2056. [Google Scholar]
- Ghosh, J.C.; Dohi, T.; Kang, B.H.; Altieri, D.C. Hsp60 regulation of tumor cell apoptosis. J. Biol. Chem. 2008, 283, 5188–5194. [Google Scholar]
- Garneau, D.; Revil, T.; Fisette, J.F.; Chabot, B. Heterogeneous nuclear ribonucleoprotein F/H proteins modulate the alternative splicing of the apoptotic mediator Bcl-x. J. Biol. Chem. 2005, 280, 22641–22650. [Google Scholar]
- Rehm, M.; Dussmann, H.; Janicke, R.U.; Tavare, J.M.; Kogel, D.; Prehn, J.H. Single-cell fluorescence resonance energy transfer analysis demonstrates that caspase activation during apoptosis is a rapid process. Role of caspase-3. J. Biol. Chem. 2002, 277, 24506–24514. [Google Scholar]
- Lopergolo, A.; Pennati, M.; Gandellini, P.; Orlotti, N.I.; Poma, P.; Daidone, M.G.; Folini, M.; Zaffaroni, N. Apollon gene silencing induces apoptosis in breast cancer cells through p53 stabilisation and caspase-3 activation. Br. J. Cancer 2009, 100, 739–746. [Google Scholar]
- Day, T.W.; Wu, C.H.; Safa, A.R. Etoposide induces protein kinase Cdelta- and caspase-3-dependent apoptosis in neuroblastoma cancer cells. Mol. Pharmacol. 2009, 76, 632–640. [Google Scholar]
- Pengju, Z.; Weiwen, C.; Aiying, W.; Zhaobo, C.; Nana, N.; Zhaoqin, H.; Qingwei, L.; Anli, J. NKX3.1 potentiates TNF-alpha/CHX-induced apoptosis of prostate cancer cells through increasing caspase-3 expression and its activity. Biochem. Biophys. Res. Commun. 2010, 398, 457–461. [Google Scholar]
- Liu, C.; Wu, X.; Luo, C.; Hu, Z.; Yin, Z.; He, Y.; Du, H.; Zhang, W.; Jiang, Q.; Lin, Y. Antisense oligonucleotide targeting Livin induces apoptosis of human bladder cancer cell via a mechanism involving caspase 3. J. Exp. Clin. Cancer Res. 2010, 29, 63. [Google Scholar]
- Jiang, X.H.; Wong, B.C.; Yuen, S.T.; Jiang, S.H.; Cho, C.H.; Lai, K.C.; Lin, M.C.; Kung, H.F.; Lam, S.K. Arsenic trioxide induces apoptosis in human gastric cancer cells through up-regulation of p53 and activation of caspase-3. Int. J. Cancer 2001, 91, 173–179. [Google Scholar]
- Vitale, I.; Antoccia, A.; Cenciarelli, C.; Crateri, P.; Meschini, S.; Arancia, G.; Pisano, C.; Tanzarella, C. Combretastatin CA-4 and combretastatin derivative induce mitotic catastrophe dependent on spindle checkpoint and caspase-3 activation in non-small cell lung cancer cells. Apoptosis 2007, 12, 155–166. [Google Scholar]
- Chen, D.; Cao, J.; Tian, L.; Liu, F.; Sheng, X. Induction of apoptosis by casticin in cervical cancer cells through reactive oxygen species-mediated mitochondrial signaling pathways. Oncol. Rep. 2011, 26, 1287–1294. [Google Scholar]
- Wang, Y.; Deng, L.; Zhong, H.; Jiang, X.; Chen, J. Natural plant extract tubeimoside I promotes apoptosis-mediated Cell death in cultured human hepatoma (HepG2) cells. Biol. Pharm. Bull. 2011, 34, 831–838. [Google Scholar]
- Hamsa, T.P.; Thejass, P.; Kuttan, G. Induction of apoptosis by sulforaphane in highly metastatic B16F-10 melanoma cells. Drug Chem. Toxicol. 2011, 34, 332–340. [Google Scholar]
- Sato, M.; Tsujino, I.; Fukunaga, M.; Mizumura, K.; Gon, Y.; Takahashi, N.; Hashimoto, S. Cyclosporine a induces apoptosis of human lung adenocarcinoma cells via caspase-dependent pathway. Anticancer Res. 2011, 31, 2129–2134. [Google Scholar]
- Hsu, H.F.; Houng, J.Y.; Kuo, C.F.; Tsao, N.; Wu, Y.C. Glossogin, a novel phenylpropanoid from Glossogyne tenuifolia, induced apoptosis in A549 lung cancer cells. Food Chem. Toxicol. 2008, 46, 3785–3791. [Google Scholar]
- Creagh, E.M.; Sheehan, D.; Cotter, T.G. Heat shock proteins—modulators of apoptosis in tumour cells. Leukemia 2000, 14, 1161–1173. [Google Scholar]
- Guimaraes, A.J.; Frases, S.; Gomez, F.J.; Zancope-Oliveira, R.M.; Nosanchuk, J.D. Monoclonal antibodies to heat shock protein 60 alter the pathogenesis of Histoplasma capsulatum. Infect. Immun. 2009, 77, 1357–1367. [Google Scholar]
- Hwang, Y.J.; Lee, S.P.; Kim, S.Y.; Choi, Y.H.; Kim, M.J.; Lee, C.H.; Lee, J.Y.; Kim, D.Y. Expression of heat shock protein 60 kDa is upregulated in cervical cancer. Yonsei Med. J. 2009, 50, 399–406. [Google Scholar]
- Lee, K.A.; Shim, J.H.; Kho, C.W.; Park, S.G.; Park, B.C.; Kim, J.W.; Lim, J.S.; Choe, Y.K.; Paik, S.G.; Yoon, D.Y. Protein profiling and identification of modulators regulated by the E7 oncogene in the C33A cell line by proteomics and genomics. Proteomics 2004, 4, 839–848. [Google Scholar]
- Kang, Y.H.; Lee, K.A.; Kim, J.H.; Park, S.G.; Yoon, D.Y. Mitomycin C modulates DNA-double strand break repair genes in cervical carcinoma cells. Amino Acids 2010, 39, 1291–1298. [Google Scholar]
- Ruan, W.; Wang, Y.; Ma, Y.; Xing, X.; Lin, J.; Cui, J.; Lai, M. HSP60, a protein downregulated by IGFBP7 in colorectal carcinoma. J. Exp. Clin. Cancer Res. 2010, 29, 41. [Google Scholar]
- Ghosh, J.; Dohi, T.; Kang, B.; Altieri, D. Hsp60 regulation of tumor cell apoptosis. J. Biol. Chem. 2008, 283, 5188–5194. [Google Scholar]
- Castagna, A.; Antonioli, P.; Astner, H.; Hamdan, M.; Righetti, S.C.; Perego, P.; Zunino, F.; Righetti, P.G. A proteomic approach to cisplatin resistance in the cervix squamous cell carcinoma cell line A431. Proteomics 2004, 4, 3246–3267. [Google Scholar]
- Cecconi, D.; Astner, H.; Donadelli, M.; Palmieri, M.; Missiaglia, E.; Hamdan, M.; Scarpa, A.; Righetti, P.G. Proteomic analysis of pancreatic ductal carcinoma cells treated with 5-aza-2′-deoxycytidine. Electrophoresis 2003, 24, 4291–4303. [Google Scholar]
- Eleuterio, E.; Di Giuseppe, F.; Sulpizio, M.; di Giacomo, V.; Rapino, M.; Cataldi, A.; Di Ilio, C.; Angelucci, S. Proteome analysis of X-ray irradiated human erythroleukemia cells. Biochim. Biophys. Acta 2008, 1784, 611–620. [Google Scholar]
- Ronquist, K.G.; Carlsson, L.; Ronquist, G.; Nilsson, S.; Larsson, A. Prostasome-derived proteins capable of eliciting an immune response in prostate cancer patients. Int. J. Cancer 2006, 119, 847–853. [Google Scholar]
- Short, D.M.; Heron, I.D.; Birse-Archbold, J.L.; Kerr, L.E.; Sharkey, J.; McCulloch, J. Apoptosis induced by staurosporine alters chaperone and endoplasmic reticulum proteins: Identification by quantitative proteomics. Proteomics 2007, 7, 3085–3096. [Google Scholar]
- Murugan, A.K.; Bojdani, E.; Xing, M. Identification and functional characterization of isocitrate dehydrogenase 1 (IDH1) mutations in thyroid cancer. Biochem. Biophys. Res. Commun. 2010, 393, 555–559. [Google Scholar]
- Reitman, Z.J.; Yan, H. Isocitrate dehydrogenase 1 and 2 mutations in cancer: Alterations at a crossroads of cellular metabolism. J. Natl. Cancer Inst. 2010, 102, 932–941. [Google Scholar]
- Senthilnathan, P.; Padmavathi, R.; Magesh, V.; Sakthisekaran, D. Modulation of TCA cycle enzymes and electron transport chain systems in experimental lung cancer. Life Sci. 2006, 78, 1010–1014. [Google Scholar]
- Xu, S.G.; Yan, P.J.; Shao, Z.M. Differential proteomic analysis of a highly metastatic variant of human breast cancer cells using two-dimensional differential gel electrophoresis. J. Cancer Res. Clin. Oncol. 2010, 136, 1545–1556. [Google Scholar]
- Jung, K.H.; Park, J.W. Suppression of mitochondrial NADP(+)-dependent isocitrate dehydrogenaseactivity enhances curcumin-induced apoptosis in HCT116 cells. Free Radic. Res. 2011, 45, 431–438. [Google Scholar]
- Qi, Y.; Chiu, J.F.; Wang, L.; Kwong, D.L.; He, Q.Y. Comparative proteomic analysis of esophageal squamous cell carcinoma. Proteomics 2005, 5, 2960–2971. [Google Scholar] [Green Version]
- Klopfleisch, R.; Klose, P.; Weise, C.; Bondzio, A.; Multhaup, G.; Einspanier, R.; Gruber, A.D. Proteome of metastatic canine mammary carcinomas: similarities to and differences from human breast cancer. J. Proteome Res. 2010, 9, 6380–6391. [Google Scholar]
- Haley, B.; Paunesku, T.; Protic, M.; Woloschak, G.E. Response of heterogeneous ribonuclear proteins (hnRNP) to ionising radiation and their involvement in DNA damage repair. Int. J. Radiat. Biol. 2009, 85, 643–655. [Google Scholar]
- Honore, B.; Baandrup, U.; Vorum, H. Heterogeneous nuclear ribonucleoproteins F and H/H′ show differential expression in normal and selected cancer tissues. Exp. Cell Res. 2004, 294, 199–209. [Google Scholar]
- Balasubramani, M.; Day, B.W.; Schoen, R.E.; Getzenberg, R.H. Altered expression and localization of creatine kinase B, heterogeneous nuclear ribonucleoprotein F, and high mobility group box 1 protein in the nuclear matrix associated with colon cancer. Cancer Res. 2006, 66, 763–769. [Google Scholar]
- Zhao, W.; Fan, G.C.; Zhang, Z.G.; Bandyopadhyay, A.; Zhou, X.; Kranias, E.G. Protection of peroxiredoxin II on oxidative stress-induced cardiomyocyte death and apoptosis. Basic Res. Cardiol. 2009, 104, 377–389. [Google Scholar]
- Chen, Y.; Zhang, H.; Xu, A.; Li, N.; Liu, J.; Liu, C.; Lv, D.; Wu, S.; Huang, L.; Yang, S.; He, D.; Xiao, X. Elevation of serum l-lactate dehydrogenase B correlated with the clinical stage of lung cancer. Lung Cancer 2006, 54, 95–102. [Google Scholar]
- Thangaraju, M.; Carswell, K.N.; Prasad, P.D.; Ganapathy, V. Colon cancer cells maintain low levels of pyruvate to avoid cell death caused by inhibition of HDAC1/HDAC3. Biochem. J. 2009, 417, 379–389. [Google Scholar]
- Sullivan, D.C.; Huminiecki, L.; Moore, J.W.; Boyle, J.J.; Poulsom, R.; Creamer, D.; Barker, J.; Bicknell, R. EndoPDI, a novel protein-disulfide isomerase-like protein that is preferentially expressed in endothelial cells acts as a stress survival factor. J. Biol. Chem. 2003, 278, 47079–47088. [Google Scholar]
- Chay, D.; Cho, H.; Lim, B.J.; Kang, E.S.; Oh, Y.J.; Choi, S.M.; Kim, B.W.; Kim, Y.T.; Kim, J.H. ER-60 (PDIA3) is highly expressed in a newly established serous ovarian cancer cell line, YDOV-139. Int. J. Oncol. 2010, 37, 399–412. [Google Scholar]
- Pressinotti, N.C.; Klocker, H.; Schafer, G.; Luu, V.D.; Ruschhaupt, M.; Kuner, R.; Steiner, E.; Poustka, A.; Bartsch, G.; Sultmann, H. Differential expression of apoptotic genes PDIA3 and MAP3K5 distinguishes between low- and high-risk prostate cancer. Mol. Cancer 2009, 8, 130. [Google Scholar]
- Cho, S.; Dawson, P.E.; Dawson, G. Antisense palmitoyl protein thioesterase 1 (PPT1) treatment inhibits PPT1 activity and increases cell death in LA-N-5 neuroblastoma cells. J. Neurosci. Res. 2000, 62, 234–240. [Google Scholar]
- Cho, S.; Dawson, P.E.; Dawson, G. Role of palmitoyl-protein thioesterase in cell death: Implications for infantile neuronal ceroid lipofuscinosis. Eur. J. Paediatr. Neurol. 2001, 5, S53–S55. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Su, C.-C.; Su, J.-H.; Lin, J.-J.; Chen, C.-C.; Hwang, W.-I.; Huang, H.H.; Wu, Y.-J. An Investigation into the Cytotoxic Effects of 13-Acetoxysarcocrassolide from the Soft Coral Sarcophyton crassocaule on Bladder Cancer Cells. Mar. Drugs 2011, 9, 2622-2642. https://doi.org/10.3390/md9122622
Su C-C, Su J-H, Lin J-J, Chen C-C, Hwang W-I, Huang HH, Wu Y-J. An Investigation into the Cytotoxic Effects of 13-Acetoxysarcocrassolide from the Soft Coral Sarcophyton crassocaule on Bladder Cancer Cells. Marine Drugs. 2011; 9(12):2622-2642. https://doi.org/10.3390/md9122622
Chicago/Turabian StyleSu, Ching-Chyuan, Jui-Hsin Su, Jen-Jie Lin, Cheng-Chi Chen, Wen-Ing Hwang, Han Hsiang Huang, and Yu-Jen Wu. 2011. "An Investigation into the Cytotoxic Effects of 13-Acetoxysarcocrassolide from the Soft Coral Sarcophyton crassocaule on Bladder Cancer Cells" Marine Drugs 9, no. 12: 2622-2642. https://doi.org/10.3390/md9122622
APA StyleSu, C. -C., Su, J. -H., Lin, J. -J., Chen, C. -C., Hwang, W. -I., Huang, H. H., & Wu, Y. -J. (2011). An Investigation into the Cytotoxic Effects of 13-Acetoxysarcocrassolide from the Soft Coral Sarcophyton crassocaule on Bladder Cancer Cells. Marine Drugs, 9(12), 2622-2642. https://doi.org/10.3390/md9122622