1. Introduction
Fucose-containing sulfated polysaccharides (FCSPs) designate a group of diverse polysaccharides that can be extracted from brown seaweeds of the class Phaeophyceae. This seaweed class includes the order Fucales, in which seaweed species such as
Fucus sp. and
Sargassum sp. belong. The most studied FCSPs, originally called fucoidin, fucoidan or just fucans, have a backbone built of (1→3)-linked α-L-fucopyranosyl residues or of alternating (1→3)- and (1→4)-linked α-L-fucopyranosyl residues [
1,
2]. These fucopyranosyl residues may be substituted with short fucoside side chains or sulfate groups at C-2 or C-4, and may also carry other minor substitutions, e.g., acetate, xylose, mannose, glucuronic acid, galactose, or glucose [
3,
4,
5]. Brown seaweed FCSPs also include sulfated galactofucans with backbones built of (1→6)-β-D-galacto- and/or (1→2)-β-D-mannopyranosyl units. In addition to sulfate these backbone residues may be substituted with fucosides, single fucose substitutions, and/or glucuronic acid, xylose or glucose substitutions [
4]. Recently it has been understood that the compositional and structural features of FCSPs differ significantly among seaweed species and that these features are markedly influenced by the conditions used to extract them [
3,
6].
FCSPs of different degrees of purity and composition, extracted from brown seaweeds such as
Sargassum sp. and
Fucus sp., have been documented to have a wide range of biological activities including anticoagulant [
7,
8], antithrombotic [
8], anti-inflammatory [
9], anti-viral [
10,
11]; and notably anti-tumoral effects [
8,
12,
13]. Unfractionated FCSPs have thus specifically been found to reduce cell proliferation of lung carcinoma and melanoma cells
in vitro; to exert immunopotentiating effects in tumor bearing animals; and to activate natural killer cells in mice leading to increased anti-tumor effectiveness [
13,
14,
15,
16]. Kim
et al. [
17] applied a crude polysaccharide composed predominantly of sulfated fucose from
Fucus vesiculosus to human colon cancer cells
in vitro, and concluded that this crude brown seaweed polysaccharide extract can induce apoptosis, and provided data that suggested that the apoptosis was induced via activation of caspases. Moreover, commercially available crude FCSPs (“fucoidan”) extracted from
F. vesiculosus have been reported to inhibit proliferation and induce apoptosis on human lymphoma HS-Sultan cells lines by activation of caspase-3 [
18]. Recently, we have reported that crude FCSPs extracted from a
Sargassum sp. and from
F. vesiculosus, respectively, induce growth inhibition and apoptosis of melanoma B16 cells
in vitro [
13]. When injected intraperitoneally into mice over four days, these same unfractionated FCSPs were found to induce enhanced natural killer cells (NK cells) activity to result in specific lysis of YAC-1 cells (a murine T-lymphoma cell line sensitive to NK cells) [
13]. Previous reports with human HS-Sultan cells and MCF-7 cells, respectively, have suggested that the FCSPs induced apoptosis initiation may take place via activation of caspase-3 and caspase-8 dependent pathways, respectively [
18,
19], but no firm evidence has been established regarding the exact mechanism responsible for the apoptotic action of the FCSPs. The objective of the present study was, therefore, to examine whether the anti-proliferative action and apoptosis of melanoma B16 cells induced by FCSPs derived from
Sargassum henslowianum C. Agardh and
Fucus vesiculosus, are accompanied by increased caspase-3 activity. We also wanted to evaluate whether any structural features of the FCSPs might be crucial for bioactivity. In this study, we present the different structural features of the FCSPs derived from
S. henslowianum and
F. vesiculosus as assessed by IR and
1H NMR spectroscopy and show that these FCSPs exert bioactive effects that inhibit the proliferation of melanoma B16 cells by apoptosis. We also show that the antiproliferative effects and the apoptosis are accompanied by activation of caspase-3.
3. Discussion
The incidence of melanoma skin cancer has risen dramatically over the past few decades [
27]. Because of the significant risk and undesirable effects of known cancer therapeutic strategies, many studies have evaluated the possible protective effects of bioactive compounds of natural origin. Fucose-containing sulfated polysaccharides (FCSPs) derived from naturally grown brown seaweeds by aqueous extraction have been shown to exert potentially beneficial bioactivities, including immuno-modulatory, anti-inflammatory and anti-tumorigenic effects. In keeping its natural properties, FCSPs must be extracted from brown seaweeds by use of a mild processing treatment and a minimal number of extraction steps.
Brown seaweeds constitute a part of the conventional diet in several Asian countries, especially in Japan, and in a Japanese cohort study the intake of seaweeds has been associated with lower mortality from all chronic diseases including cancer [
28]. It has recently been demonstrated that FCSPs from brown seaweeds exert growth inhibitory activity on certain cancer cell lines
in vitro [
13,
17,
18]. Incorporation of brown seaweeds into animal diets has also revealed cancer inhibitory effects with no direct lethal consequences [
29,
30]. Natural FCSPs from brown seaweeds may therefore have significant potential as protective agents to control or prevent skin cancer provided that the FCSPs do indeed exert cancer-preventive effects.
In this study, in accordance with previous data [
13], we found that unfractionated FCSPs,
i.e., FSAR and FVES, extracted from the brown seaweeds
S. henslowianum and
F. vesiculosus, respectively, were composed of fucose, galactose, xylose, mannose and glucuronic acid, and showed that the fucose, galactose and glucuronic acid contents differed significantly among the two FCSPs products, but that their sulfate contents were similar (
Table 1).
We also found both distinct differences and several similarities in the structural make-up of these FCPSs by use of FTIR and
1H NMR spectroscopy. The FT-IR analyses thus corroborated the presence of sulfate groups in both the FSAR and the FVES sample (
Figure 1). The IR spectra indicated that the sulfate substitutions of the FCSPs extracted from the
Sargassum sp. (FSAR) were located in the equatorial C-2 and/or C-3 positions as depicted by absorption bands at 817 cm
−1. This finding was in agreement with data reported for fucoidan fractions isolated from
Sargassum stenophyllum [
4]. However, Duarte
et al. [
4] also reported that two other saccharide fractions from
S. stenophyllum had an absorption band at 837 cm
−1 indicating sulfate groups at the C-4 positions of the structural monosaccharides [
4]. The spectra of the FCSPs from
F. vesiculosus (FVES) displayed an absorption band at 838 cm
−1 with a small shoulder at ~822 cm
−1 indicating sulfate groups at both the C-4 and the C-2 position (
Figure 1). This finding corresponds to previously reported
1H NMR data of FCSPs from
F. vesiculosus that have indicated a typical structure of algal fucoidan consisting of α3-linked 2-mono-
O-sulfated L-fucopyranose residues, and/or α3-linked 2,4-di-
O-sulfated L-fucopyranose residues [
2,
26]. Small disparities in the IR spectra from different published reports can be due to factors such as sample handling and the FCSPs extraction procedure employed.
The present study also aimed at establishing whether crude FCSPs extracted from
Sargassum henslowianum C. Agardh (FSAR) contained fucoidan-like structures composed of α-3-linked or/and α-3,4-linked L-fucopyranose residues. Even though signals consistent with the presence of α-L-fucopyranose entities were recorded (with
1H NMR signals at 5.10 and 5.18 ppm,
Figure 2a), the probability that the FSAR may contain a cocktail of polysaccharides is likely. Hence, the
1H NMR spectra also showed that the FSAR sample contained 3-linked D-galactopyranose residues as indicated by an independent signal at 4.61 ppm (
Figure 2a). β-(1→)3-linked galactopyranose residues are known to be a typical structural feature of seaweed polysaccharides, from e.g.,
Laminaria angustata var.
longissima,
Botryocladia occidentalis [
25,
31]. However, another possibility might be that the FSAR sample was not composed of a mixture of different types of polysaccharides but rather, that the sample consisted of one type of a highly complex hetero-polysaccharide as suggested by Duarte
et al. [
4] for the fucoidans from
Sargassum stenophyllum. It can safely be said that the
1H NMR spectra of the FCSPs samples were complex and overlapping. It is therefore difficult to draw any definite conclusions about the detailed structural features and differences among the two FCSPs; the detailed elucidations of the definite structural details were also beyond the scope of this present study, but clearly deserve further investigation. Nonetheless, the data confirmed that the diversity,
i.e., the compositional and structural complexity of (potentially bioactive) algal fucose-containing sulfated polysaccharides, is much wider than originally believed.
The biological activities of the FCSPS against skin cancer cells were investigated
in vitro, and the results revealed that both FSAR and FVES can exert anti-proliferative effects on melanoma B16 cells
in vitro. The FSAR sample induced more significant reductions of the cell viability of melanoma cells than the FVES sample at low dosage levels (
Figure 3). At higher dosages, the FSAR treatment still induced gradually more loss of cell viability, but the FVES had more potent anti-proliferative effects at higher dosages than FSAR which could indicate direct cell toxicity. The bioactivities of these FCSPs may be attributable to their distinct structural features, notably the level of sulfation (charge density), the distribution (e.g., random
versus clustered) and bonding of the sulfate substitutions, as well as other specific structural features of the sulfated fucans and the sulfated galactofucan complexes. The sulfate groups of FVES were substituted at the C-2 and C-4 position of the fucose substituents, typical for fucoidan from
F. vesiculosus, and consistent with previously published data indicating that the sulfate groups were substituted at C-2 of α3-linked L-fucopyranose residues in fucoidan from e.g.,
Fucus evanescens [
21]. In contrast, the sulfate substitutions in the FSAR were interpreted to be mainly at the C-2 and/or C-3 positions of the monosaccharides according to the IR spectra (
Figure 1); the observation of C-2 linked sulfate groups agreed with the data mentioned above for fucoidan from
Fucus spp. [
3,
21], but is also in agreement with the proposition that the sulfate groups were substituted at C-2 on the 3-linked galactopyranose residues [
25]. The possible presence of sulfated, 3-linked galactan in the structure of FSAR may contribute to the efficacy of FSAR to induce anti-proliferative effects as it has been reported that 2-
O-sulfated 3-linked galactan is more bioactive than 2-
O-sulfated 3-linked fucans and 3-
O-sulfated 4-linked galactan [
32,
33,
34].
The findings that the FCSPs induced apoptosis of the melanoma B16 cells
in vitro were in agreement with recent reports [
13,
16,
35], but the differential apoptotic efficacies, and the dose-response effects of differently structured FCSPs (
Figure 4) have not been reported earlier. In particular, it is a novel finding that significantly different sulfated, polysaccharide structures from brown seaweeds—as evaluated in the present work—exert relatively similar apoptotic effects on melanoma cells. The results of this work thus indicate that not only the well-studied, classical type of FCSPs having a backbone made up of (1→3)-linked α-L-fucopyranosyl or of alternating (1→3)- and (1→4)-linked α-L-fucopyranosyl residues have potential tumor-preventing effecs, but also that the more complex sulfated fucose-rich galacto-mannans from
Sargassum spp. exert promising cancer-preventive effects. The principal objective of this study was to assess whether any structural features of the FCSPs might be crucial for bioactivity, and the data suggest that the sulfate substitutions, and not necessarily only the fucose-backbone structure itself, confer this decisive bioactivity. It is however important to investigate whether other differently structured FCSPs may exert similar growth inhibitory and apoptosis inducing effects on cancer cells.
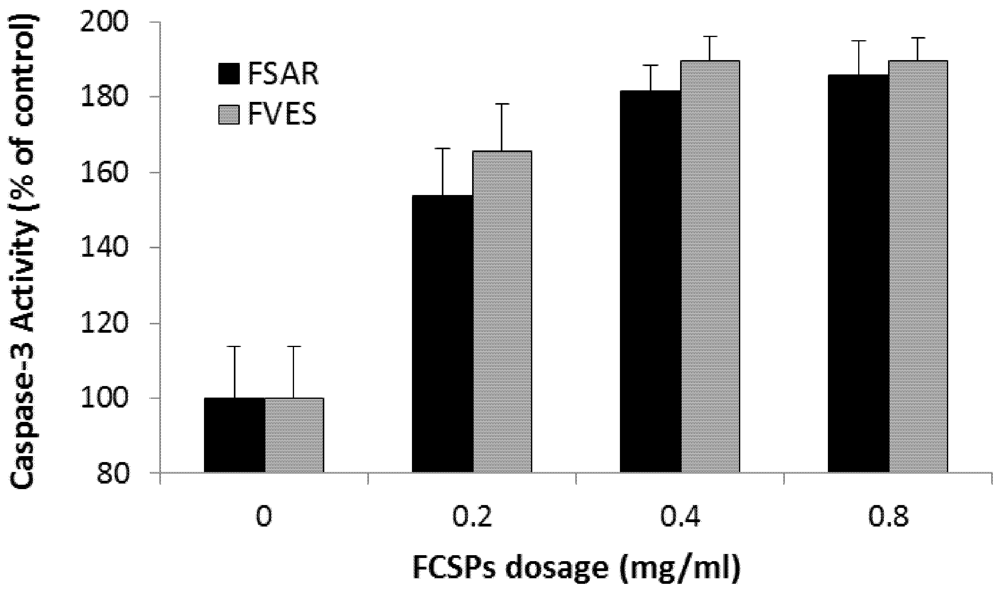
In this work we noted that both FCSPs activate caspase-3 in a dosage-response fashion (
Figure 5). These findings affirmed the results reported previously which have shown that FCSPs (“fucoidan”) from
F. vesiculosus induce apoptosis in human lymphoma HS-Sultan cell lines and in HT-29 and HCT116 human colon cancer cells
in vitro, and moreover that the exposure of these cells to the
F. vesiculosus FCSPs appear to activate caspase-3 [
17,
18]. The
F. vesiculosus FCSPs treatment was also shown to enhance mitochondrial membrane permeability of human colon cancer cells
in vitro, and to induce cytochrome c and Smac/Diablo release from the mitochondria [
17]. It has also been reported that pretreatment of HT-29 and HCT116 colon cancer cells with individual caspase-8 or caspase-9 inhibitors (Z-IETD-FMK and Z-LEHD-FMK, respectively) prior to fucoidan exposure reduced the levels of caspases, including caspase-3 [
17]. It has likewise been shown that pretreatment of human lymphoma HS Sultan cells with a pan-caspase inhibitor, z-VAD-FMK, reduced fucoidan-induced apoptosis [
18]. Hence, the available data support the proposition that fucoidan-induced apoptosis occurs through caspase activation pathways. The cascade mechanism by which the caspase-activation is presumed to take place via the mitochondria-mediated apoptotic pathway is illustrated in
Figure 6.
Loss of plasma membrane is one of the earliest features of apoptosis and Annexin V staining can identify apoptosis at an early stage. However this assay does not distinguish between cells that have undergone apoptotic death
versus those that have died as a result of a necrotic pathway, because in either case the dead cells will stain with both Annexin V and 7-AAD. Both early and later apoptosis stages were observed by the FACS scanning indicating that the FCSPs had a direct apoptotic effect on the melanoma B16 cells (
in vitro) (
Figure 4). The direct apoptotic action of the FCSPs was probably due to the interaction of the highly negative charge density of the FCSPs with the melanoma B16 cells (as a result of the sulfation). Recently, we reported that crude fucoidan from
Sargassum sp. could trigger apoptosis indirectly by enhancing the activity of natural killer (NK) cells activity
in vivo [
13]. NK cells produce immunologically important cytokines, notably IFN-γ, which can promote the activation of T-cells to produce interleukin-2 and -12 that in turn further enhance the NK cell activation [
14,
36].
Figure 6.
Proposed mechanism for inhibition of the proliferation of melanoma cells by FCSPs: Activation of macrophages via membrane receptors, which leads to the production of cytokines that enhance NK cell activation. Activated NK cells release Granzyme B and perforin through granule exocytosis into the space between NK cells and melanoma cells to initiate caspase cascades in melanoma cells. Assimilation of Granzyme B by the tumor cells is facilitated by perforin. Granzyme B then initiates apoptosis by triggering the release of mitochondrial cytochrome c and apoptosome formation leading to caspase-3 activation, which in turn translocates the nucleus causing DNA fragmentation—the distinct morphological change of cells by apoptosis [
36,
37].
Figure 6.
Proposed mechanism for inhibition of the proliferation of melanoma cells by FCSPs: Activation of macrophages via membrane receptors, which leads to the production of cytokines that enhance NK cell activation. Activated NK cells release Granzyme B and perforin through granule exocytosis into the space between NK cells and melanoma cells to initiate caspase cascades in melanoma cells. Assimilation of Granzyme B by the tumor cells is facilitated by perforin. Granzyme B then initiates apoptosis by triggering the release of mitochondrial cytochrome c and apoptosome formation leading to caspase-3 activation, which in turn translocates the nucleus causing DNA fragmentation—the distinct morphological change of cells by apoptosis [
36,
37].
The apoptosis induced by FCSPS via the activation of caspase-3 was reported previously to be mediated through a mitochondrial pathway [
17,
18,
19,
38]. However, it remains to be determined whether differences in FCSPs structures will influence the apoptotic mechanism, including the mitochondrial pathway apoptosis cascade. The route of the mitochondrially dependent apoptotic pathway is the release of apoptosis-inducing factor (AIF) and cytochrome
c from the inner mitochondrial membrane into the cytosol. Cytochrome
c interacts with Apaf-1 (apoptotic protease activating factor 1) and procaspase-9 to form the active apoptosome. The apoptosome then initiates the cleavage of procaspase-3, producing active caspase-3, which initiates the execution phase of apoptosis by proteolysis of substances whose cleavage commits the cell to apoptosis [
39] (
Figure 6). The influence of the different FCSPs structures on the mitochondrial membrane permeability and electric potential requires further study. We hope in the future to investigate the bioactivity and mechanism of FCSPs on certain degenerative diseases
in vivo and to further elucidate specific molecular targets of FSCPs for inhibition of cancer cells.
4. Experimental Section
4.1. Chemicals
Dried
S. henslowianum C. Agardh was obtained from Viet Delta Ltd. (Ho Chi Menh, Vietnam) and the Fucose-containing sulfated polysaccharides (FCSPs) from the
S. henslowianum (FSAR) were extracted in our laboratory (see below). Crude fucoidan from
F. vesiculosus (FVES) was obtained from Sigma-Aldrich (Steinheim, Germany); according to the product description the FVES had been prepared from
F. vesiculosus via the extraction method described by Black and Dewar [
40]. Hydrochloric acid (37%), D-glucose and D-xylose were purchased from Merck (Darmstadt, Germany). Trifluoracetic acid (99%, TFA), trichloroacetic acid (99%, TCA), CaCl
2, Na
2SO
4, BaCl
2, arabinose, rhamnose, D-galactose and L-fucose were from Sigma–Aldrich Co. (Steinheim, Germany). Agarose D-2 was obtained from Hispanagar (Burgos, Spain). Caspase-3 colorimetric assay kit was obtained from Biovision, Inc. (Mountain View, CA. USA). Minimal essential medium eagle (MEM-eagle) cell culture media was purchased from Sigma–Aldrich Co. (Steinheim, Germany); foetal bovine serum (FBS) was from Flow Laboratories (North Ryde, N.S.W., Australia); streptomycin–penicillin and Trypan Blue were from Gibco (Canada). Cell Proliferation Kit 1 was obtained from Roche Applied Science, Germany. The PE Annexin V Apoptosis Detection Kit 1 was obtained from BD Biosciences (Franklin Lakes, NJ, USA). All chemicals used were analytical grade.
4.3. Acid Hydrolysis and FCSPs Composition Analysis
The freeze dried FSAR and FVES samples (20 mg) were hydrolyzed separately in 2 M TFA (final concentration) at 121 °C for 2 h, then the hydrolyzed mixture were freeze dried at −57 °C (Heto Lyolab 3000, England). Each dried powder sample was resolubilized in doubly distilled water and centrifuged at 10,000 rpm for 10 min to collect the supernatant (Sigma Laboratory Centrifuge 4K15, VWR, Denmark). Each supernatant was filtered through a 0.2 µm syringe tip filter (SUN-Sri, Rockwood, TN) prior to injection into the HPAEC-PAD for monosaccharide analysis [
41]. Analysis of sulfate content was done according to the method described by Jackson and McCandless [
42].
4.4. 1H NMR and FTIR Spectroscopy
The 1H NMR spectra were obtained using an INOVA 600 NMR spectrometer (Agilent Technologies Japan, Ltd., Tokyo Japan) equipped with a 1H[15N-31P] pulse field gradient indirect-detecting probe. Standard pulse sequences were used in all operations. The 1H chemical shift (δH) was referenced to HOD (δH 4.76 ppm, 2H2O). The 1H NMR spectrum was assigned through the 1H–1H decoupling technique. An NMR spectrum of L-fucose was utilized as a reference for chemical shift assignment. The lyophilized FCSPs powders were dissolved in deuterium oxide (2H2O) and evaporated to exchange the unstable 1H with 2H. The evaporation and dissolution step was repeated five times, and the samples (10 mg) were finally dissolved in 0.75 mL 2H2O and then subjected to NMR spectroscopy. The IR spectra were obtained using a Spectrum One FT-IR spectrometer (Perkin Elmer, Waltham, MA, USA) equipped with universal attenuated total reflectance (UATR) accessories. Analysis of each of the FSAR and FVES powders, ~1 mg of each, was done using diffuse reflectance infrared transform spectroscopy (DRIFTS) and the spectrum was evaluated by Perkin Elmer Spectrum software version 5 (Perkin Elmer, Waltham, MA, USA).
4.5. Cell Culture and Anti-Proliferative Assay
Melanoma B16 cells (MC) were grown in MEM eagle medium supplemented with 10% (v/v) heat inactivated FBS, 1% (w/v) streptomycin–penicillin and 1% (v/v) of 200 mM L-glutamine at 37 °C under 5% CO2. Monolayer cultivation was carried out by adding 100 µL of the cell-MEM-FBS mixture into separate wells in 96-flat well plates at a density of 6 × 104 cells per well followed by incubation for 24 h in 5% CO2 at 37 °C. For the anti-proliferation assay the medium was removed after the 24 h of monolayer cell cultivation and replaced with 100 µL of MEM medium containing 2% FBS and varying concentrations (0.1–1.0 mg/mL) of the crude FCSPs, i.e., FSAR and FVES, respectively, and the mixtures were then incubated for 24 h. Quantification of cell proliferation was carried out using a tetrazolium salt (MTT (3-(4,5-dimethyl-thiazolyl-2)-2,5-diphenyltetrazolium bromide)) based colorimetric assay following the protocol supplied with the Cell Proliferation Kit 1 (Roche Applied Science, Germany). Briefly, 20 µL MTT solution (5 mg/mL) was added to the cell cultures after the 24 h of incubation with the FCSPs, and the cell cultures were then re-incubated for 4 h. Finally, 100 µL of stabilization solution was added to each well and the plates were incubated overnight at 37 °C under 5% CO2. Absorbance was measured using an Elisa reader at 550–690 nm.
4.6. Cell Culture and Caspase-3 Assay
Melanoma B16 cells (MC) were grown in MEM eagle culture medium supplemented with 10% (v/v) heat inactivated FBS, 1% (w/v) streptomycin–penicillin and 1% (v/v) of 200 mM L-glutamine maintained at 37 °C under 5% CO2. For the caspase-3 assay, monolayer cultivation was carried out in a petri dish (60 × 15 mm) by adding 5 mL culture medium containing melanoma cells at a density of 1 × 105 per mL and varying concentrations (0.2, 0.4 and 0.8 mg/mL) of the FSAR and FVES, respectively. The mixtures were then incubated for 24 and 48 h in 5% CO2 at 37 °C. The caspase-3 assay was performed according to the protocol supplied with the assay kit (Biovision Inc., Mountain View, CA, USA) used to assay the activity of caspases that recognize the amino acid sequence DEVD. The assay was based on spectrophotometric detection of the chromophore p-nitroaniline (pNA) after cleavage from the labeled substrate DEVD-pNA. Concisely, the melanoma B16 cells exposed to FSAR and FVES, respectively, were harvested and resuspended in 50 µL of cell lysis buffer and incubated on ice for 10 min. and the mixture centrifuged for 1 min (14,000 × g, 4 °C). Each supernatant was then transferred to a fresh tube, and reaction buffer (50 µL) and 4 mM DEVD-pNA substrate (5 µL) were added, and this reaction mixture was then incubated at 37 °C for 1 h. Absorbance of pNA light emission was quantified using a microtiter plate reader at 405 nm.
4.7. Apoptosis Assay by Fluorescence-Activated Cell Sorting (FACS)
After 24 h of monolayer cultivation of melanoma B16 cells with 0.2 mg/mL of FSAR or FVES, and no FCSPs addition as control, the culture medium was removed, and the cells harvested by addition of 1 mL Trypsin-EDTA. The harvested cells were washed twice with 0.1 M PBS and then resuspended in binding buffer according to the protocol for the Annexin V Apoptosis Detection Kit I (BD Biosciences, Franklin Lakes, NJ, USA). 100 µL of this solution at 1 × 105 cells was transferred to a culture tube and 5 µL of Annexin V and 5 µL of 7-amino-actinomycin (7-ADD) were added, and the mixture incubated at room temperature for 25 min. Then, 400 μL of binding buffer was added and the extent of apoptosis and staining pattern of the cells were tracked by flow cytometric analysis on a FACScan instrument (Becton Dickinson).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





