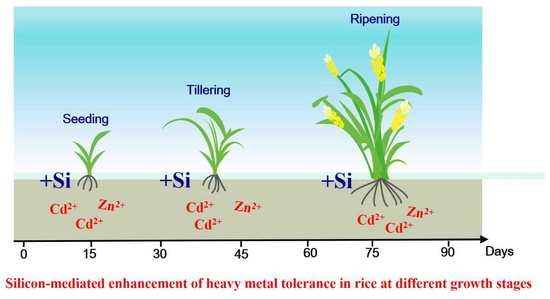
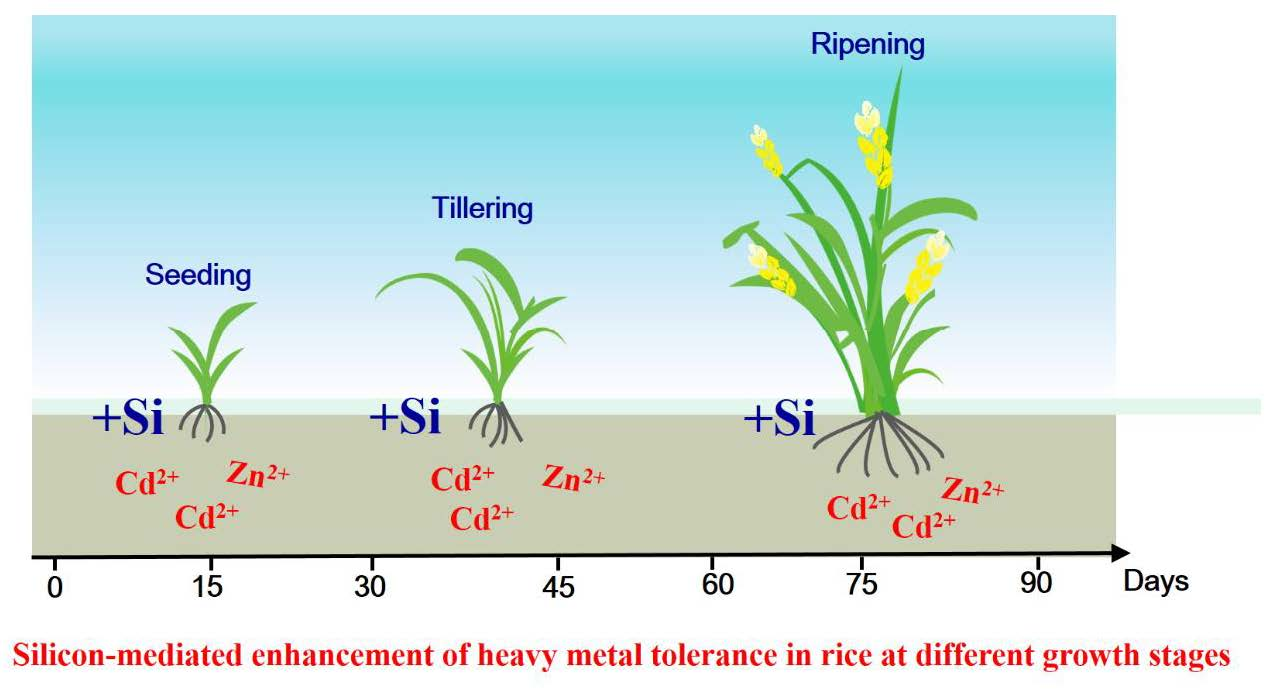
Silicon-Mediated Enhancement of Heavy Metal Tolerance in Rice at Different Growth Stages


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Soil Sample and Experimental Treatments
2.2. Chemical Analysis of Plant Tissues
2.3. Biochemical Analysis
2.3.1. Electrolyte Leakage Measurement and MDA Content
2.3.2. Total Chlorophyll Content and Chlorophyll Fluorescence Parameters
2.3.3. Anti-Oxidative Enzyme Activities
2.4. Statistical Analysis
3. Results
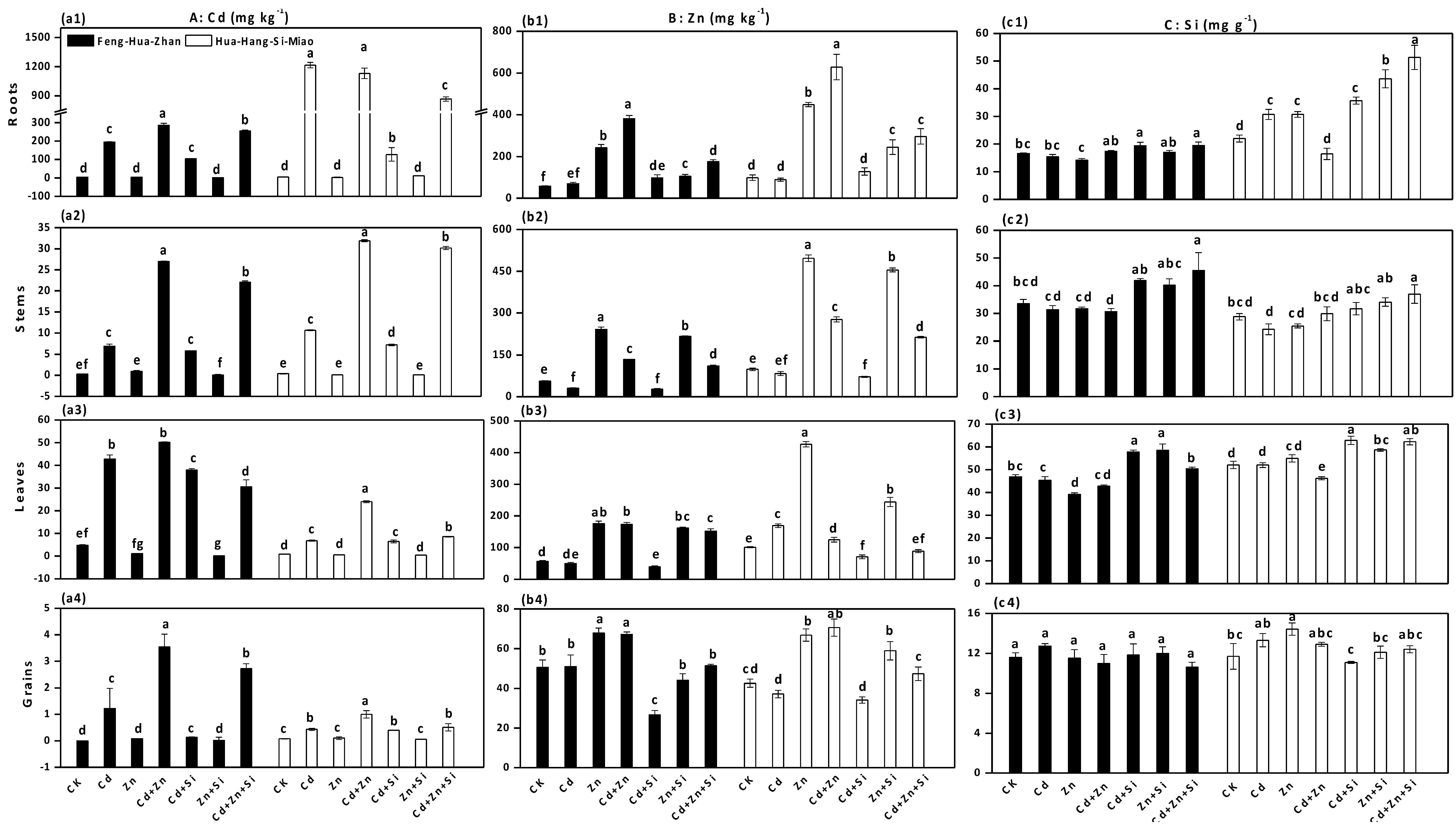
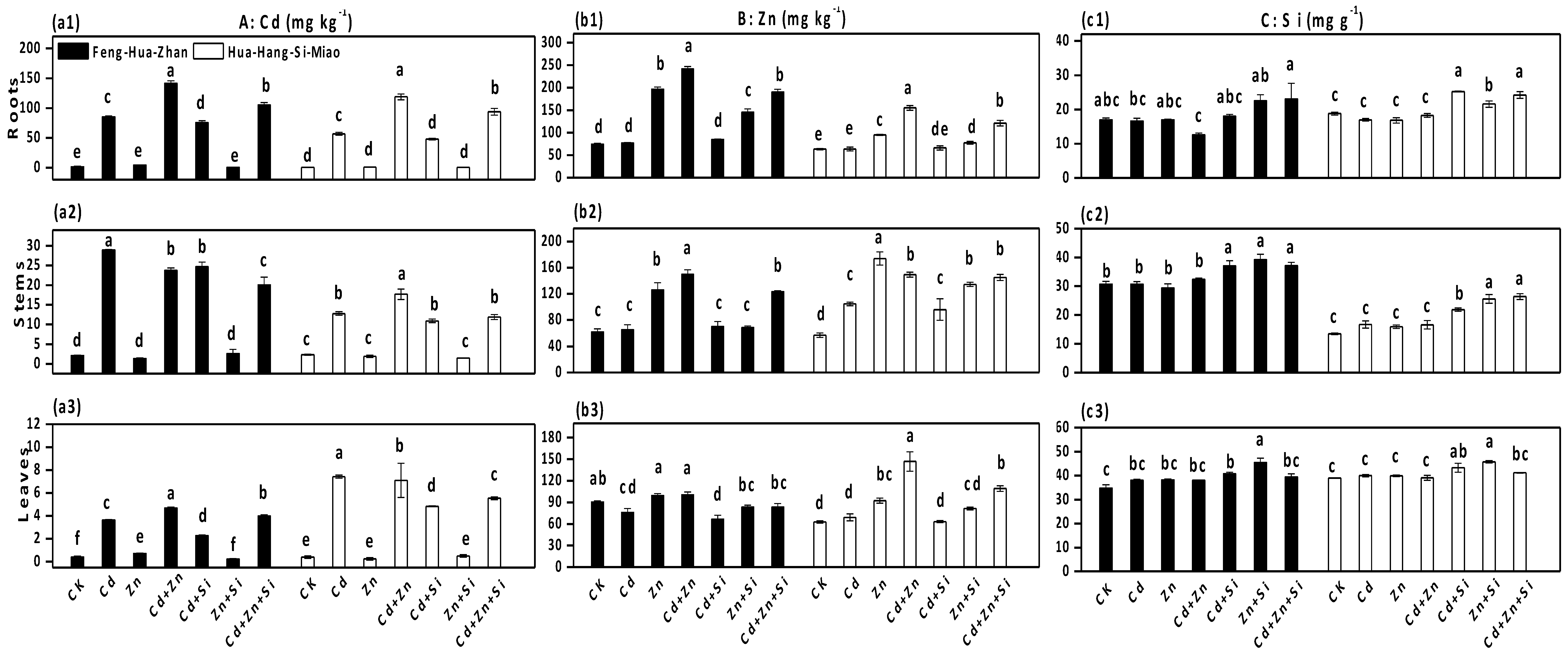
3.1. Effect of Si on Plant Growth
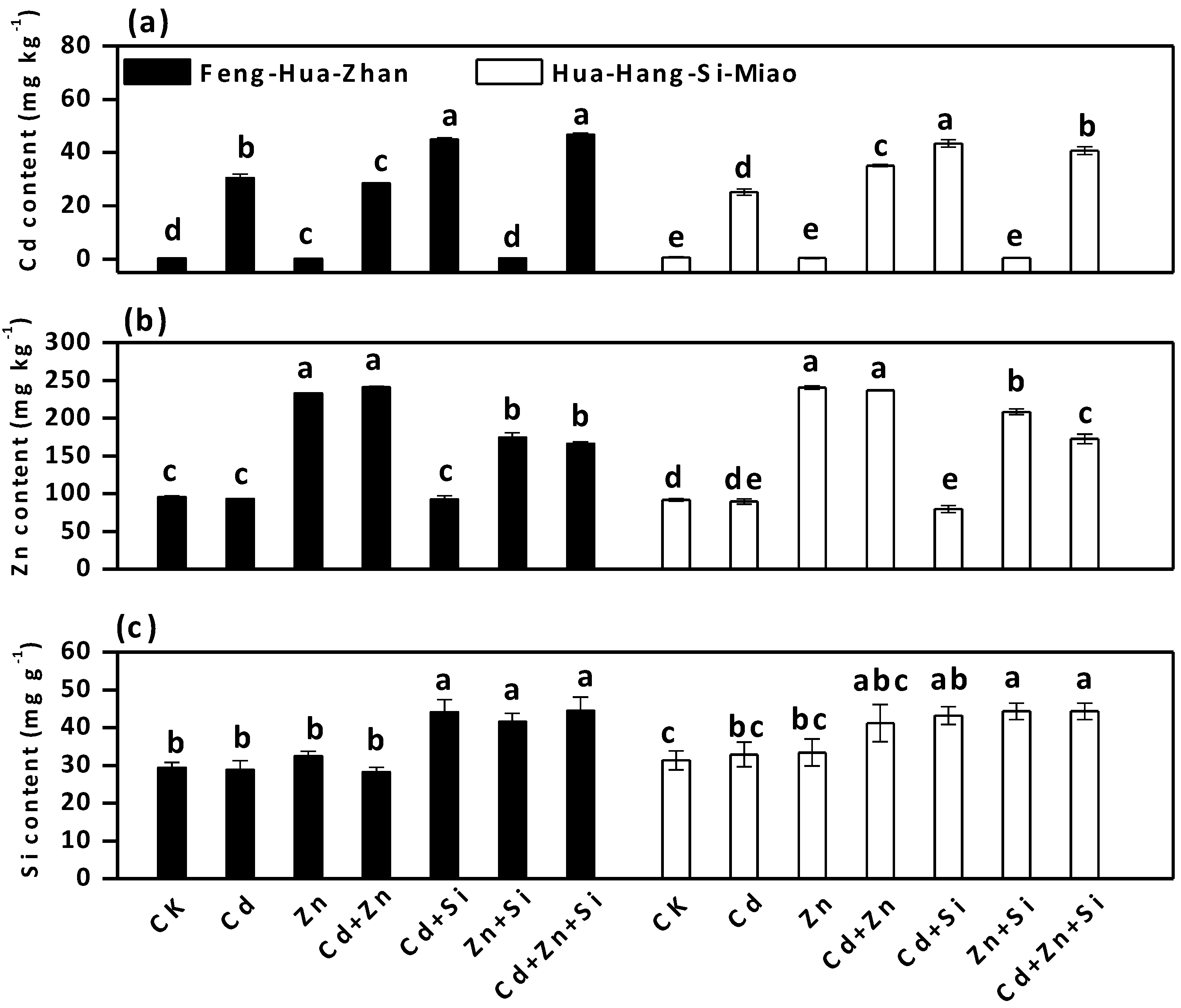
3.2. Effect of Si on Cd and Zn Accumulation in Rice Plants
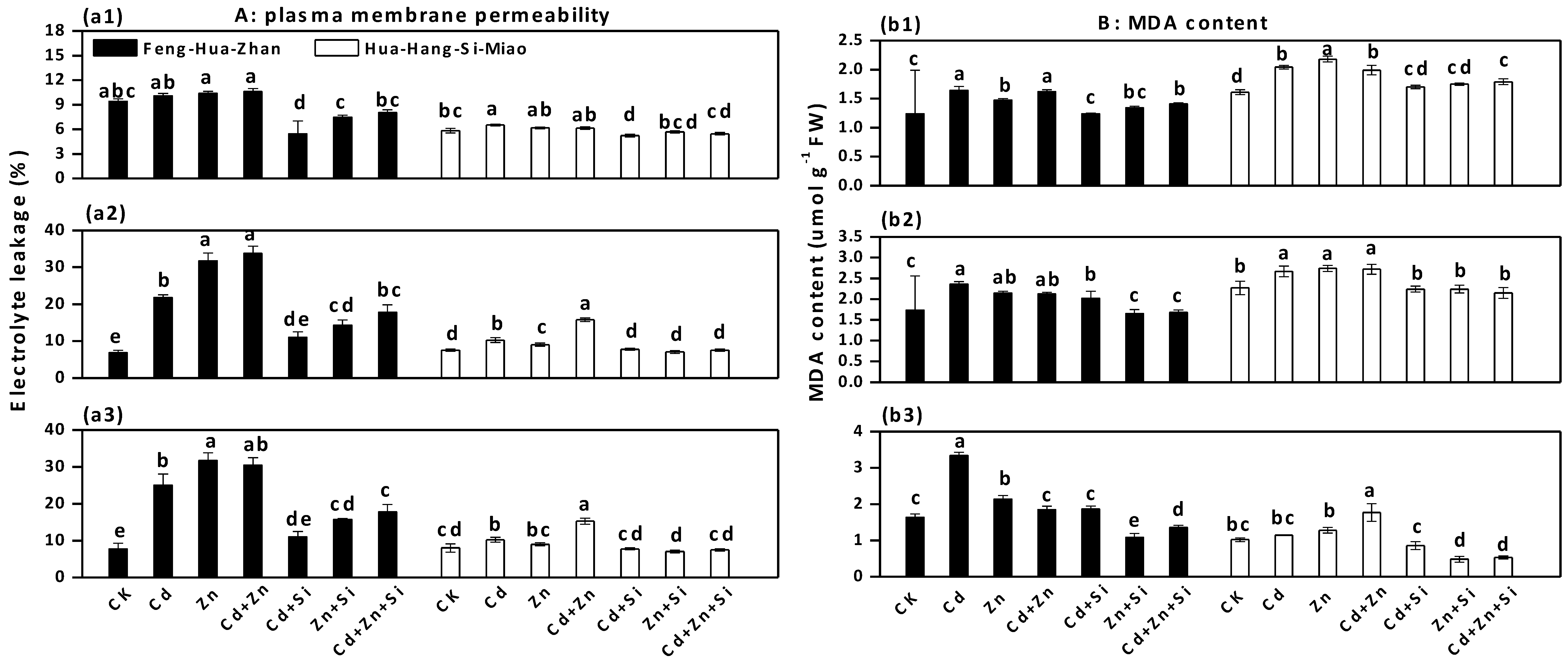
3.3. Effect of Si on Plasma Membrane Permeability and MDA Content
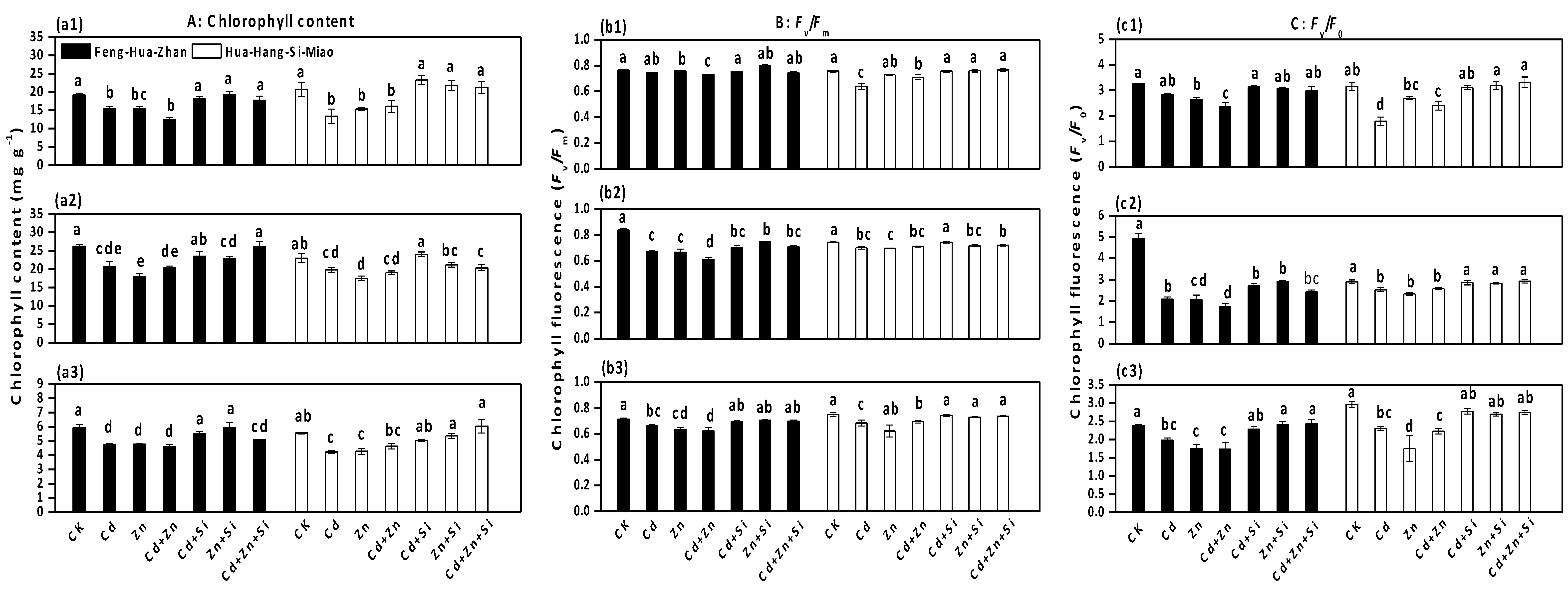
3.4. Effect of Si on Total Chlorophyll Content and Chlorophyll Fluorescence Parameters
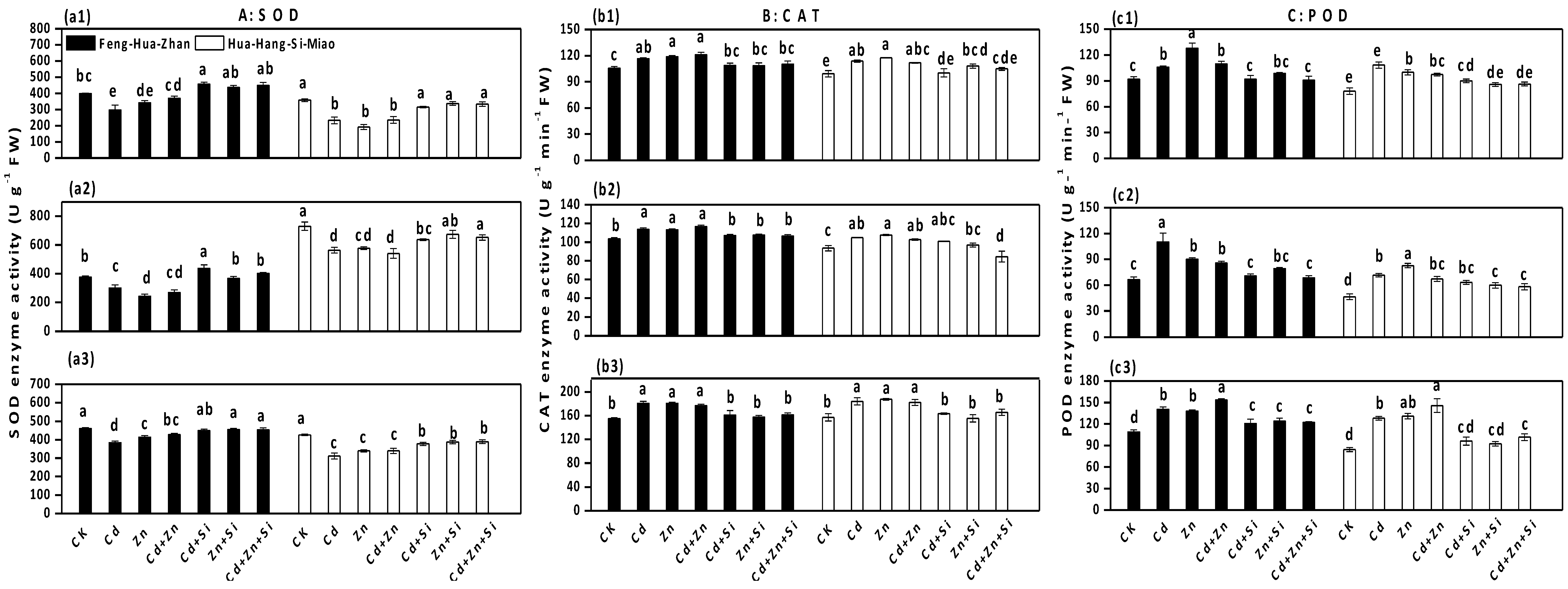
3.5. Effect of Si on Antioxidant Enzymes Activities
3.6. Effect of Si on the Cd and Zn Concentrations in Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Ovečka, M.; Takáč, T. Managing heavy metal toxicity stress in plants: Biological and biotechnological tools. Biotechnol. Adv. 2014, 32, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Linguaa, G.; Franchinb, C.; Todeschinia, V.; Castiglione, S.; Biondi, S.; Burlando, B.; Parravicini, V.; Torrigiani, P.; Berta, G. Arbuscularmycorrhizal fungi differentially affect the response to high zinc concentrations of two registered poplar clones. Environ. Pollut. 2008, 153, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E. Silicon. Annu. Rev. Plant Physiol. Mol. Boil. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.H.; Zhan, S.S.; Wang, S.Z.; Tang, Y.T.; Chaney, R.L.; Fang, X.H.; Cai, X.D.; Qiu, R.L. Silicon-mediated amelioration of zinc toxicity in rice (Oryze sativa L.) seedlings. Plant Soil 2012, 350, 193–204. [Google Scholar] [CrossRef]
- Li, P.; Wang, X.; Zhang, T.; Zhou, D.; He, Y. Effects of several amendments on rice growth and uptake of copper and cadmium from a contaminated soil. J. Environ. Sci. 2008, 20, 449–455. [Google Scholar] [CrossRef]
- Ma, J.; Cai, H.; He, C.; Zhang, W.; Wang, L. A hemicellulose-bound form of silicon inhibits cadmium ion uptake in rice (Oryza sativa) cells. New Phytol. 2015, 206, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.P.; Deng, J.W.; Zhang, H.M.; Ma, H.Y.; Cao, D.J.; Ma, R.X.; Liu, R.J.; Liu, C.; Liang, Y.G. Effects of cadmium on bioaccumulation and biochemical stress response in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2015, 122, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Wong, J.W.C.; Wei, L. Silicon-mediated enhancement of cadmium tolerance in maize (Zea mays L.) grown in cadmium contaminated soil. Chemosphere 2005, 58, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Hussain, I.; Ashraf, M.A.; Rasheed, R.; Asghar, A.; Sajid, M.A.; Iqbal, M. Exogenous application of silicon at the boot stage decreases accumulation of cadmium in wheat (Triticumaestivum L.) grains. Braz. J. Bot. 2015, 38, 223–234. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Nwugo, C.C.; Huerta, A.J. Silicon-induced cadmium resistance in rice (Oryza sativa). J. Plant Nutr. Soil Sci. 2008, 171, 841–848. [Google Scholar] [CrossRef]
- Nwugo, C.C.; Huerta, A.J. Effect of silicon nutrition on cadmium uptake, growth and photosynthesis of rice plants exposed to low-level cadmium. Plant Soil 2008, 311, 73–86. [Google Scholar] [CrossRef]
- Nwugo, C.C.; Huerta, A.J. The effect of silicon on the leaf proteome of rice (Oryza sativa L.) plants under cadmium-stress. J. Proteome Res. 2011, 10, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Fang, C.; Li, Y.; Lin, W.; He, J.; Lin, R.; Lin, W. Cadmium-stress mitigation through gene expression of rice and silicon addition. Plant Growth Regul. 2016, l, 1–11. [Google Scholar] [CrossRef]
- Ma, J.; Sheng, H.; Li, X.; Wang, L. iTRAQ-based proteomic analysis reveals the mechanisms of silicon-mediated cadmium tolerance in rice (Oryza sativa) cells. Plant Physiol. Biochem. 2016, 104, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, J.; He, C.; Li, X.; Zhang, W.; Xu, F.; Lin, Y.; Wang, L. Inhibition of cadmium ion uptake in rice (Oryza sativa) cells by a wall-bound form of silicon. New Phytol. 2013, 200, 691–699. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, L.; Liu, J.; Liu, X.; Li, X.; Ma, J.; Lin, Y.; Xu, F. Evidence for silicon within the cell walls of suspension-cultured rice cells. New Phytol. 2013, 200, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Song, A.; Li, P.; Fan, F.; Li, Z.; Liang, Y. The effect of silicon on photosynthesis and expression of its relevant genes in rice (Oryza sativa L.) under high-zinc stress. PLoS ONE 2014, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.H.; Qiu, H.; Tian, T.; Zhan, S.S.; Deng, T.H.B.; Chaney, R.L.; Wang, S.Z.; Tang, Y.T.; Morel, J.L.; Qiu, R.L. Mitigation effects of silicon rich amendments on heavy metal accumulation in rice (Oryza sativa L.) planted on multi-metal contaminated acidic soil. Chemosphere 2011, 83, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Mehrabanjoubani, P.; Abdolzadeh, A.; Sadeghipour, H.R.; Aghdasi, M. Impacts of silicon nutrition on growth and nutrient status of rice plants grown under varying zinc regimes. Theor. Exp. Plant Physiol. 2015, 27, 19–29. [Google Scholar] [CrossRef]
- Wang, S.; Wang, F.; Gao, S. Foliar application with nano-silicon alleviates Cd toxicity in rice seedlings. Environ. Sci. Pollut. Res. 2015, 22, 2837–2845. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Fang, C.; Li, Y.; Lin, W.; He, J.; Lin, R.; Lin, W. Effect of silicon on grain yield of rice under cadmium-stress. Acta Physiol. Plant. 2016, 38, 186–199. [Google Scholar] [CrossRef]
- Fan, X.; Wen, X.; Huang, F.; Cai, Y.; Cai, K. Effects of silicon on morphology, ultrastructure and exudates of rice root under heavy metal stress. Acta Physiol. Plant. 2016, 38, 197–206. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Sonmez, O.; Ince, F.; Higgs, D. Mitigation effects of silicon on maize plants grown at high zinc. J. Plant Nutr. 2009, 32, 1788–1798. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Feng, J.; Shi, Q.; Wang, X.; Wei, M.; Yang, F.; Xu, H. Silicon supplementation ameliorated the inhibition of photosynthesis and nitrate metabolism by cadmium (Cd) toxicity in Cucumissativus L. Sci. Hortic. 2010, 123, 521–530. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophyll a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta (BBA)-Bioenerg. 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Gao, D.; Cai, K.; Chen, J.; Luo, S.; Zeng, R.; Yang, J.; Zhu, X. Silicon enhances photochemical efficiency and adjusts mineral nutrient absorption in Magnaportheoryzea infected rice plants. Acta Physiol. Plant. 2011, 33, 675–682. [Google Scholar] [CrossRef]
- Li, L.; Zhou, W.; Dai, H.; Cao, F.; Zhang, G.; Wu, F. Selenium reduces cadmium uptake and mitigates cadmium toxicity in rice. J. Hazard. Mater. 2012, 343–351. [Google Scholar]
- Hu, H.; Zhang, J.; Wang, H.; Li, R.; Pan, F.; Wu, J.; Feng, Y.; Ying, Y.; Liu, Q. Effect of silicate supplementation on the alleviation of arsenite toxicity in 93-11 (Oryza sativa L. indica). Environ. Sci. Pollut. Res. 2013, 20, 8579–8589. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, D.K.; Singh, V.P.; Prasad, S.M.; Chauhan, D.K.; Dubey, N.K.; Rai, A.K. Silicon-mediated alleviation of Cr(VI) toxicity in wheat seedlings as evidenced by chlorophyll florescence, laser induced breakdown spectroscopy and anatomical changes. Ecotoxicol. Environ. Saf. 2015, 113, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Chalmardi, Z.K.; Abdolzadeh, A.; Sadeghipour, H.R. Silicon nutrition potentiates the antioxidant metabolism of rice plants under iron toxicity. Acta Physiol. Plant. 2014, 36, 493–502. [Google Scholar] [CrossRef]
- Liang, Y.C.; Yang, C.G.; Shi, H.H. Effects of silicon on growth and mineral composition of barley grown under toxic levels of aluminum. J. Plant Nutr. 2001, 24, 229–243. [Google Scholar] [CrossRef]
- Li, P.; Song, A.; Li, Z.; Fan, F.; Liang, Y. Silicon ameliorates manganese toxicity by regulating both physiological processes and expression of genes associated with photosynthesis in rice (Oryza sativa L.). Plant Soil 2015, 397, 289–301. [Google Scholar] [CrossRef]
- Collin, B.; Doelsch, E.; Keller, C.; Panfili, F.; Meunier, J.D. Distribution and variability of silicon, copper and zinc in different bamboo species. Plant Soil 2012, 351, 377–387. [Google Scholar] [CrossRef]
- Lu, H.P.; Zhuang, P.; Li, Z.A.; Tai, Y.P.; Zou, B.; Li, Y.W.; McBride, M.B. Contrasting effects of silicates on cadmium uptake by three dicotyledonous crops grown in contaminated soil. Environ. Sci. Pollut. Res. 2014, 21, 9921–9930. [Google Scholar] [CrossRef] [PubMed]
- Uragauchi, S.; Fujiwara, T. Cadmium transport and tolerance in rice: perspectives for reducing grain cadmium accumulation. Rice 2012, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Neumann, D.; Zur Nieden, U. Silicon and heavy metal tolerance of higher plants. Phytochemistry 2001, 56, 685–692. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, C.; Wang, H.; Zhang, F. Effect of Si on the distribution of Cd in rice seedlings. Plant Soil 2005, 272, 53–60. [Google Scholar] [CrossRef]
- Šimková, L.; Fialová, I.; Vaculíková, M.; Luxová, M. The effect of silicon on the activity and isozymes pattern of antioxidative enzymes of young maize roots under zinc stress. Silicon 2016. [Google Scholar] [CrossRef]
- Bokor, B.; Vaculík, M.; Slováková, Ĺ.; Masarovič, D.; Lux, A. Silicon does not always mitigate zinc toxicity in maize. Acta Physiol. Plant. 2014, 36, 733–743. [Google Scholar] [CrossRef]
- Anwaar, S.A.; Ali, S.; Ali, S.; Ishaque, W.; Farid, M.; Farooq, M.A.; Najeeb, U.; Abbas, F.; Sharif, M. Silicon (Si) alleviates cotton (Gossypiumhirsutum L.) from zinc (Zn) toxicity stress by limiting Zn uptake and oxidative damage. Environ. Sci. Pollut. Res. 2015, 22, 3441–3450. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Cai, Q.; Liu, C.; Wu, L. Silicon alleviates cadmium toxicity in peanut plants in relation to cadmium distribution and stimulation of antioxidative enzymes. Plant Growth Regul. 2010, 61, 45–52. [Google Scholar] [CrossRef]
- Bharwana, S.A.; Ali, S.; Faroop, M.A.; Iqbal, N.; Abbas, F.; Ahmad, M.S.A. Alleviation of lead toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes suppressed lead uptake and oxidative stress in cotton. J. Bioremediat. Biodegrad. 2013, 4, 1–11. [Google Scholar]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doncheva, S.; Poschenrieder, C.; Stoyanova, Z.; Georgieva, K.; Velichkova, M.; Barcelo, J. Silicon amelioration of manganese toxicity in Mn-sensitive and Mn tolerant maize varieties. Environ. Exp. Bot. 2009, 65, 189–197. [Google Scholar] [CrossRef]
- Al-aghabary, K.; Zhu, Z.; Shi, Q. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Physiol. 2004, 27, 2101–2115. [Google Scholar] [CrossRef]
- Hattori, T.; Inanaga, S.; Araki, H.; An, P.; Morita, S.; Luxová, M.; Lux, A. Application of silicon enhanced drought tolerance in sorghum bicolor. Physiol. Plant. 2005, 123, 459–466. [Google Scholar] [CrossRef]
- Song, A.; Li, P.; Li, Z.; Fan, F.; Nikolic, M.; Liang, Y. The alleviation of zinc toxicity by silicon is related to zinc transport and antioxidative reactions in rice. Plant Soil 2011, 344, 319–333. [Google Scholar] [CrossRef]
- Jin, X.; Yang, X.; Islam, E.; Liu, D.; Mahmood, Q. Effects of cadmium on ultrastructure and antioxidative defense system in hyperaccumulator and non-hyperaccumulator ecotypes of Sedum alfredii Hance. J. Hazard. Mater. 2008, 156, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Hasan, K.M.; Ahammed, J.G.; Yin, L.; Shi, K.; Xia, X.; Zhou, Y.; Yu, J.; Zhou, J. Melatonin mitigates cadmium phytotoxicity through modulation of phytochelatins biosynthesis, vacuolar sequestration, and antioxidant potential in Solanum lycopersicum L. Front. Plant Sci. 2015, 6, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, H.; Zhang, Y.; Chai, T. Silicon attenuates cadmium toxicity in Solanumnigrum L. by reducing cadmium uptake an oxidative stress. Plant Physiol. Biochem. 2013, 68, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Maksimović, J.D.; Bogdanović, J.; Maksimović, V.; Nikolic, M. Silicon modulates the metabolism and utilization of phenolic compounds in cucumber (Cucumissativus L.) grown at excess manganese. J. Plant Nutr. Soil Sci. 2007, 170, 739–744. [Google Scholar]
- Shen, X.; Xiao, X.; Dong, Z.; Chen, Y. Silicon effects on antioxidative enzymes and lipid peroxidation in leaves and roots of peanut under aluminum stress. Acta Physiol. Plant. 2014, 36, 3063–3069. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Pandey, P.; Rajpoot, R.; Rani, A.; Gautam, A.; Dubey, R.S. Exogenous application of calcium and silica alleviates cadmium toxicity by suppressing oxidative damage in rice seedlings. Protoplasma 2015, 252, 959–975. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Treatments | Tillering Stage | Ripening Stage | |
|---|---|---|---|---|
| Shoot (g pot−1) | Root (g pot−1) | Shoot (g pot−1) | ||
| Feng-Hua-Zhan | CK | 47 ± 2.3b | 6 ± 0.2a | 83 ± 4.1ab |
| Cd | 42 ± 0.4c | 5 ± 0.4b | 57 ± 3.7d | |
| Zn | 40 ± 0.8c | 4 ± 0.4b | 60 ± 0.7d | |
| Cd + Zn | 48 ± 0.9b | 6 ± 0.2b | 56 ± 2.8d | |
| Cd + Si | 54 ± 2.2b | 7 ± 0.3a | 73 ± 3.1c | |
| Zn + Si | 51 ± 2.1ab | 7 ± 0.6a | 77 ± 2.7bc | |
| Cd + Zn + Si | 54 ± 1.5a | 7 ± 0.6a | 91 ± 3.0a | |
| Hua-Hang-Si-Miao | CK | 47 ± 3.6ab | 3 ± 0.2cd | 145 ± 7.4ab |
| Cd | 41 ± 0.4b | 3 ± 0.5cde | 78 ± 5.3d | |
| Zn | 38 ± 1.9b | 2 ± 0.1e | 82 ± 5.8d | |
| Cd + Zn | 45 ± 2.6ab | 2 ± 0.2de | 94 ± 8.7d | |
| Cd + Si | 45 ± 0.1ab | 4 ± 0.1c | 116 ± 4.4c | |
| Zn + Si | 43 ± 1.8ab | 5 ± 0.6b | 132 ± 8.5bc | |
| Cd + Zn + Si | 51 ± 4.7a | 7 ± 0.5a | 158 ± 5.2a | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, F.; Wen, X.-H.; Cai, Y.-X.; Cai, K.-Z. Silicon-Mediated Enhancement of Heavy Metal Tolerance in Rice at Different Growth Stages. Int. J. Environ. Res. Public Health 2018, 15, 2193. https://doi.org/10.3390/ijerph15102193
Huang F, Wen X-H, Cai Y-X, Cai K-Z. Silicon-Mediated Enhancement of Heavy Metal Tolerance in Rice at Different Growth Stages. International Journal of Environmental Research and Public Health. 2018; 15(10):2193. https://doi.org/10.3390/ijerph15102193
Chicago/Turabian StyleHuang, Fei, Xiao-Hui Wen, Yi-Xia Cai, and Kun-Zheng Cai. 2018. "Silicon-Mediated Enhancement of Heavy Metal Tolerance in Rice at Different Growth Stages" International Journal of Environmental Research and Public Health 15, no. 10: 2193. https://doi.org/10.3390/ijerph15102193
APA StyleHuang, F., Wen, X. -H., Cai, Y. -X., & Cai, K. -Z. (2018). Silicon-Mediated Enhancement of Heavy Metal Tolerance in Rice at Different Growth Stages. International Journal of Environmental Research and Public Health, 15(10), 2193. https://doi.org/10.3390/ijerph15102193