Ascorbate-Glutathione Cycle and Ultrastructural Analyses of Two Kenaf Cultivars (Hibiscus cannabinus L.) under Chromium Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Measurement of Chromium Content
2.3. Bio-Concentration Factor (BCF), Translocation Factor (TF) and Tolerance Index (TI)
2.4. Measurement of Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2) Contents
2.5. Chloroplast Ultrastructural Observation
2.6. Measurement of Enzymatic Activities
2.7. Measurement of Ascorbate and Glutathione Contents
2.8. Statistical Analysis
3. Results
3.1. Screening Cr-Tolerant and Cr-Sensitive Kenaf Cultivars
3.2. Plant Biomass and Cr Accumulation
3.3. Effect of Chromium on MDA and H2O2 Contents
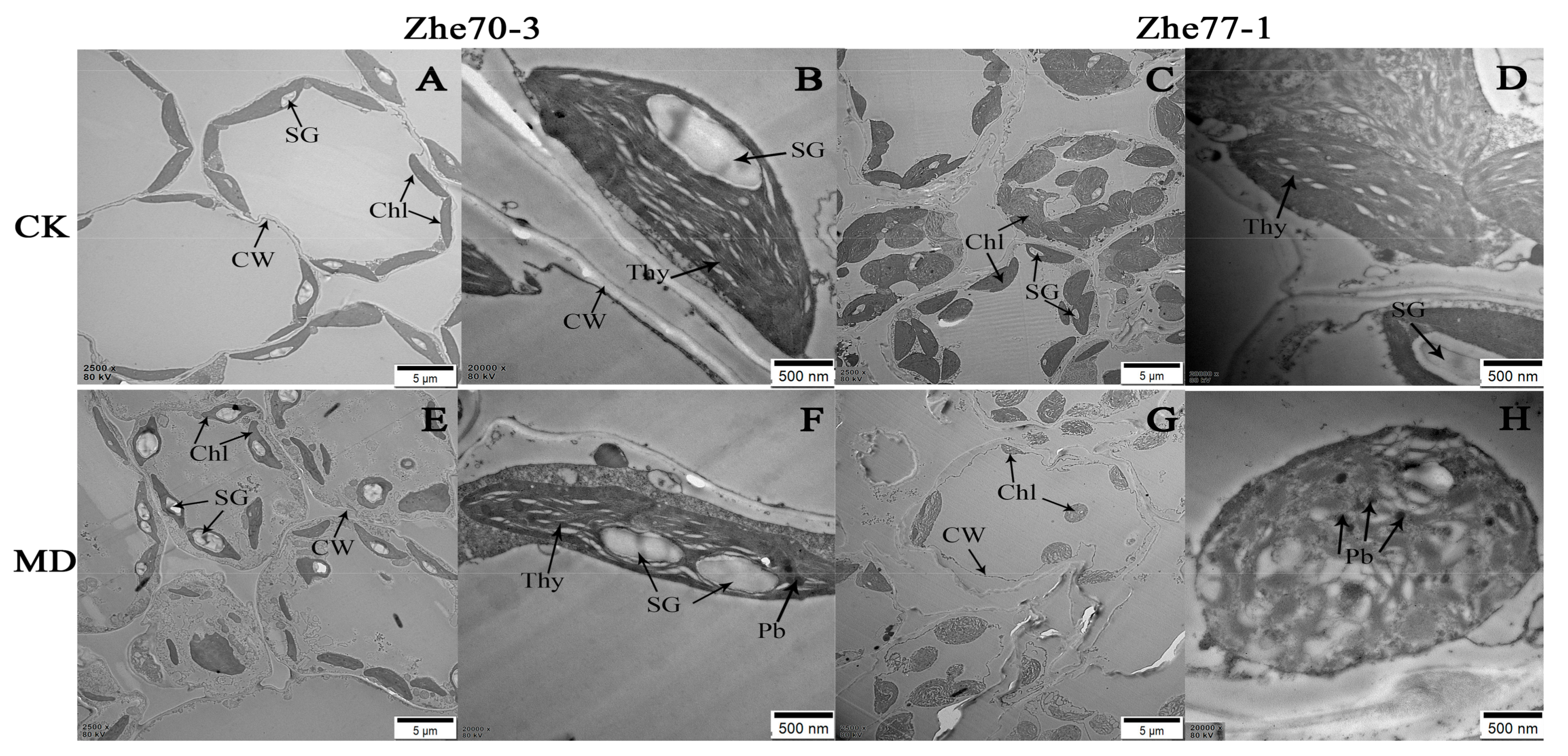
3.4. Effect of Chromium on Chloroplast Ultrastructure
3.5. Effect of Chromium on Enzyme Activities of AsA-GSH Cycle
3.6. Effect of Chromium on Ascorbate and Glutathione Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hawley, E.L.; Deeb, R.A.; Kavanaugh, M.C.; Jacobs, J.A. Treatment Technologies for Chromium (VI). Cheminform 2010, 37. [Google Scholar] [CrossRef]
- Schiavon, M.; Pilonsmits, E.A.; Wirtz, M.; Hell, R.; Malagoli, M. Interactions between chromium and sulfur metabolism in Brassica juncea. J. Environ. Qual. 2008, 37, 1536–1545. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.; Santos, C.; Azevedo, R.; Moutinhopereira, J.; Correia, C.; Dias, M.C. Chromium (VI) induces toxicity at different photosynthetic levels in pea. Plant Physiol. Biochem. 2012, 53, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, A.; Bibi, I.; Niazi, N.K.; Ok, Y.S.; Murtaza, G.; Shahid, M.; Kunhikrishnan, A.; Li, D.; Mahmood, T. Chromium (VI) sorption efficiency of acid-activated banana peel over organo-montmorillonite in aqueous solutions. Int. J. Phytoremediat. 2016, 19, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Becquer, T.; Quantin, C.; Sicot, M.; Boudot, J. Chromium availability in ultramafic soils from New Caledonia. Sci. Total Environ. 2003, 301, 251–261. [Google Scholar] [CrossRef]
- Prado, F.E.; Hilal, M.; Chocobar-Ponce, S.; Pagano, E.; Rosa, M.; Prado, C. Chapter 6—Chromium and the Plant: A Dangerous Affair? Elsevier Inc: New York, NY, USA, 2016. [Google Scholar]
- Azmat, R.; Khanum, R. Effect of Chromium Metal on the Uptakes of Mineral Atoms in Seedlings of Bean Plant Vigna radiata (L.) Wilckzek. Pak. J. Boil. Sci. 2005, 8, 281–283. [Google Scholar] [CrossRef]
- Sharma, P.; Kumar, A.; Bhardwaj, R. Plant steroidal hormone epibrassinolide regulate—Heavy metal stress tolerance in Oryza sativa L. by modulating antioxidant defense expression. Environ. Exp. Bot. 2016, 122, 1–9. [Google Scholar] [CrossRef]
- Chaney, R.L.; Malik, M.; Li, Y.M.; Brown, S.L.; Brewer, E.P.; Angle, J.S.; Baker, A.J. Phytoremediation of soil metal. Curr. Opin. Biotechnol. 1997, 8, 279–284. [Google Scholar] [CrossRef]
- Zhao, F.J.; McGrath, S.P. Biofortification and phytoremediation. Curr. Opin. Plant Biol. 2009, 12, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Meints, P.D.; Smith, C.A. Kenaf seed storage duration on germination, emergence, and yield. Ind. Crops Prod. 2003, 17, 9–14. [Google Scholar] [CrossRef]
- Catroga, A.; Fernando, A.; Oliveira, J.S. Effects on growth, productivity and biomass quality of kenaf of soils contaminated with heavy metals. Eur. Biomass Conf. 2005, 14, 149–152. [Google Scholar]
- Ashori, A.; Harun, J.; Raverty, W.; Yusoff, M. Chemical and morphological characteristics of Malaysian cultivated kenaf (Hibiscus cannabinus) fiber. Polym. Plast. Technol. Eng. 2006, 45, 131–134. [Google Scholar] [CrossRef]
- Saba, N.; Paridah, M.T.; Jawaid, M. Mechanical properties of kenaf fibre reinforced polymer composite: A review. Constr. Build. Mater. 2015, 76, 87–96. [Google Scholar] [CrossRef]
- Patnaik, A.R.; Achary, V.M.M.; Panda, B.B. Chromium (VI)-induced hormesis and genotoxicity are mediated through oxidative stress in root cells of Allium cepa L. Plant Growth Regul. 2013, 71, 157–170. [Google Scholar] [CrossRef]
- Islam, M.K.; Khanam, M.S.; Siyoung, L.; Alam, I.; Mooryong, H. The interaction of arsenic (As) and chromium (Cr) influences growth and antioxidant status in tossa jute (Corchorus olitorius). Plant Omics 2014, 7, 499–509. [Google Scholar]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Riaz, M.; Shahzad, S.M.; Imran, Q.; Ali, I. Combined ability of chromium (Cr) tolerant plant growth promoting bacteria (PGPB) and salicylic acid (SA) in attenuation of chromium stress in maize plants. Plant Physiol. Biochem. 2016, 108, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Cıftcıyiılmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.Y.; Du, J.; Yuan, Y.H.; Shu, S.; Sun, J.; Guo, S.R. Effects of 24-epibrassinolide on ascorbate-glutathione cycle and polyamine levels in cucumber roots under Ca(NO3)2 stress. Acta Physiol. Plant 2013, 35, 253–262. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquezgarcia, B.; Queval, G.; Foyer, C.H. Glutathione in plants: An integrated overview. Plant Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef] [PubMed]
- Qiu, B.; Zeng, F.; Cai, S.; Wu, X.; Haider, S.I.; Wu, F.; Zhang, G. Alleviation of chromium toxicity in rice seedlings by applying exogenous glutathione. J. Plant Physiol. 2013, 170, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Nahakpam, S.; Shah, K. Expression of key antioxidant enzymes under combined effect of heat and cadmium toxicity in growing rice seedlings. Plant Growth Regul. 2011, 63, 23–35. [Google Scholar] [CrossRef]
- Scoccianti, V.; Iacobucci, M.; Paoletti, M.F.; Fraternale, A.; Speranza, A. Species-dependent chromium accumulation, lipid peroxidation, and glutathione levels in germinating kiwifruit pollen under Cr(III) and Cr(VI) stress. Chemosphere 2008, 73, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.K.; Jena, P.P.; Kundu, S. Antioxidative efficiency of Triticum aestivum L. exposed to chromium stress. J. Environ. Biol. 2009, 30, 539–544. [Google Scholar] [PubMed]
- Ajuwon, O.R.; Bada, B.S.; Olujimi, O.O. Growth and antioxidative responses to excess cadmium in Kenaf (Hibiscus cannabinus L.). Fresenius Environ. Bull. 2010, 19, 2637–2643. [Google Scholar]
- Cao, F.; Wang, N.; Zhang, M.; Dai, H.; Dawood, M.; Zhang, G.; Wu, F. Comparative study of alleviating effects of GSH, Se and Zn under combined contamination of cadmium and chromium in rice (Oryza sativa). BioMetals 2013, 26, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Wang, G.; Lou, L.; Lv, J. Physiological responses and tolerance of kenaf (Hibiscus cannabinus L.) exposed to chromium. Ecotoxicol. Environ. Saf. 2016, 133, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 357–359. [Google Scholar]
- Li, C. Physiological and biochemical responses to high Mn concentrations in two contrasting Populus cathayana populations. Chemosphere 2007, 68, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Cai, Q. Cadmium tolerance and accumulation in eight potential energy crops. Biotechnol. Adv. 2009, 27, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef] [Green Version]
- Logan, B.A.; Grace, S.C.; Adams, W.W.; Demmig-Adams, B. Seasonal Differences in Xanthophyll Cycle Characteristics and Antioxidants in Mahonia repens Growing in Different Light Environments. Oecologia 1998, 116, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pinto, M.C.D.; Tommasi, F.; Gara, L.D. Enzymes of the ascorbate biosynthesis and ascorbate-glutathione cycle in cultured cells of tobacco Bright Yellow 2. Plant Physiol. Biochem. 2000, 38, 541–550. [Google Scholar] [CrossRef]
- Esaka, M.; Hattori, T.; Fujisawa, K.; Sakajo, S.; Asahi, T. Molecular cloning and nucleotide sequence of full-length cDNA for ascorbate oxidase from cultured pumpkin cells. FEBS J. 1990, 191, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,5′-dithiobis(2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef]
- Zhang, J.X.; Kirkham, M.B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallick, S.; Sinam, G.; Kumar, M.R.; Sinha, S. Interactive effects of Cr and Fe treatments on plants growth, nutrition and oxidative status in Zea mays L. Ecotoxicol. Environ. Saf. 2010, 73, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Qing, X.; Zhao, X.; Hu, C.; Wang, P.; Zhang, Y.; Zhang, X.; Wang, P.; Shi, H.; Jia, F.; Qu, C. Selenium alleviates chromium toxicity by preventing oxidative stress in cabbage (Brassica campestris L. ssp. Pekinensis) leaves. Ecotoxicol. Environ. Saf. 2015, 114, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Ghazy, U.M.M. Modifications of evaluation index and subordinate function formulae to determine superiority of mulberry silkworm crosses. J. Basic Appl. Zool. 2014, 67, 1–9. [Google Scholar] [CrossRef]
- Bishehkolaei, R.; Fahimi, H.; Saadatmand, S.; Nejadsattari, T. Ultrastructural localisation of chromium in Ocimum basilicum. Turk. J. Bot. 2014, 35, 261–268. [Google Scholar] [CrossRef]
- Ali, S.; Chaudhary, A.; Rizwan, M.; Anwar, H.T.; Adrees, M.; Farid, M.; Irshad, M.K.; Hayat, T.; Anjum, S.A. Alleviation of chromium toxicity by glycinebetaine is related to elevated antioxidant enzymes and suppressed chromium uptake and oxidative stress in wheat (Triticum aestivum L.). Environ. Sci. Pollut. Res. Int. 2015, 22, 10669–10678. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.; Panda, S.K. Toxic Effects, Oxidative Stress and Ultrastructural Changes in Moss Taxithelium nepalense (Schwaegr.) Broth. Under Chromium and Lead Phytotoxicity. Water Air Soil Pollut. 2005, 167, 73–90. [Google Scholar] [CrossRef]
- Gill, R.A.; Zang, L.; Ali, B.; Farooq, M.A.; Cui, P.; Yang, S.; Ali, S.; Zhou, W. Chromium-induced physio-chemical and ultrastructural changes in four cultivars of Brassica napus L. Chemosphere 2015, 120, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Seth, C.S.; Chaturvedi, P.K.; Misra, V. The role of phytochelatins and antioxidants in tolerance to Cd accumulation in Brassica juncea L. Ecotoxicol. Environ. Saf. 2008, 71, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Wahid, A.; Ahmad, S.S.; Butt, Z.A.; Tariq, M. Ecophysiological responses of rice (Oryza sativa L.) to hexavalent chromium. Pak. J. Bot. 2011, 43, 2853–2859. [Google Scholar]
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Chromium toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Liu, Q.; Yao, H. Antioxidant activities of barley seeds extracts. Food Chem. 2007, 102, 732–737. [Google Scholar] [CrossRef]
- Haroldsen, V.M.; ChiHam, C.L.; Kulkarni, S.; Lorence, A.; Bennett, A.B. Constitutively expressed DHAR and MDHAR influence fruit, but not foliar ascorbate levels in tomato. Plant Physiol. Biochem. 2011, 49, 1244–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Bai, P.; Zeng, F.; Cai, S.; Shamsi, I.H.; Qiu, B.; Wu, F.; Zhang, G. The ecotoxicological and interactive effects of chromium and aluminum on growth, oxidative damage and antioxidant enzymes on two barley genotypes differing in Al tolerance. Environ. Exp. Bot. 2011, 70, 185–191. [Google Scholar] [CrossRef]
- Cai, Y.; Cao, F.; Cheng, W.; Zhang, G.; Wu, F. Modulation of exogenous glutathione in phytochelatins and photosynthetic performance against Cd stress in the two rice genotypes differing in Cd tolerance. Biol. Trace Element Res. 2011, 143, 1159–1173. [Google Scholar] [CrossRef] [PubMed]
- Seregin, I.V.; Ivanov, V.B. Physiological Aspects of Cadmium and Lead Toxic Effects on Higher Plants. Russ. J. Plant Physiol. 2001, 48, 523–544. [Google Scholar] [CrossRef]
- Toppi, L.S.D.; Fossati, F.; Musetti, R.; Mikerezi, I.; Favali, M.A. Effects of hexavalent chromium on maize, tomato, and cauliflower plants. J. Plant. Nutr. 2002, 25, 701–717. [Google Scholar] [CrossRef]
- Gomez, L.D.; Noctor, G.; Knight, M.R.; Foyer, C.H. Regulation of calcium signalling and gene expression by glutathione. J. Exp. Bot. 2004, 55, 1851–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, V.; Vajpayee, P.; Singh, S.N.; Mehrotra, S. Effect of chromium accumulation on photosynthetic pigments, oxidative stress defense system, nitrate reduction, proline level and eugenol content of Ocimum tenuiflorum L. Plant Sci. 2004, 167, 1159–1169. [Google Scholar] [CrossRef]
- Martí, M.C.; Camejo, D.; Olmos, E.; Sandalio, L.M.; Fernández-García, N.; Jiménez, A.; Sevilla, F. Characterisation and changes in the antioxidant system of chloroplasts and chromoplasts isolated from green and mature pepper fruits. Plant Boil. 2009, 11, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.Y.; Hong, C.Y.; Chinghuei, K.; Tullio, M.C.D.; Stasolla, C. The decline in ascorbic acid content is associated with cadmium toxicity of rice seedlings. Plant Physiol. Biochem. 2010, 48, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Hojati, M.; Modarres-Sanavy, S.A.; Enferadi, S.T.; Majdi, M.; Ghanati, F.; Farzadfar, S.; Pazoki, A. Cadmium and copper induced changes in growth, oxidative metabolism and terpenoids of Tanacetum parthenium. Environ. Sci. Pollut. Res. Int. 2017, 24, 12261–12272. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Root Length | Shoot Height | Shoot DW | Root DW | Mean | Rank |
|---|---|---|---|---|---|---|
| Zhe70-3 | 1.154 | 1.000 | 0.337 | 0.310 | 0.700 | 1 |
| Yueyin83-23 | 0.423 | 0.203 | 1.000 | 1.000 | 0.657 | 2 |
| Hongyin135 | 1.000 | 0.432 | 0.408 | 0.103 | 0.486 | 3 |
| Zhe54-3 | 0.808 | 0.619 | 0.000 | 0.379 | 0.452 | 4 |
| Fuhong992 | 0.500 | 0.751 | 0.245 | 0.241 | 0.434 | 5 |
| Kransdoy | 0.000 | 0.482 | 0.735 | 0.414 | 0.408 | 6 |
| VG | 0.269 | 0.482 | 0.622 | 0.241 | 0.404 | 7 |
| T19 | 0.539 | 0.787 | 0.276 | 0.000 | 0.400 | 8 |
| Zhe77-1 | 0.115 | 0.000 | 0.133 | 0.241 | 0.122 | 9 |
| Cultivars | Treatment Cr (mM) | Growth Characteristic | Cr Content (mg kg−1 DW) | |||||
|---|---|---|---|---|---|---|---|---|
| Shoot Height (cm) | Root Length (cm) | Fresh Weight (g) | Dry Weight (g) | Shoot | Root | |||
| Zhe70-3 | CK | 21.500 ± 1.637e | 17.133 ± 1.270d | 0.928 ± 0.080e | 0.067 ± 0.005c | ND | ND | |
| 0.5 | 19.733 ± 0.473d | 15.933 ± 0.404d | 0.710 ± 0.101d | 0.061 ± 0.008c | 420 ± 0.453a | 7897 ± 0.766a | ||
| 1 | 16.633 ± 0.208c | 13.167 ± 2.003c | 0.563 ± 0.067bc | 0.052 ± 0.002b | 3344 ± 1.260c | 10,467 ± 0.467b | ||
| 1.5 | 15.167 ± 0.321b | 12.900 ± 0.700c | 0.503 ± 0.078ab | 0.045 ± 0.002a | 4874 ± 0.897d | 11,233 ± 0.645c | ||
| Zhe77-1 | CK | 19.133 ± 0.451d | 13.567 ± 0.808c | 0.847 ± 0.046e | 0.057 ± 0.002c | ND | ND | |
| 0.5 | 17.000 ± 0.346c | 11.033 ± 0.551b | 0.648 ± 0.081cd | 0.050 ± 0.003b | 678 ± 0.178ab | 6134 ± 0.269a | ||
| 1 | 16.033 ± 0.416bc | 8.467 ± 1.159a | 0.486 ± 0.038ab | 0.045 ± 0.005a | 2889 ± 2.865c | 10,345 ± 0.563b | ||
| 1.5 | 12.700 ± 0.656a | 7.700 ± 0.458a | 0.433 ± 0.033a | 0.040 ± 0.002a | 4156 ± 0.278d | 11,789 ± 1.678c | ||
| Cultivars | Treatment | MDA (mmol·g−1 FW) | H2O2 (µmol·g−1 FW) | ||
|---|---|---|---|---|---|
| Cr (mM) | Shoot | Root | Shoot | Root | |
| Zhe70-3 | CK | 0.623 ± 0.271a | 0.578 ± 0.042a | 0.912 ± 0.213a | 0.879 ± 0.092a |
| 0.5 | 1.542 ± 0.330ab | 0.843 ± 0.359ab | 1.348 ± 0.379ab | 1.154 ± 0.313ab | |
| 1 | 2.377 ± 0.219b | 1.470 ± 0.650ab | 2.791 ± 0.147b | 2.532 ± 0.131bc | |
| 1.5 | 2.775 ± 0.204b | 2.656 ± 0.174c | 3.917 ± 0.219c | 3.214 ± 0.326c | |
| Zhe77-1 | CK | 0.869 ± 0.102a | 0.746 ± 0.233a | 1.386 ± 0.132a | 0.945 ± 0.147a |
| 0.5 | 1.781 ± 0.121ab | 1.418 ± 0.347ab | 1.731 ± 0.426ab | 1.246 ± 0.314ab | |
| 1 | 2.496 ± 0.253b | 2.356 ± 0.195c | 2.324 ± 0.202b | 1.958 ± 0.420b | |
| 1.5 | 3.464 ± 0.146c | 2.853 ± 0.169c | 3.905 ± 0.324c | 3.567 ± 0.589c | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, L.; Cao, R.; Kang, J.; Zhang, X.; Lv, J. Ascorbate-Glutathione Cycle and Ultrastructural Analyses of Two Kenaf Cultivars (Hibiscus cannabinus L.) under Chromium Stress. Int. J. Environ. Res. Public Health 2018, 15, 1467. https://doi.org/10.3390/ijerph15071467
Niu L, Cao R, Kang J, Zhang X, Lv J. Ascorbate-Glutathione Cycle and Ultrastructural Analyses of Two Kenaf Cultivars (Hibiscus cannabinus L.) under Chromium Stress. International Journal of Environmental Research and Public Health. 2018; 15(7):1467. https://doi.org/10.3390/ijerph15071467
Chicago/Turabian StyleNiu, Lianmei, Rang Cao, Jingquan Kang, Xu Zhang, and Jinyin Lv. 2018. "Ascorbate-Glutathione Cycle and Ultrastructural Analyses of Two Kenaf Cultivars (Hibiscus cannabinus L.) under Chromium Stress" International Journal of Environmental Research and Public Health 15, no. 7: 1467. https://doi.org/10.3390/ijerph15071467
APA StyleNiu, L., Cao, R., Kang, J., Zhang, X., & Lv, J. (2018). Ascorbate-Glutathione Cycle and Ultrastructural Analyses of Two Kenaf Cultivars (Hibiscus cannabinus L.) under Chromium Stress. International Journal of Environmental Research and Public Health, 15(7), 1467. https://doi.org/10.3390/ijerph15071467