Country-Level Relationships of the Human Intake of N and P, Animal and Vegetable Food, and Alcoholic Beverages with Cancer and Life Expectancy

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection and Preparation
2.2. Statistical Analyses
Bayesian Models
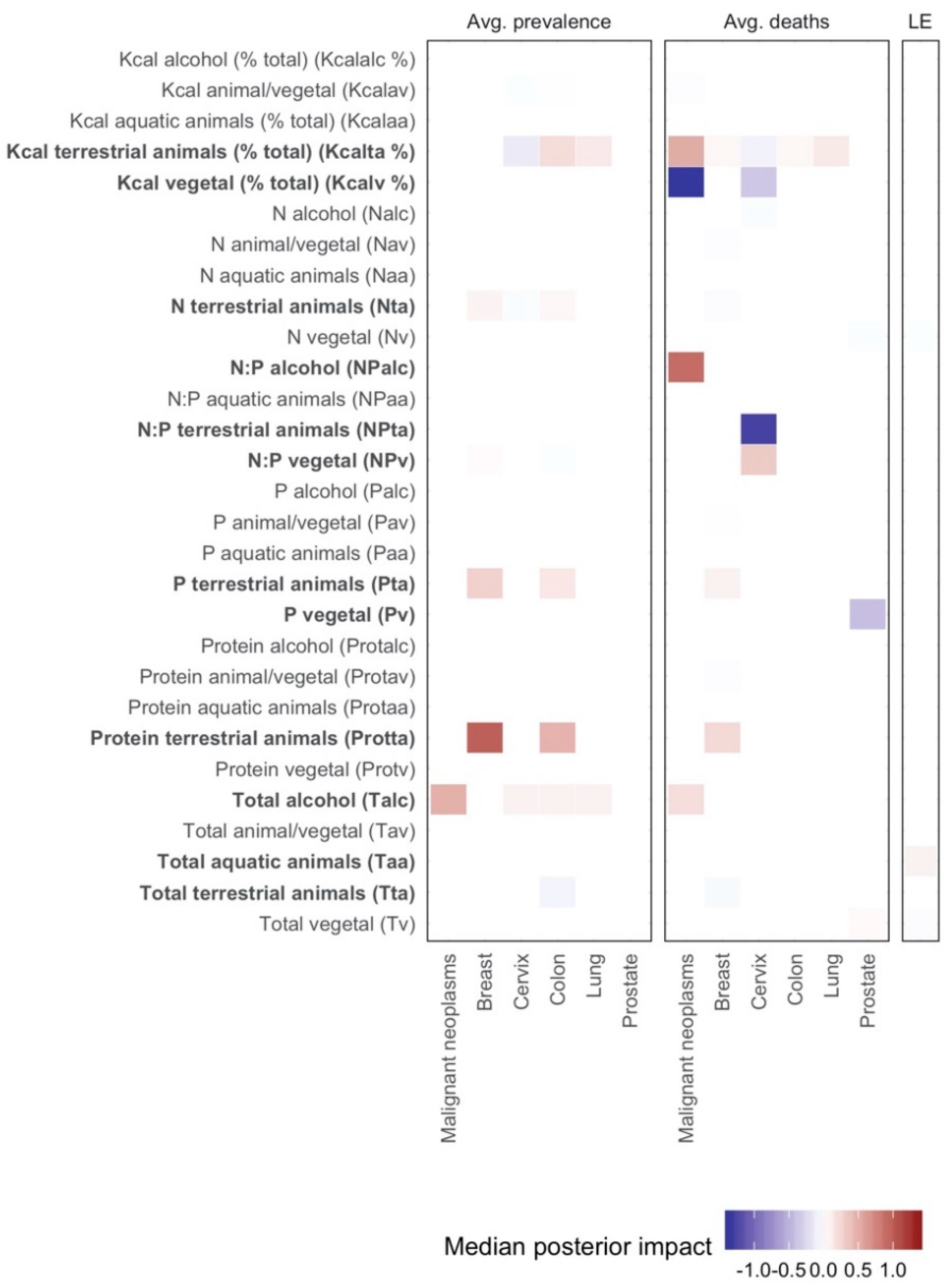
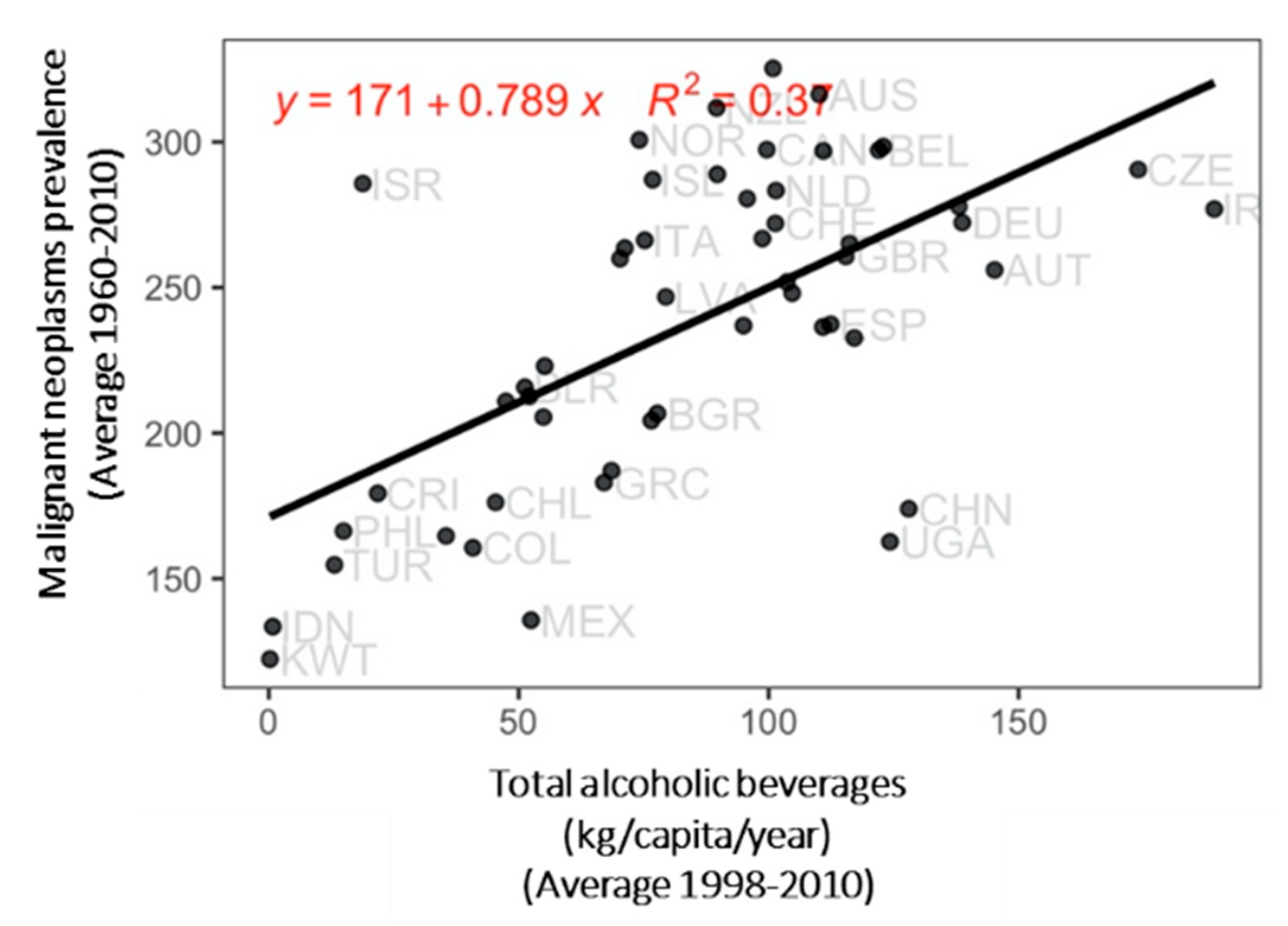
- In the first analysis, we attempted to explain the average prevalence of total malignant neoplasms, as well as breast, cervix, colon, lung, and prostate cancers using our set of averaged indicators in the period 1998–2010 ( 52).
- In the second analysis, we attempted to explain the average deaths (per 100,000 population) during the period 1960–2010 due to malignant neoplasms, as well as breast, cervix, colon, lung, and prostate cancers using our set of averaged covariates in the period 1960–2010 ( 85).
- Finally, we regressed the average increase in life expectancy from 1960 to 2010 on our set of covariates measured in their averages over the period 1960–2010 (100). We also regressed the increase in life expectancy on the change of our set of covariates during this period.
2.3. Reduced Major-Axis Analyses, Generalized Linear Models, and Principal Component Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Peñuelas, J.; Janssens, I.A.; Ciais, P.; Obersteiner, M.; Krisztin, T.; Piao, S.; Sardans, J. Increasing gap in human height between rich and poor countries associated to their different intakes of N and P. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, A.C.; Caballero, B.H.; Cousins, R.J.; Tucker, K.L.; Ziegler, T.R. Modern Nutrition in Health and Disease; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2014. [Google Scholar]
- Wada, K.; Oba, S.; Tsuji, M.; Tamura, T.; Konishi, K.; Goto, Y.; Mizuta, F.; Koda, S.; Hori, A.; Tanabashi, S.; et al. Meat consumption and colorectal cancer risk in Japan: The Takayama study. Cancer Sci. 2017, 108, 1065–1070. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.M.; Mucci, L.A.; Drake, B.F.; Preston, M.A.; Stampfer, M.J.; Giovannucci, E.; Kibel, A.S. Meat, Fish, Poultry, and Egg Intake at Diagnosis and Risk of Prostate Cancer Progression. Cancer Prev. Res. 2016, 9, 933–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.H.; Dai, Q.; Xiang, Y.B.; Zhao, G.M.; Zheng, W.; Gao, Y.T.; Ruan, Z.X.; Cheng, J.R.; Shu, X.O. Animal food intake and cooking methods in relation to endometrial cancer risk in Shanghai. Br. J. Cancer 2006, 95, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Dwan, C.; Miles, A. The role of attitude and attitude ambivalence in acceptance of the cancer risk associated with red meat. Health Risk Soc. 2018, 20, 147–162. [Google Scholar] [CrossRef]
- Hur, S.J.; Jo, C.; Yoon, Y.; Jeong, J.Y.; Lee, K.T. Controversy on the correlation of red and processed meat consumption with colorectal cancer risk: An Asian perspective. Crit. Rev. Food Sci. Nutr. 2019, 59, 3526–3537. [Google Scholar] [CrossRef]
- Zheng, W.; Kushi, L.; Potter, J.; Sellers, T.; Doyle, T.; Bostick, R.; Folsom, A. Dietary intake of energy and animal foods and endometrial cancer incidence. The Iowa women’s health study. Am. J. Epidemiol. 1995, 142, 388–394. [Google Scholar] [CrossRef]
- IARC. 2020. Available online: https://monographs.iarc.fr/agents-classified-by-the-iarc/ (accessed on 20 March 2020).
- IARC, American Institute for Cancer Research. Health Eat. Am. Inst. Cancer Res. (2020), WRC. 2020. Available online: https://www.aicr.org/cancer-prevention/healthy-eating/ (accessed on 12 April 2020).
- Crippa, A.; Larsson, S.C.; Discacciati, A.; Wolk, A.; Orsini, N. Red and processed meat consumption and risk of bladder cancer: A dose–response meta-analysis of epidemiological studies. Eur. J. Nutr. 2018, 57, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Perloy, A.; Maasland, D.H.E.; van den Brandt, P.A.; Kremer, B.; Schouten, L.J. Intake of meat and fish and risk of head–neck cancer subtypes in the Netherlands Cohort Study. Cancer Causes Control 2017, 28, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Le Marchand, L.; Wilkens, L.R.; Hankin, J.H.; Kolonel, L.N.; Lyu, L.C. A case-control study of diet and colerectal cancer in a multiethnic population in Hawaii (United States): Lipids and foods of animal origin. Cancer Causes Control 1997, 8, 637–648. [Google Scholar] [CrossRef]
- Joshi, A.D.; Corral, R.; Catsburg, C.; Lewinger, J.P.; Koo, J.; John, E.M.; Ingles, S.A.; Stern, M.C. Red meat and poultry, cooking practices, genetic susceptibility and risk of prostate cancer: Results from a multiethnic case-control study. Carcinogenesis 2012, 33, 2108–2118. [Google Scholar] [CrossRef] [Green Version]
- Bao, P.P.; Shu, X.O.; Zheng, Y.; Cai, H.; Ruan, Z.X.; Gu, K.; Su, Y.; Gao, Y.T.; Zheng, W.; Lu, W. Fruit, vegetable, and animal food intake and breast cancer risk by hormone receptor status. Nutr. Cancer 2012, 64, 806–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diallo, A.; Deschasaux, M.; Latino-Martel, P.; Hercberg, S.; Galan, P.; Fassier, P.; Allès, B.; Guéraud, F.; Pierre, F.H.; Touvier, M. Red and processed meat intake and cancer risk: Results from the prospective NutriNet-Santé cohort study. Int. J. Cancer 2018, 142, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Van Hecke, T.; Vossen, E.; Hemeryck, L.Y.; Vanden Bussche, J.; Vanhaecke, L.; De Smet, S. Increased oxidative and nitrosative reactions during digestion could contribute to the association between well-done red meat consumption and colorectal cancer. Food Chem. 2015, 187, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Samraj, A.N.; Pearce, O.M.T.; Läubli, H.; Crittenden, A.N.; Bergfeld, A.K.; Band, K.; Gregg, C.J.; Bingman, A.E.; Secrest, P.; Diaz, S.L.; et al. A red meat-derived glycan promotes inflammation and cancer progression. Proc. Natl. Acad. Sci. USA 2015, 112, 542–547. [Google Scholar] [CrossRef] [Green Version]
- Schulz, M.; Nöthlings, U.; Allen, N.; Onland-Moret, N.C.; Agnoli, C.; Engeset, D.; Galasso, R.; Wirfält, E.; Tjønneland, A.; Olsen, A.; et al. No association of consumption of animal foods with risk of ovarian cancer. Cancer Epidemiol. Biomark. Prev. 2007, 16, 852–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcondes, L.H.; Franco, O.H.; Ruite, R.; Arfan Ikram, M.; Mulder, M.; Stricker, B.H.; Kiefte-de Jon, J.C. Animal foods and postmenopausal breast cancer risk: A prospective cohort study. Br. J. Nutr. 2019, 122, 586–591. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.E.; Key, T.J.; Appleby, P.N.; Travis, R.C.; Roddam, A.W.; Tjønneland, A.; Johnsen, N.F.; Overvad, K.; Linseisen, J.; Rohrmann, S.; et al. Animal foods, protein, calcium and prostate cancer risk: The European prospective investigation into cancer and nutrition. Br. J. Cancer 2008, 98, 1574–1581. [Google Scholar] [CrossRef] [Green Version]
- Do, M.; Lee, S.; Jung, P.; Lee, M. Intake of fruits, vegetables, and soy foods in relation to breast cancer risk in Korean women: A case-control study. Nutr. Cancer 2007, 57, 20–27. [Google Scholar] [CrossRef]
- Ferreira, P.M.P.; Rodrigues, L.A.R.L.; de Alencar Carnib, L.P.; de Lima Sousa, P.V.; Nolasco Lugo, L.M.; Nunes, N.M.F.; do Nascimento Silva, J.; da Silva Araûjo, L.; de Macêdo Gonçalves Frota, K. Cruciferous Vegetables as Antioxidative, Chemopreventive and Antineoplasic Functional Foods: Preclinical and Clinical Evidences of Sulforaphane Against Prostate Cancers. Curr. Pharm. Des. 2019, 24, 4779–4793. [Google Scholar] [CrossRef]
- Freudenheim, J.L.; Ritz, J.; Smith-Warner, S.A.; Albanes, D.; Bandera, E.V.; Van Den Brandt, P.A.; Colditz, G.; Feskanich, D.; Goldbohm, R.A.; Harnack, L.; et al. Alcohol consumption and risk of lung cancer: A pooled analysis of cohort studies. Am. J. Clin. Nutr. 2005, 82, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, A.; Claggett, B.; Jhund, P.S.; Rosamond, W.; Deswal, A.; Aguilar, D.; Shah, A.M.; Cheng, S.; Solomon, S.D. Alcohol consumption and risk of heart failure: The Atherosclerosis Risk in Communities Study. Eur. Heart J. 2015, 36, 939–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Chen, Y.M.; Huang, J.; Fang, Y.J.; Huang, W.Q.; Yan, B.; Lu, M.S.; Pan, Z.Z.; Zhang, C.X. Flavonoid intake from vegetables and fruits is inversely associated with colorectal cancer risk: A case-control study in China. Br. J. Nutr. 2016, 116, 1275–1287. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Trudo, S.P.; Gallaher, D.D. Apiaceous and Cruciferous Vegetables Fed during the Post-Initiation Stage Reduce Colon Cancer Risk Markers in Rats. J. Nutr. 2019, 149, 249–257. [Google Scholar] [CrossRef]
- IARC. Biological agents. A review of human carcinogenesis. In Monographs on the Evolution of Carcinogenic Risk to Humans; World Health Organization, Ed.; International Agency for Research on Cancer: Lyon, Paris, 2010; Volume 100, 487p. [Google Scholar]
- ARC. Alcohol consumption and ethyl carbamate. In Monographs on the Evolution of Carcinogenic Risk to Humans; World Health Organization, Ed.; International Agency for Research on Cancer: Lyon, Paris, 2010; Volume 96, 1428p. [Google Scholar]
- Bagnardi, V.; Blangiardo, M.; La Vecchia, C.; Corrao, G. A meta-analysis of alcohol drinking and cancer risk. Br. J. Cancer 2001, 85, 1700–1705. [Google Scholar] [CrossRef] [PubMed]
- Pelucchi, C.; Gallus, S.; Garavello, W.; Bosetti, C.; La Vecchia, C. Cancer risk associated with alcohol and tobacco use: Focus on upper aero-digestive tract and liver. Alcohol Res. Health 2006, 29, 193–198. [Google Scholar]
- Handa, A.; Fatima, T.; Mattoo, A. Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front. Chem. Chem. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Prudhomme, M. Global nitrogen fertilizer supply and demand outlook. Sci. China Ser. C Life Sci. 2005, 48, 818–826. [Google Scholar] [CrossRef]
- Gupta, U.C.; Gupta, S.C. Sources and Deficiency Diseases of Mineral Nutrients in Human Health and Nutrition: A Review. Pedosphere 2014, 24, 13–38. [Google Scholar] [CrossRef]
- Kelley, L.A.; Gardner, S.P.; Sutcliffe, M.J. An automated approach for clustering an ensemble of NMR-derived protein structures into conformationally related subfamilies. Protein Eng. 1996, 9, 1063–1065. [Google Scholar] [CrossRef] [Green Version]
- Altieri, A.; Garavello, W.; Bosetti, C.; Gallus, S.; La Vecchia, C. Alcohol consumption and risk of laryngeal cancer. Oral Oncol. 2005, 41, 956–965. [Google Scholar] [CrossRef] [PubMed]
- Webb, P.M.; Purdie, D.M.; Bain, C.J.; Green, A.C. Alcohol, wine, and risk of epithelial ovarian cancer. Cancer Epidemiol. Biomark. Prev. 2004, 13, 592–599. [Google Scholar]
- Elser, J.J.; Acharya, K.; Kyle, M.; Cotner, J.; Makino, W.; Markow, T.; Watts, T.; Hobbie, S.; Fagan, W.; Schade, J.; et al. Growth rate-stoichiometry couplings in diverse biota. Ecol. Lett. 2003, 6, 936–943. [Google Scholar] [CrossRef] [Green Version]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princenton, NJ, USA, 2002. [Google Scholar]
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R.; Folarin, A.; Huberty, A.; Interlandi, S.; Kilham, S.S.; McCauley, E.; Schulz, K.L.; et al. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578–580. [Google Scholar] [CrossRef]
- Elser, J.J.; Sterner, R.W.; Galford, A.E.; Chrzanowski, T.H.; Findlay, D.L.; Mills, K.H.; Paterson, M.J.; Stainton, M.P.; Schindler, D.W. Pelagic C:N:P stoichiometry in a eutrophied lake: Responses to a whole-lake food-web manipulation. Ecosystems 2000, 3, 293–307. [Google Scholar] [CrossRef]
- Hessen, D.O.; Jensen, T.C.; Kyle, M.; Elser, J.J. RNA responses to N- and P-limitation; reciprocal regulation of stoichiometry and growth rate in Brachionus. Funct. Ecol. 2007, 21, 956–962. [Google Scholar] [CrossRef]
- Elser, J.J.; Kyle, M.M.; Smith, M.S.; Nagy, J.D. Biological stoichiometry in human cancer. PLoS ONE 2007, 2, e1028. [Google Scholar] [CrossRef] [Green Version]
- Peñuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen–phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef] [Green Version]
- Everitt, A.V.; Le Couteur, D.G. Life extension by calorie restriction in humans. Ann. N. Y. Acad. Sci. 2007, 1114, 428–433. [Google Scholar] [CrossRef]
- Willcox, D.C.; Willcox, B.J.; Todoriki, H.; Curb, J.D.; Suzuki, M. Caloric restriction and human longevity: What can we learn from the Okinawans? Biogerontology 2006, 7, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Shanley, D.P.; Kirkwood, T.B.L. Caloric restriction does not enhance longevity in all species and is unlikely to do so in humans. Biogerontology 2006, 7, 165–168. [Google Scholar] [CrossRef] [PubMed]
- Anjana; Umar, S.; Iqbal, M. Nitrate accumulation in plants, factors affecting the process, and human health implications. A review. Agron. Sustain. Dev. 2007, 27, 45–57. [Google Scholar] [CrossRef]
- Segi, M. Cancer Mortality for Selected Sites in 24 Countries (1950–57); Dept. of Public Health, Tohoku University School of Medicine: Sendai, Japan, 1960. [Google Scholar]
- Cooke, A. Dietary Food-Additive Phosphate and Human Health Outcomes. Compr. Rev. Food Sci. Food Saf. 2017, 16, 906–1021. [Google Scholar] [CrossRef] [Green Version]
- Trautvetter, U.; Jahreis, G.; Kiehntopf, M.; Glei, M. Consequences of a high phosphorus intake on mineral metabolism and bone remodeling in dependence of calcium intake in healthy subjects—A randomized placebo-controlled human intervention study. Nutr. J. 2016, 15, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Seitz, H.K.; Becker, P. Alcohol Metabolism and Cancer Risk. Alcohol Res. Health 2007, 30, 38–47. [Google Scholar] [PubMed]
- George, E.I.; McCulloch, R.E. Variable selection via Gibbs sampling. J. Am. Stat. Assoc. 1993, 88, 881–889. [Google Scholar] [CrossRef]
- Turati, F.; Rossi, M.; Pelucchi, C.; Levi, F.; La Vecchia, C. Fruit and vegetables and cancer risk: A review of southern European studies. Br. J. Nutr. 2015, 113, S102–S110. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 2014, 3, 1–117. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penuelas, J.; Krisztin, T.; Obersteiner, M.; Huber, F.; Winner, H.; Janssens, I.A.; Ciais, P.; Sardans, J. Country-Level Relationships of the Human Intake of N and P, Animal and Vegetable Food, and Alcoholic Beverages with Cancer and Life Expectancy. Int. J. Environ. Res. Public Health 2020, 17, 7240. https://doi.org/10.3390/ijerph17197240
Penuelas J, Krisztin T, Obersteiner M, Huber F, Winner H, Janssens IA, Ciais P, Sardans J. Country-Level Relationships of the Human Intake of N and P, Animal and Vegetable Food, and Alcoholic Beverages with Cancer and Life Expectancy. International Journal of Environmental Research and Public Health. 2020; 17(19):7240. https://doi.org/10.3390/ijerph17197240
Chicago/Turabian StylePenuelas, Josep, Tamás Krisztin, Michael Obersteiner, Florian Huber, Hannes Winner, Ivan A. Janssens, Philippe Ciais, and Jordi Sardans. 2020. "Country-Level Relationships of the Human Intake of N and P, Animal and Vegetable Food, and Alcoholic Beverages with Cancer and Life Expectancy" International Journal of Environmental Research and Public Health 17, no. 19: 7240. https://doi.org/10.3390/ijerph17197240
APA StylePenuelas, J., Krisztin, T., Obersteiner, M., Huber, F., Winner, H., Janssens, I. A., Ciais, P., & Sardans, J. (2020). Country-Level Relationships of the Human Intake of N and P, Animal and Vegetable Food, and Alcoholic Beverages with Cancer and Life Expectancy. International Journal of Environmental Research and Public Health, 17(19), 7240. https://doi.org/10.3390/ijerph17197240