Evaluation of Polish Wild Mushrooms as Beta-Glucan Sources



Abstract
:1. Introduction
2. Materials and Methods
2.1. Studied Materials
2.2. Sample Preparation Procedure
2.3. Glucan Determination
2.3.1. The Megazyme Method
2.3.2. The Congo Red Method
2.4. Statistical Analysis
3. Results and Discussion
3.1. Evaluation of the Total Glucan, Alpha-Glucan and Beta-Glucan Content Using Megazyme Assay Kit
3.2. Evaluation of Beta-Glucans Using Triple Stage Extraction and the Congo Red Method
3.3. Extraction and Quantitative Determination Methods of β-Glucans
- -
- Extraction with water at different temperatures;
- -
- Extraction in aqueous solutions of different pH;
- -
- Extraction under special physical conditions, e.g., using ultrasound, microwave, radiation or increased pressure [50].
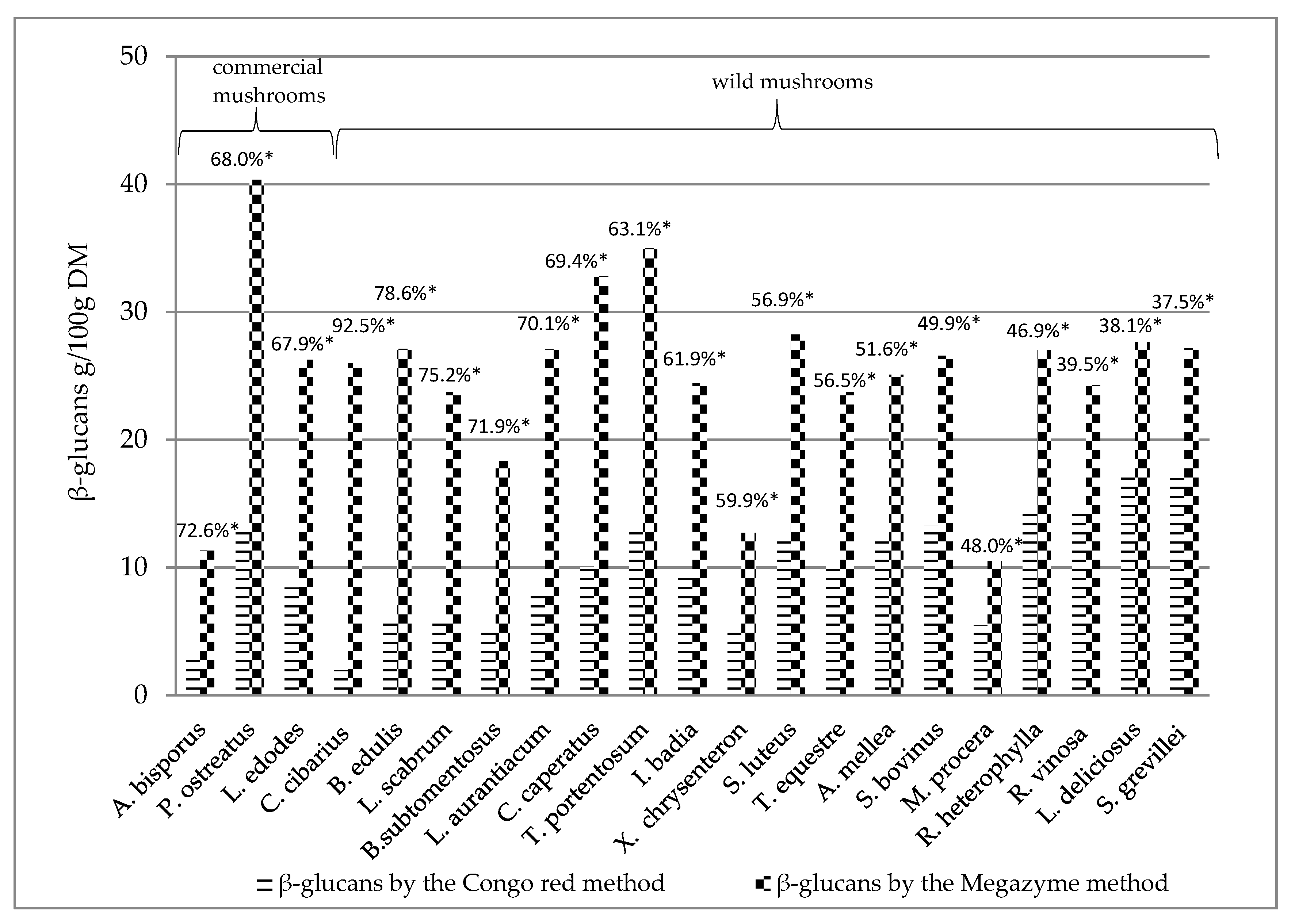
3.4. Evaluation of β-Glucans in Wild and Commercial Cultivated Mushrooms
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gow, N.A.R.; Latge, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 3. [Google Scholar]
- Liu, X.; Wang, R.; Bi, J.; Kang, L.; Zhou, J.; Duan, B.; Liu, Z.; Yuan, S. A novel endo-β-1,6-glucanase from the mushroom Coprinopsis cinerea and its application in studying of cross-linking of β-1,6-glucan and the wall extensibility in stipe cell walls. Int. J. Biol. Macromol. 2020, 160, 612–622. [Google Scholar] [PubMed]
- Baeva, E.; Bleha, R.; Lavrova, E.; Sushytskyi, L.; Čopíková, J.; Jablonsky, I.; Klouček, P.; Synytsya, A. Polysaccharides from basidiocarps of cultivating mushroom Pleurotus ostreatus: Isolation and structural characterization. Molecules 2019, 24, 2740. [Google Scholar]
- Morales, D.; Rutckeviski, R.; Villalva, M.; Abreu, H.; Soler-Rivas, C.; Santoyo, S.; Iacomini, M.; Smiderle, F.R. Isolation and comparison of α- and β-d-glucans from shiitake mushrooms (Lentinula edodes) with different biological activities. Carbohydr. Polym. 2020, 229, 115521. [Google Scholar]
- Samuelsen, A.B.C.; Rise, F.; Wilkins, A.L.; Teveleva, L.; Nyman, A.A.T.; Aachmann, F.L. The edible mushroom Albatrellus ovinus contains a α-l-fuco-α-d-galactan, α-d-glucan, a branched (1 → 6)-β-d-glucan and a branched (1 → 3)-β-d-glucan. Carbohydr. Res. 2019, 471, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Vetvickova, J. Anti-infectious and anti-tumor activities of β-glucans. Anticancer Res. 2020, 40, 3139–3145. [Google Scholar] [PubMed]
- De Felice, B.; Damiano, S.; Montanino, C.; Del Buono, A.; La Rosa, G.; Guida, B.; Santillo, M. Effect of beta- and alpha-glucans on immune modulating factors expression in enterocyte-like Caco-2 and goblet-like LS 174T cells. Int. J. Biol. Macromol. 2020, 153, 600–607. [Google Scholar]
- Ruthes, A.C.; Smiderle, F.R.; Iacomini, M. d-glucans from edible mushrooms: A review on the extraction, purification and chemical characterization approaches. Carbohydr. Polym. 2015, 117, 753–761. [Google Scholar]
- Semedo, M.C.; Karmali, A.; Fonseca, L. A high throughput colorimetric assay of β-1,3-d-glucans by Congo red dye. J. Microbiol. Methods 2015, 109, 140–148. [Google Scholar]
- Meng, Y.; Lyu, F.; Xu, X.; Zhang, L. Recent advances in chain conformation and bioactivities of triple-helix polysaccharides. Biomacromolecules 2020, 21, 1653–1677. [Google Scholar] [PubMed]
- Ruthes, A.C.; Smiderle, F.R.; Iacomini, M. Mushroom heteropolysaccharides: A review on their sources, structure and biological effects. Carbohydr. Polym. 2016, 136, 358–375. [Google Scholar] [PubMed]
- Ruiz-Herrera, J.; Ortiz-Castellanos, L. Cell wall glucans of fungi. A review. Cell. Surf. 2019, 5, 100022. [Google Scholar] [PubMed]
- Yoshimi, A.; Miyazawa, K.; Abe, K. Function and biosynthesis of cell wall α-1,3-glucan in fungi. J. Fungi 2017, 3, 63. [Google Scholar]
- McCleary, B.V.; Draga, A. Measurement of β-glucan in mushrooms and mycelial products. J. AOAC Int. 2016, 99, 364–373. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Cui, S.W.; Cheung, P.C.K.; Wang, Q. Antitumor polysaccharides from mushrooms: A review on their isolation process, structural characteristic and antitumor activity. Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhou, W.; Zhang, Y. Fungal polysaccharides. Adv. Pharmacol. 2020, 87, 277–299. [Google Scholar]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar]
- Laurino, L.F.; Viroel, F.J.M.; Caetano, E.; Spim, S.; Pickler, T.B.; Rosa-Castro, R.M.; Vasconcelos, E.A.; Jozala, A.F.; Hataka, A.; Grotto, D.; et al. Lentinus edodes exposure before and after fetus implantation: Materno-fetal development in rats with gestational diabetes mellitus. Nutrients 2019, 11, 2720. [Google Scholar]
- Roupas, P.; Keogh, J.; Noakes, M.; Margetts, C.; Taylor, P. The role of edible mushrooms in health. J. Funct. Foods 2012, 4, 687–709. [Google Scholar] [CrossRef]
- Morales, D.; Tejedor-Calvo, E.; Jurado-Chivato, N.; Polo, G.; Tabernero, M.; Ruiz-Rodríguez, A.; Largo, C.; Soler-Rivasa, C. In vitro and in vivo testing of the hypocholesterolemic activity of ergosterol- and β-glucan-enriched extracts obtained from shiitake mushrooms (Lentinula edodes). Food Funct. 2019, 10, 7325–7332. [Google Scholar] [PubMed]
- Ina, K.; Kataoka, T.; Ando, T. The use of lentinan for treating gastric cancer. Anticancer Agents Med. Chem. 2013, 13, 681–688. [Google Scholar] [PubMed] [Green Version]
- Du, B.; Meenu, M.; Liu, H.; Xu, B. A concise review on the molecular structure and function relationship of β-Glucan. Int. J. Mol. Sci. 2019, 20, 4032. [Google Scholar]
- Wang, Q.; Sheng, X.; Shi, A.; Hu, H.; Yang, Y.; Liu, L.; Fei, L.; Liu, H. β-Glucans: Relationships between modification, conformation and functional activities. Molecules 2017, 22, 257. [Google Scholar]
- Nakashima, A.; Yamada, K.; Iwata, O.; Sugimoto, R.; Atsuji, K.; Ogawa, T.; Ishibashi-Ohgo, N.; Suzuki, K. β-Glucan in foods and its physiological functions. J. Nutr. Sci. Vitaminol. Tokyo 2018, 64, 8–17. [Google Scholar]
- Maheshwari, G.; Sowrirajan, S.; Joseph, B. β-Glucan, a dietary fiber in effective prevention of lifestyle diseasesAn insight. Bioact. Carbohydr. Diet. Fibre 2019, 19, 100187. [Google Scholar]
- Wu, J.Y. Polysaccharide-protein complexes from edible fungi and applications. In Polysaccharides: Bioactivity and Biotechnology; Ramawat, K.G., Merillon, J.M., Eds.; Springer International Publishing: New York, NY, USA, 2015; pp. 927–937. [Google Scholar] [CrossRef]
- Seo, G.; Hyun, C.; Choi, S.; Kim, Y.M.; Cho, M. The wound healing effect of four types of beta-glucan. Appl. Biol. Chem. 2019, 62, 20. [Google Scholar]
- Rathore, H.; Prasad, S.; Sharma, S. Mushroom nutraceuticals for improved nutrition and better human health: A review. Pharm. Nutr. 2017, 5, 35–46. [Google Scholar]
- Nitschke, J.; Modick, H.; Busch, E.; von Rekowski, R.W.; Altenbach, H.J.; Mölleken, H. A new colorimetric method to quantify β-1,3-1,6-glucans in comparison with total β-1,3-glucans in edible mushrooms. Food Chem. 2011, 127, 791–796. [Google Scholar] [PubMed]
- Sari, M.; Prange, A.; Lelley, J.I.; Hambitzer, R. Screening of beta-glucan contents in commercially cultivated and wild growing mushrooms. Food Chem. 2017, 216, 45–51. [Google Scholar]
- Synytsya, A.; Míčková, K.; Jablonský, I.; Siuková, M.; Čopiková, J. Mushrooms of genus Pleurotus as a source of dietary fibres and glucans for food supplements. Czech J. Food Sci. 2008, 26, 441–446. [Google Scholar]
- Kanagasabapathy, G.; Kuppusamy, U.R.; Abd Malek, S.N.; Abdulla, M.A.; Chua, K.H.; Sabaratnam, V. Glucan-rich polysaccharides from Pleurotus sajor-caju (Fr.) Singer prevents glucose intolerance, insulin resistance and inflammation in C57BL/6J mice fed a high-fat diet. BMC Complement. Altern. Med. 2012, 12, 261. [Google Scholar]
- Park, H.; Ka, K.H.; Ryu, S.R. Enhancement of β-glucan content in the cultivation of cauliflower mushroom (Sparassis latifolia) by elicitation. Mycobiology 2014, 42, 41–45. [Google Scholar]
- Ogawa, K.; Tsurugi, J.; Watanabe, T. The dependence of the conformation of a (1→3)-β-d-glucan on chain-length in alkaline solution. Carbohydr. Res. 1973, 29, 397–403. [Google Scholar]
- Mao, C.F.; Hsu, M.C.; Hwang, W.H. Physicochemical characterization of grifolan: Thixotropic properties and complex formation with Congo Red. Carbohydr. Polym. 2007, 68, 502–510. [Google Scholar] [CrossRef]
- Mirończuk-Chodakowska, I.; Socha, K.; Zujko, M.E.; Terlikowska, K.M.; Borawska, M.H.; Witkowska, A.M. Copper, manganese, selenium and zinc in wild-growing edible mushrooms from the eastern territory of “Green Lungs of Poland”: Nutritional and toxicological implications. Int. J. Environ. Res. Public Health 2019, 16, 3614. [Google Scholar]
- Fluck, M. Atlas of Mushrooms. Identification, Collection, Using; Publishing House Delta W-Z: Warsaw, Poland, 2012. [Google Scholar]
- Mushrooms Key. Available online: https://www.grzyby.pl/pelna/atlas-grzyby-przyrodnika.htm (accessed on 12 September 2019).
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E.; Terlikowska, K.M. Quantitative evaluation of 1,3-1,6-β-d-glucan contents in wild-growing species of edible Polish mushrooms. Rocz. Panstw. Zakl. Hig. 2017, 68, 281–290. [Google Scholar] [PubMed]
- Kalač, P. Chemical composition and nutritional value of European species of wild growing mushrooms. A review. Food Chem. 2009, 113, 9–16. [Google Scholar]
- Ouzouni, P.K.; Petridis, D.; Koller, W.D.; Riganakos, K.A. Nutritional value and metal content of wild edible mushrooms collected from West Macedonia and Epirus, Greece. Food Chem. 2009, 115, 1575–1580. [Google Scholar] [CrossRef]
- Árvay, J.; Tomáš, J.; Hauptvogl, M.; Kopernická, M.; Kováčik, A.; Bajčan, D.; Massányi, P. Contamination of wild-grown edible mushrooms by heavy metals in a former mercury-mining area. J. Environ. Sci. Health B 2014, 49, 815–827. [Google Scholar] [PubMed]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonsky, I.; Spěváčekc, J.; Erband, V.; Kováříková, E.; Čopíková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar]
- Avni, S.; Ezove, N.; Hanani, H.; Yadid, I.; Karpovsky, M.; Hayby, H.; Gover, O.; Hadar, Y.; Schwartz, B.; Danay, O. Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii. Int. J. Mol. Sci. 2017, 18, 1564. [Google Scholar]
- Smiderle, F.R.; Olsen, L.M.; Carbonero, E.R.; Beggio, C.H.; Freitas, C.S.; Marcon, R.; Santos, A.R.S.; Gorin, P.A.J.; Iacomini, M. Anti-inflammatory and analgesic properties in a rodent model of a (1-3),(1-6)-linked beta-glucan isolated from Pleurotus pulmonarius. Eur. J. Pharmacol. 2008, 597, 86–91. [Google Scholar]
- de Jesus, L.I.; Smiderle, F.R.; Cordeiro, L.M.C.; de Freitas, R.A.; van Griensven, L.J.L.D.; Iacomini, M. Simple and effective purification approach to dissociate mixed water-insoluble α- and β-d-glucans and its application on the medicinal mushroom Fomitopsis betulina. Carbohydr. Polym. 2018, 200, 353–360. [Google Scholar]
- Yan, J.K.; Wang, W.Q.; Wu, J.Y. Recent advances in Cordyceps sinensis polysaccharides: Mycelial fermentation, isolation, structure and bioactivities: A review. J. Funct. Foods. 2014, 6, 33–47. [Google Scholar]
- Bhanja, S.K.; Rout, D.; Patra, P.; Sen, I.K.; Nandan, C.K.; Islam, S.S. Water-insoluble glucans from the edible fungus Ramaria botrytis. Bioact. Carbohydr. Diet. Fibre 2014, 3, 52–58. [Google Scholar] [CrossRef]
- Dong, Q.; Wang, Y.; Shi, L.; Yao, J.; Li, J.; Ma, F.; Ding, K. A novel water-soluble β-d-glucan isolated from the spores of Ganoderma lucidum. Carbohydr. Res. 2012, 353, 100–105. [Google Scholar]
- Gil-Ramírez, A.; Smiderle, F.R.; Morales, D.; Iacomini, M.; Soler-Rivas, C. Strengths and weaknesses of the aniline-blue method used to test mushroom (1→3)-β-d-glucans obtained by microwave-assisted extractions. Carbohydr. Polym. 2019, 217, 135–143. [Google Scholar] [PubMed] [Green Version]
- Mizuno, M.; Minato, K.; Kawakami, S.; Tatsuoka, S.; Denpo, Y.; Tsuchida, H. Contents of anti-tumor polysaccharides in certain mushrooms and their immunomodulating activities. Food Sci. Technol. Res. 2001, 7, 31–34. [Google Scholar]
- Ko, Y.T.; Lin, Y.L. 1,3-beta-glucan quantification by a fluorescence microassay and analysis of its distribution in foods. J. Agric. Food Chem. 2004, 52, 3313–3318. [Google Scholar]
- Cha, Y.J.; Alam, N.; Lee, J.S.; Lee, K.R.; Shim, M.J.; Lee, M.W.; Kim, H.Y.; Shin, P.G.; Cheong, J.C.; Yoo, Y.B.; et al. Anticancer and immunopotentiating activities of crude polysaccharides from Pleurotus nebrodensis on mouse sarcoma 180. Mycobiology 2012, 40, 236–243. [Google Scholar] [PubMed] [Green Version]
- Kang, S.C.; Koo, H.J.; Park, S.; Lim, J.D.; Kim, Y.J.; Kim, T.; Namkoong, S.; Jang, K.H.; Pyo, S.; Jang, S.A.; et al. Effects of β-glucans from Coriolus versicolor on macrophage phagocytosis are related to the Akt and CK2/Ikaros. Int. J. Biol. Macromol. 2013, 57, 9–16. [Google Scholar] [PubMed]
- Gründemann, C.; Garcia-Käufer, M.; Sauer, B.; Scheer, R.; Merdivan, S.; Bettin, P.; Huber, R.; Lindequist, U. Comparative chemical and biological investigations of β-glucan-containing products from shiitake mushrooms. J. Funct. Foods. 2015, 18, 692–702. [Google Scholar]
- Toledo, R.C.C.; Carvalho, M.A.; Lima, L.C.O.; de Barros Vilas-Boas, E.V.; Dias, E.S. Measurement of β-glucan and other nutritional characteristic in disttict strains of Agaricus subrufescens mushrooms. Afic. J. Biotechnol. 2013, 12, 6203–6209. [Google Scholar] [CrossRef]
- Kono, H.; Kondo, N.; Hirabayashi, K.; Ogata, M.; Totani, K.; Ikematsu, S.; Osada, M. NMR spectroscopic structural characterization of a water-soluble β-(1→ 3, 1→ 6)-glucan from Aureobasidium pullulans. Carbohydr. Polym. 2017, 174, 876–886. [Google Scholar]
- Fu, Y.; Lin, S.; Lu, M.; Wei, S.Y.; Zhou, J.; Zhao, L.; Zhang, Q.; Lin, D.R.; Liu, Y.T.; Chen, H.; et al. Quantitative evaluation of ultrasound-assisted extraction of 1,3-β-glucans from Dictyophora indusiata using an improved fluorometric assay. Polymers 2019, 11, 864. [Google Scholar]
- McCleary, B.V.; Holmes, M.G. Enzymic quantification of (1-3), (1-4) β-glucan in barley and malt. J. Inst. Brew. 1985, 91, 285–295. [Google Scholar]
- Manzi, P.; Pizzoferrato, L. Beta glucans in edible mushrooms. Food Chem. 2000, 68, 315–318. [Google Scholar]
- Bak, W.C.; Park, J.H.; Park, Y.A.; Ka, K.H. Determination of glucan contents in the fruiting bodies and mycelia of Lentinula edodes cultivars. Mycobiology 2014, 42, 301–304. [Google Scholar]
- Danielson, M.D.; Dauth, R.; Elmasry, N.A.; Langeslay, R.R.; Magee, A.S.; Will, P.M. Enzymatic method to measure β-1,3-β-1,6-glucan content in extracts and formulated products (GEM assay). J. Agric. Chem. 2010, 58, 10305–10308. [Google Scholar] [CrossRef]
- Manzi, P.; Aguzzi, A.; Pizzoferrato, L. Nutritional value of mushrooms widely consumed in Italy. Food Chem. 2001, 73, 321–325. [Google Scholar] [CrossRef]
- Rhee, S.J.; Cho, S.Y.; Kim, K.M.; Cha, D.S.; Park, H.J. A comparative study of analytical methods for alkali-soluble b-glucan in medicinal mushroom, Chaga (Inonotus obliquus). Food Sci. Technol. 2008, 41, 545–549. [Google Scholar]
- Zhu, F.; Du, B.; Bian, Z.; Xu, B. Beta-glucans from edible and medicinal mushrooms: Characteristics, physicochemical and biological activities. J. Food Compos. Anal. 2015, 41, 165–173. [Google Scholar]
- Guo, L.Q.; Li, J.Y.; Lin, J.F. Non-volatile components of several novel species of edible fungi in China. Food Chem. 2007, 100, 643–649. [Google Scholar]
- Boonyanuphap, J.; Hansawasdi, C. Spatial distribution of Beta glucan containing wild mushroom communities in subtropical dry forest, Thailand. Fung. Divers. 2011, 46, 29–42. [Google Scholar]
- Chen, J.; Seviour, R. Medical importance of fungal β-(1-3), (1-6)-glucans. Mycol. Res. 2007, 111, 635–652. [Google Scholar] [CrossRef]
- Nile, S.H.; Park, S.W. Total, soluble, and insoluble dietary fibre contents of wild growing edible mushrooms. Czech J. Food Sci. 2014, 32, 302–307. [Google Scholar]
- Zhang, Y.; Kong, H.; Fang, Y.; Nishinari, K.; Phillips, G.O. Schizophyllan: A review on its structure, properties, bioactivities and recent developments. Bioact. Carbohydr. Diet. Fibre 2013, 1, 53–71. [Google Scholar]
- Khan, A.A.; Gani, A.; Khanday, F.A.; Masoodi, F.A. Biological and pharmaceutical activities of mushroom β-glucan discussed as a potential functional food ingredient. Bioact. Carbohydr. Diet. Fibre 2018, 16, 1–13. [Google Scholar] [CrossRef]
- Ruthes, A.C.; Carbonero, E.R.; Córodva, M.M.; Baggio, C.H.; Santos, A.R.S.; Sassaki, G.L.; Cipriani, T.R.; Gorin, P.A.J.; Iacomini, M. Lactarius rufus (1-3),(1-6)-β-d-glucans: Structure, antinociceptive and anti-inflammatory effects. Carbohydr. Polym. 2013, 94, 129–136. [Google Scholar]
- Hou, Y.; Ding, X.; Hou, W.; Song, B.; Wang, T.; Wang, F.; Zhong, J. Immunostimulant activity of a novel polysaccharide isolated from Lactarius deliciosus (L. ex Fr.). Gray. Indian J. Pharm Sci. 2013, 75, 393–399. [Google Scholar]
- Gil-Ramírez, A.; Clavijo, C.; Palanisamy, M.; Soler, R.C.; Ruiz-Rodriguez, A.; Marin, F.R.; Reglero, G.; Pėrez, M. Edible mushrooms as potential sources of new hypocholesterolemic compounds. In Mushroom Biology and Mushroom Products, Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Products, Arcachon, France, 4–7 October 2011; Institut National de la Recherche Agronomique (INRA): Arcachon, France; Volume 2, pp. 110–119.
- Khatua, S.; Acharya, K. Water soluble antioxidative crude polysaccharide from Russula senecis elicits TLR modulated NF-κB signaling pathway and pro-inflammatory response in murine macrophages. Front. Pharmacol. 2018, 9, 985. [Google Scholar] [CrossRef]
- Falandysz, J. Selenium in edible mushrooms. J. Environ. Sci. Health Part C 2008, 26, 256–299. [Google Scholar]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Indole compounds in some culinary-medicinal higher basidiomycetes from Poland. Int. J. Med. Mush. 2011, 13, 449–454. [Google Scholar]
- Witkowska, A.M.; Zujko, M.E.; Mirończuk-Chodakowska, I. Comparative study of wild edible mushrooms as sources of antioxidants. Int. J. Med. Mush. 2011, 13, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Bovi, M.; Carrizo, M.E.; Capaldi, S.; Perduca, M.; Chiarelli, L.R.; Galliano, M.; Monaco, H.L. Structure of a lectin with antitumoral properties in king bolete (Boletus edulis) mushrooms. Glycobiology 2011, 21, 1000–1009. [Google Scholar]
- Wu, S.; Wang, G.; Ruhi, Y.; Cui, Y. Anti-inflammatory effects of Boletus edulis polysaccharide on asthma pathology. Am. J. Transl. Res. 2016, 8, 4478–4489. [Google Scholar]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food. Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef]
- Zhang, D.; Frankowska, A.; Jarzyńska, G.; Kojta, A.K.; Drewnowska, M.; Wydmańska, D.; Bielawski, L.; Wang, J.; Falandysz, J. Metals of king bolete (Boletus edulis) bull.: Fr. collected at the same site over two years. Afr. J. Agric. Res. 2010, 5, 3050–3055. [Google Scholar]
- Ryu, S.R.; Ka, K.H.; Park, H.; Bak, W.C.; Lee, B.H. Cultivation characteristics of Sparassis crispa strains using sawdust medium of Larix kaempferi. Korea J. Med. Mycol. 2009, 37, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T. Natural Products and Biological Activity of the Pharmacologically Active Cauliflower Mushroom Sparassis crispa. Biomed. Res. Int. 2013, 2013, 982317. [Google Scholar]
- Chaichian, S.; Moazzami, B.; Sadoughi, F.; Kashani, H.H.; Zaroudi, M.; Asemi, Z. Functional activities of beta-glucans in the prevention or treatment of cervical cancer. J. Ovarian Res. 2020, 13, 24. [Google Scholar]
- Abraham, A.; Ostroff, G.; Levitz, S.M.; Oyston, P.C.F. A novel vaccine platform using glucan particles for induction of protective responses against Francisella tularensis and other pathogens. Clin. Exp. Immunol. 2019, 198, 143–152. [Google Scholar]
- Peltzer, M.A.; Delgado, J.F.; Salvay, A.G.; Wagner, J.R. β-Glucan, a promising polysaccharide for bio-based films developments for food contact materials and medical application. Curr. Org. Chem. 2017, 21, 1–6. [Google Scholar] [CrossRef]
- Murphy, E.J.; Masterson, C.; Rezoagli, E.; O’Toole, D.; Major, I.; Stack, G.D.; Lynch, M.; Laffey, J.G.; Rowan, N.J. β-Glucan extracts from the same edible shiitake mushroom Lentinus edodes produce differential in-vitro immunomodulatory and pulmonary cytoprotective effects—Implications for coronavirus disease (COVID-19) immunotherapies. Sci. Total Environ. 2020, 732, 139330. [Google Scholar]



{kind=link}
{kind=link}
| No. | Mushroom Species | N | Moisture % | Total Glucans * | Alpha–Glucans ** | Beta-Glucans *** | |||
|---|---|---|---|---|---|---|---|---|---|
| X ± SD | Range | X ± SD | Range | X ± SD | Range | ||||
| g/100 g DM | |||||||||
| wild edible mushrooms | |||||||||
| 1. | Armillaria mellea (Vahl) P. Kumm. | 3 | 86.57–91.76 | 26.2 ± 2.92 | 23.1–28.9 | 1.11 ± 0.08 | 1.05–1.20 | 25.09 ± 2.85 | 22.05–27.70 |
| 2. | Imleria badia (Fr.) Fr. | 4 | 89.23–93.55 | 27.7 ± 0.38 | 27.4–28.1 | 3.24 ± 1.38 | 1.82–4.58 | 24.43 ± 1.05 | 23.52–25.58 |
| 3. | Boletus edulis Bull. | 3 | 90.72–92.04 | 31.0 ± 0.87 | 30.0–31.5 | 3.90 ± 1.87 | 2.20–5.90 | 27.10 ± 1.95 | 25.60–29.30 |
| 4. | Boletus subtomentosus L. | 3 | 88.39–92.62 | 22.4 ± 3.44 | 19.8–26.3 | 4.10 ± 3.13 | 1.35–7.50 | 18.30 ± 1.75 | 16.35–19.75 |
| 5. | Cantharellus cibarius Fr. | 4 | 91.50–93.17 | 27.4 ± 1.01 | 26.5–28.3 | 1.40 ± 0.22 | 1.11–1.64 | 25.98 ± 1.20 | 24.86–27.09 |
| 6. | Lactarius deliciosus (L.) Pers. | 3 | 90.79–92.11 | 29.4 ± 0.38 | 29.1–29.8 | 1.74 ± 0.68 | 1.28–2.52 | 27.62 ± 0.30 | 27.28–27.77 |
| 7. | Leccinum aurantiacum (Bull.) Gray); | 3 | 90.50–91.53 | 32.9 ± 3.45 | 30.3–36.8 | 5.80 ± 2.79 | 3.44–8.88 | 27.06 ± 5.59 | 22.62–33.34 |
| 8. | Leccinum scabrum (Bull.) Gray) | 3 | 83.91–91.84 | 26.8 ± 2.94 | 23.6–29.4 | 3.05 ± 1.46 | 1.68–4.58 | 23.71 ± 2.45 | 21.92–26.50 |
| 9. | Macrolepiota procera (Scop.) Singer. | 3 | 82.00–87.06 | 11.4 ± 2.34 | 10.0–11.4 | 0.90 ± 0.58 | 0.23–1.25 | 10.50 ± 0.30 | 8.89–12.85 |
| 10. | Rozites caperatus (Pers.) P. Karst. | 4 | 91.15–93.26 | 33.6 ± 1.20 | 32.4–34.8 | 0.79 ± 0.24 | 0.51–0.95 | 32.78 ± 1.41 | 31.49–34.29 |
| 11. | Russula heterophylla (Fr.) Fr. | 3 | 92.50–94.83 | 35.6 ± 1.85 | 33.5–37.0 | 8.57 ± 2.64 | 6.40–11.50 | 27.03 ± 1.50 | 25.50–28.50 |
| 12. | Russula vinosa Lindblad | 3 | 86.27–90.36 | 31.8 ± 0.30 | 31.5–32.1 | 7.56 ± 4.13 | 2.80–10.20 | 24.24 ± 4.12 | 21.83–29.00 |
| 13. | Suillus bovinus (Pers.) Roussel | 3 | 82.91–85.73 | 28.4 ± 0.14 | 28.3–28.5 | 1.83 ± 0.28 | 1.52–2.06 | 26.57 ± 0.52 | 26.24–26.98 |
| 14. | Suillus grevillei (Klotzsch) Singer | 3 | 92.38–95.73 | 27.9 ± 1.06 | 27.1–29.0 | 0.70 ± 0.08 | 0.65–0.79 | 27.15 ± 1.11 | 26.22–28.38 |
| 15. | Suillus luteus (L.) Roussel | 3 | 91.32–94.22 | 29.7 ± 2.08 | 28.4–32.1 | 1.48 ± 0.06 | 1.42–1.54 | 28.22 ± 2.14 | 26.86–30.68 |
| 16. | Tricholoma equestre (L.) P. Kumm. | 3 | 86.30–92.09 | 24.2 ± 1.62 | 23.3–26.1 | 0.96 ± 0.53 | 0.55–1.56 | 23.27 ± 2.01 | 21.74–25.55 |
| 17. | Tricholoma portentosum (Fr.) Quel. | 3 | 91.59–93.36 | 36.9 ± 1.06 | 35.7–37.7 | 1.93 ± 0.15 | 1.80–2.10 | 34.97 ± 1.21 | 33.60–35.90 |
| 18. | Xerocomellus chrysenteron (Bull.) Šutara | 3 | 93.44–94.38 | 15.5 ± 2.62 | 11.3–16.5 | 1.35 ± 0.33 | 1.00–1.65 | 12.72 ± 2.94 | 9.65–15.50 |
| commercial cultivated mushrooms | |||||||||
| 19. | Lentinula edodes (Berk.) Pegler | 3 | 90.94–90.76 | 26.7 ± 3.22 | 24.74–30.40 | 0.42 ± 0.05 | 0.38–0.47 | 26.26 ± 3.23 | 24.74–30.40 |
| 20. | Pleurotus ostreatus (Jacq.) P. Kumm. | 4 | 90.93–91.91 | 45.9 ± 1.62 | 45.0–47.80 | 5.59 ± 0.62 | 5.09–6.29 | 40.34 ± 3.23 | 39.61–41.51 |
| 21. | Agaricus bisporus (J.E. Lange) Imbach) | 3 | 91.48–92.54 | 15.8 ± 3.47 | 12.60–19.50 | 4.47 ± 2.04 | 2.41–6.48 | 11.36 ± 2.85 | 8.07–13.02 |
| No. | Mushroom Species | N | β-Glucans | KOH β-Glucans Fraction a | HCl β-Glucans Fraction b | NaOH β-Glucans Fraction c |
|---|---|---|---|---|---|---|
| X ± SD g/100 g DM | ||||||
| wild edible mushrooms | ||||||
| 1. | Armillaria mellea (Vahl) P. Kumm. | 3 | 12.156 ± 1.751 | 6.205 ± 1.168 | 2.133 ± 0.206 | 3.818 ± 0.418 |
| 2. | Imleria badia (Fr.) Fr. | 4 | 9.318 ± 1.871 | 5.214 ± 1.294 | 1.551 ± 0.185 | 2.615 ± 0.418 |
| 3. | Boletus edulis Bull. | 3 | 5.801 ± 1.217 | 3.583 ± 1.097 | 1.114 ± 0.158 | 1.105 ± 0.313 |
| 4. | Boletus subtomentosus L. | 3 | 5.138 ± 1.041 | 1.797 ± 0.650 | 1.099 ± 0.075 | 2.242 ± 0.489 |
| 5. | Cantharellus cibarius Fr. | 4 | 1.946 ± 0.182 | ND | 1.010 ± 0.085 | 0.936 ± 0.103 |
| 6. | Lactarius deliciosus (L.) Pers. | 3 | 17.110 ± 1.708 | 7.423 ± 0.692 | 3.037 ± 0.390 | 6.649 ± 0.732 |
| 7. | Leccinum aurantiacum (Bull.) Gray | 3 | 8.085 ± 2.041 | 2.842 ± 0.941 | 1.287 ± 0.303 | 3.956 ± 0.831 |
| 8. | Leccinum scabrum (Bull.) Gray | 3 | 5.908 ± 0.585 | 2.213 ± 0.645 | 1.110 ± 0.192 | 2.585 ± 0.038 |
| 9. | Macrolepiota procera (Scop.) Singer. | 3 | 5.461 ± 0.601 | 1.523 ± 0.305 | 1.580 ± 0.017 | 2.357 ± 0.420 |
| 10. | Rozites caperatus (Pers.) P. Karst. | 4 | 10.026 ± 3.377 | 3.970 ± 1.653 | 1.544 ± 0.530 | 4.512 ± 1.527 |
| 11. | Russula heterophylla (Fr.) Fr. | 3 | 14.347 ± 0.812 | 6.765 ± 0.587 | 2.929 ± 0.260 | 4.653 ± 0.620 |
| 12. | Russula vinosa Lindblad | 3 | 14.665 ± 0.756 | 5.427 ± 0.459 | 3.063 ± 0.533 | 6.176 ± 0.444 |
| 13. | Suillus bovinus (Pers.) Roussel | 3 | 13.299 ± 0.651 | 9.509 ± 0.642 | 0.926 ± 0.188 | 2.864 ± 0.458 |
| 14. | Suillus grevillei (Klotzsch) Singer | 4 | 16.965 ± 1.694 | 7.810 ± 1.150 | 2.744 ± 0.207 | 6.411 ± 0.570 |
| 15. | Suillus luteus (L.) Roussel | 3 | 12.149 ± 1.043 | 8.479 ± 1.277 | 1.190 ± 0.169 | 2.480 ± 0.312 |
| 16. | Tricholoma equestre (L.) P. Kumm. | 3 | 10.323 ± 1.273 | 3.269 ± 0.630 | 1.877± | 5.178 ± 0.066 |
| 17. | Tricholoma portentosum (Fr.) Quel. | 3 | 12.912 ± 1.546 | 5.391 ± 1.136 | 2.394 ± 0.147 | 5.127 ± 0.475 |
| 18. | Xerocomellus chrysenteron (Bull.) Šutara | 3 | 5.095 ± 1.275 | 3.034 ± 1.302 | 0.644 ± 0.005 | 1.417 ± 0.035 |
| commercial mushrooms | ||||||
| 19. | Pleurotus ostreatus (Jacq.) P. Kumm. | 4 | 12.393 ± 1.100 | 4.698 ± 0.734 | 1.709 ± 0.217 | 5.986 ± 0.365 |
| 20. | Agaricus bisporus (J.E. Lange) Imbach) | 3 | 3.117 ± 0.101 | 0.879 ± 0.070 | 1.106 ± 0.049 | 1.132 ± 0.108 |
| 21. | Lentinula edodes (Berk.) Pegler | 3 | 8.417 ± 0.627 | 3.894 ± 0.523 | 1.005 ± 0.044 | 3.518 ± 0.147 |
| Glucans | Wild Grown N = 58 | Commercial Cultivated N = 10 | p | ||
|---|---|---|---|---|---|
| g/100 g DM | |||||
| The Megazyme Method | |||||
| β-glucans | Q1 | Q3 | Q1 | Q3 | ─ |
| 22.52 | 27.77 | 13.02 | 39.61 | ─ | |
| Me | Me | ─ | |||
| 26.23 | 27.25 | 0.516 | |||
| X ± SD | X ± SD | ─ | |||
| 25.01 ± 6.05 | 27.34 ± 12.76 | ─ | |||
| The Congo red method | |||||
| β-glucans (KOH fraction) | Q1 | Q3 | Q1 | Q3 | ─ |
| 2.36 | 6.65 | 0.96 | 4.39 | ─ | |
| Me | Me | ─ | |||
| 4.34 | 4.06 | 0.156 | |||
| X ± SD | X ± SD | ─ | |||
| 4.66 ± 2.71 | 3.311 ± 1.78 | ─ | |||
| β-glucans (HCl fraction) | Q1 | Q3 | Q1 | Q3 | ─ |
| 1.06 | 2.45 | 1.05 | 1.68 | ─ | |
| Me | Me | ─ | |||
| 1.51 | 1.13 | 0.236 | |||
| X ± SD | X ± SD | ─ | |||
| 1.72 ± 0.80 | 1.32 ± 0.36 | ─ | |||
| β-glucans (NaOH fraction) | Q1 | Q3 | Q1 | Q3 | ─ |
| 2.12 | 5.10 | 1.24 | 5.86 | ─ | |
| Me | Me | ─ | |||
| 3.13 | 3.59 | 0.736 | |||
| X ± SD | X ± SD | ─ | |||
| 3.60 ± 1.85 | 3.79 ± 2.14 | ─ | |||
| β-glucans (KOH fraction + HCl fraction + NaOH fraction) | Q1 | Q3 | Q1 | Q3 | ─ |
| 6.05 | 13.68 | 3.20 | 11.82 | ─ | |
| Me | Me | ─ | |||
| 10.13 | 8.75 | 0.295 | |||
| X ± SD | X ± SD | ─ | |||
| 10.02 ± 4.57 | 8.41 ± 4.11 | ─ | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirończuk-Chodakowska, I.; Witkowska, A.M. Evaluation of Polish Wild Mushrooms as Beta-Glucan Sources. Int. J. Environ. Res. Public Health 2020, 17, 7299. https://doi.org/10.3390/ijerph17197299
Mirończuk-Chodakowska I, Witkowska AM. Evaluation of Polish Wild Mushrooms as Beta-Glucan Sources. International Journal of Environmental Research and Public Health. 2020; 17(19):7299. https://doi.org/10.3390/ijerph17197299
Chicago/Turabian StyleMirończuk-Chodakowska, Iwona, and Anna Maria Witkowska. 2020. "Evaluation of Polish Wild Mushrooms as Beta-Glucan Sources" International Journal of Environmental Research and Public Health 17, no. 19: 7299. https://doi.org/10.3390/ijerph17197299
APA StyleMirończuk-Chodakowska, I., & Witkowska, A. M. (2020). Evaluation of Polish Wild Mushrooms as Beta-Glucan Sources. International Journal of Environmental Research and Public Health, 17(19), 7299. https://doi.org/10.3390/ijerph17197299