
Seaweeds as Valuable Sources of Essential Fatty Acids for Human Nutrition

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Seaweed Harvesting
2.2. Fatty Acid Analysis
2.3. Statistical Analysis
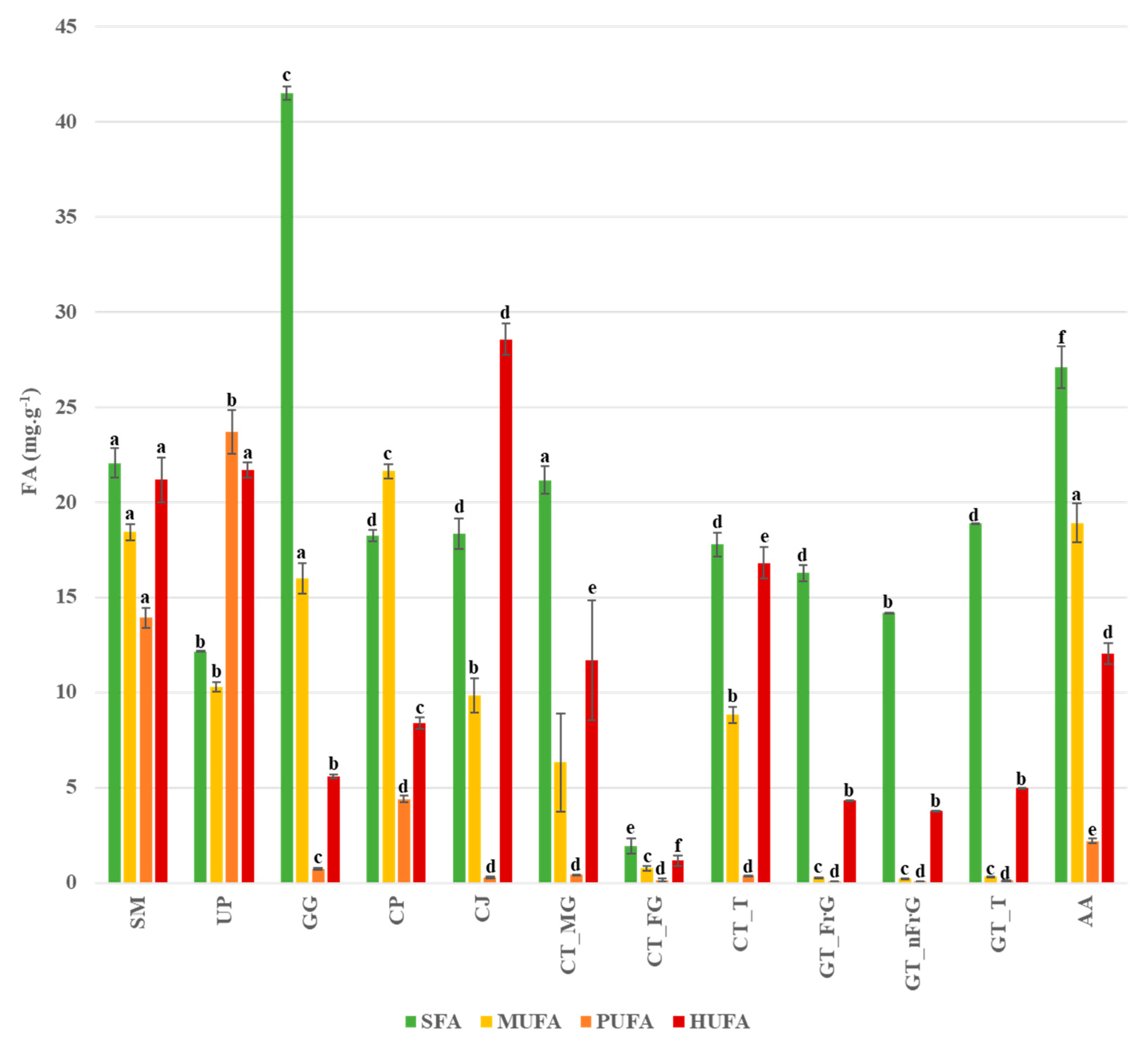
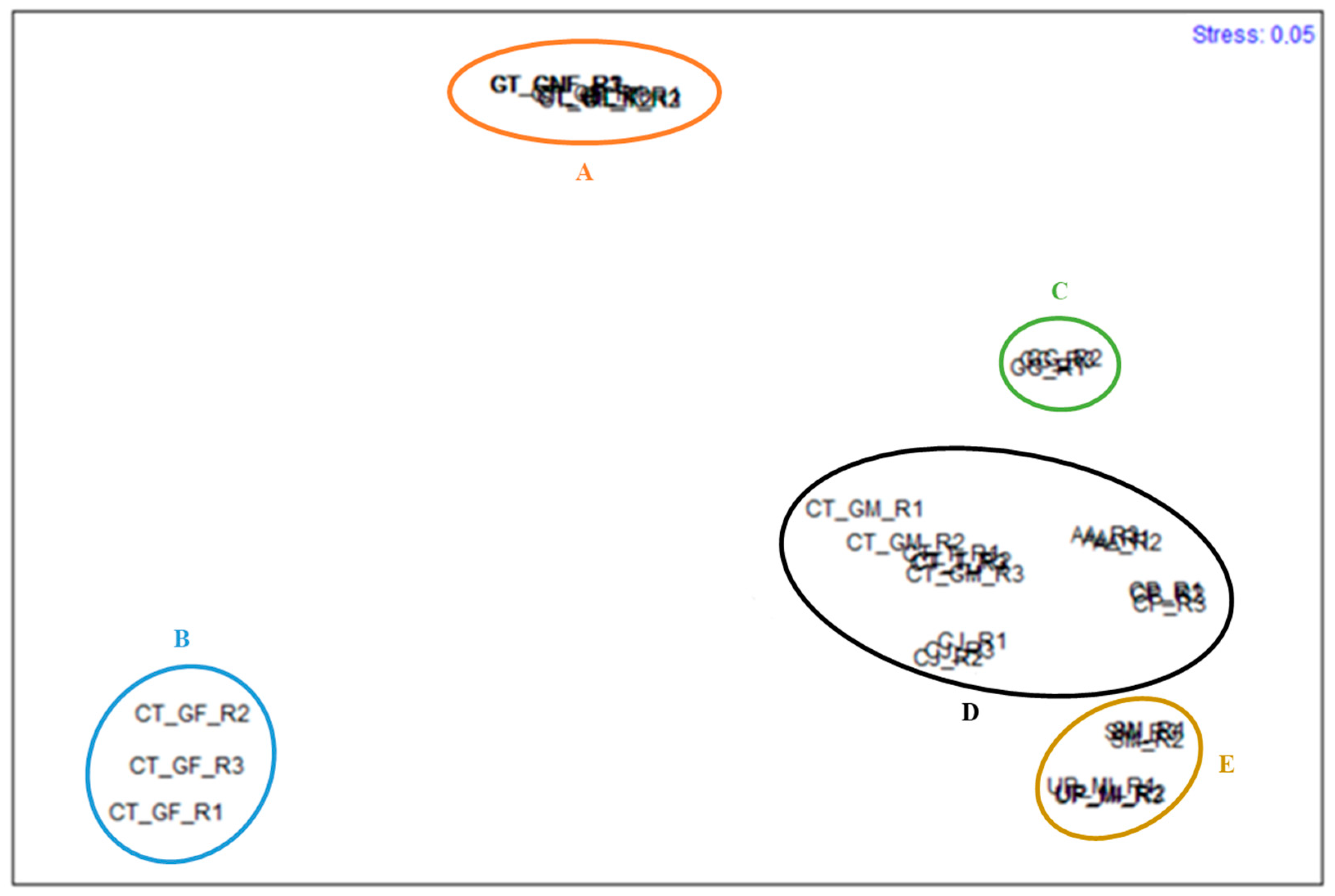
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, U.C.; Gupta, S.C. Sources and Deficiency Diseases of Mineral Nutrients in Human Health and Nutrition: A Review. Pedosphere 2020, 24, 13–38. [Google Scholar] [CrossRef]
- Simopoulos, A.P. An increase in the Omega-6/Omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8. [Google Scholar] [CrossRef] [Green Version]
- Miles, E.A.; Calder, P.C. Modulation of immune function by dietary fatty acids. Proc. Nutr. Soc. 1998, 57, 277–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccardi, G.; Giacco, R.; Rivellese, A. Dietary fat, insulin sensitivity and the metabolic syndrome. Clin. Nutr. 2004, 23, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.M.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M.M. Seaweed’s Bioactive Candidate Compounds to Food Industry and Global Food Security. Life 2020, 10, 140. [Google Scholar] [CrossRef]
- Duarte, C.; Boccardi, V.; Amaro Andrade, P.; Souza Lopes, A.C.; Jacques, P.F. Dairy versus other saturated fats source and cardiometabolic risk markers: Systematic review of randomized controlled trials. Crit. Rev. Food Sci. Nutr. 2021, 61, 450–461. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food Security: The Challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Michalak, I.; Chojnacka, K. Algae as production systems of bioactive compounds. Eng. Life Sci. 2015, 15, 160–176. [Google Scholar] [CrossRef]
- Galloway, A.W.E.; Britton-Simmons, K.H.; Duggins, D.O.; Gabrielson, P.W.; Brett, M.T. Fatty acid signatures differentiate marine macrophytes at ordinal and family ranks. J. Phycol. 2012, 48, 956–965. [Google Scholar] [CrossRef]
- Ibañez, E.; Herrero, M.; Mendiola, J.A.; Castro-Puyana, M. Extraction and Characterization of Bioactive Compounds with Health Benefits from Marine Resources: Macro and Micro Algae, Cyanobacteria, and Invertebrates. In Marine Bioactive Compounds; Springer: Boston, MA, USA, 2012; Volume 9781461412, pp. 55–98. [Google Scholar]
- Tanna, B.; Mishra, A. Metabolites Unravel Nutraceutical Potential of Edible Seaweeds: An Emerging Source of Functional Food. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1613–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Union Convention on the Conservation of European Wildlife and Natural Habitats. Environ. Conserv. 1981, 8, 206. [CrossRef]
- van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, D.; Araújo, G.S.; Cotas, J.; Gaspar, R.; Neto, J.M.; Pereira, L. Invasive Seaweeds in the Iberian Peninsula: A Contribution for Food Supply. Mar. Drugs 2020, 18, 560. [Google Scholar] [CrossRef]
- Salvaterra, T.; Green, D.S.; Crowe, T.P.; O’Gorman, E.J. Impacts of the invasive alga Sargassum muticum on ecosystem functioning and food web structure. Biol. Invasions 2013, 15, 2563–2576. [Google Scholar] [CrossRef]
- Milledge, J.J.; Nielsen, B.V.; Bailey, D. High-value products from macroalgae: The potential uses of the invasive brown seaweed, Sargassum muticum. Rev. Environ. Sci. Biotechnol. 2016, 15. [Google Scholar] [CrossRef]
- Milledge, J.; Harvey, P. Golden Tides: Problem or Golden Opportunity? The Valorisation of Sargassum from Beach Inundations. J. Mar. Sci. Eng. 2016, 4, 60. [Google Scholar] [CrossRef]
- Balboa, E.M.; Gallego-Fábrega, C.; Moure, A.; Domínguez, H. Study of the seasonal variation on proximate composition of oven-dried Sargassum muticum biomass collected in Vigo Ria, Spain. J. Appl. Phycol. 2016, 28, 1943–1953. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.M.; Pereira, L. A comprehensive review of the nutraceutical and therapeutic applications of red seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Poza, S.; Leandro, A.; Cotas, C.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. The Evolution Road of Seaweed Aquaculture: Cultivation Technologies and the Industry 4.0. Int. J. Environ. Res. Public Health 2020, 17, 6528. [Google Scholar] [CrossRef]
- García-Jiménez, P.; Robaina, R.R. On reproduction in red algae: Further research needed at the molecular level. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Katsanevakis, S.; Acar, Ü.; Ammar, I.; Balci, B.A.; Bekas, P.; Belmonte, M.; Chintiroglou, C.C.; Consoli, P.; Dimiza, M.; Fryganiotis, K.; et al. New Mediterranean Biodiversity Records (October, 2014). Mediterr. Mar. Sci. 2014, 15, 675. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Silva, P. A concise review of the red macroalgae Chondracanthus teedei (Mertens ex Roth) Kützing and Chondracanthus teedei var. lusitanicus (J.E. De Mesquita Rodrigues) Bárbara & Cremades. J. Appl. Phycol. 2021, 33, 111–131. [Google Scholar] [CrossRef]
- Cardoso, I.; Cotas, J.; Rodrigues, A.; Ferreira, D.; Osório, N.; Pereira, L. Extraction and Analysis of Compounds with Antibacterial Potential from the Red Alga Grateloupia turuturu. J. Mar. Sci. Eng. 2019, 7, 220. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L. Population Studies and Carrageenan Properties in Eight Gigartinales (Rhodophyta) from Western Coast of Portugal. Sci. World J. 2013, 2013, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Mesquita, J.F. Carrageenophytes of occidental Portuguese coast: 1-spectroscopic analysis in eight carrageenophytes from Buarcos bay. Biomol. Eng. 2003, 20, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Soares, F.; Fernandes, C.; Silva, P.; Pereira, L.; Gonçalves, T. Antifungal activity of carrageenan extracts from the red alga Chondracanthus teedei var. lusitanicus. J. Appl. Phycol. 2016, 28, 2991–2998. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef] [Green Version]
- De Alencar, D.B.; Diniz, J.C.; Rocha, S.A.S.; Pires-Cavalcante, K.M.S.; De Lima, R.L.; De Sousa, K.C.; Freitas, J.O.; Bezerra, R.M.; Baracho, B.M.; Sampaio, A.H.; et al. Fatty acid composition from the marine red algae Pterocladiella capillacea (S. G. Gmelin) Santelices & Hommersand 1997 and Osmundaria obtusiloba (C. Agardh) R. E. Norris 1991 and its antioxidant activity. An. Acad. Bras. Cienc. 2018, 90, 449–459. [Google Scholar] [CrossRef]
- Kendel, M.; Wielgosz-Collin, G.; Bertrand, S.; Roussakis, C.; Bourgougnon, N.B.; Bedoux, G. Lipid composition, fatty acids and sterols in the seaweeds Ulva armoricana, and Solieria chordalis from brittany (France): An analysis from nutritional, chemotaxonomic, and antiproliferative activity perspectives. Mar. Drugs 2015. [CrossRef] [PubMed]
- Zárate, R.; Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Gonçalves, A.M.M.; Azeiteiro, U.M.; Pardal, M.A.; De Troch, M. Fatty acid profiling reveals seasonal and spatial shifts in zooplankton diet in a temperate estuary. Estuar. Coast. Shelf Sci. 2012, 109, 70–80. [Google Scholar] [CrossRef]
- Fomenko, S.E.; Kushnerova, N.F.; Sprygin, V.G.; Drugova, E.S.; Lesnikova, L.N.; Merzlyakov, V.Y.; Momot, T.V. Lipid Composition, Content of Polyphenols, and Antiradical Activity in Some Representatives of Marine Algae. Russ. J. Plant Physiol. 2019, 66, 942–949. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Engelen, A.; Varela, J. Polyunsaturated Fatty Acids of Marine Macroalgae: Potential for Nutritional and Pharmaceutical Applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [Green Version]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Fatty acid contents and profiles of 16 macroalgae collected from the Irish Coast at two seasons. J. Appl. Phycol. 2014, 26, 451–463. [Google Scholar] [CrossRef]
- Rohani-Ghadikolaei, K.; Abdulalian, E.; Ng, W.-K. Evaluation of the proximate, fatty acid and mineral composition of representative green, brown and red seaweeds from the Persian Gulf of Iran as potential food and feed resources. J. Food Sci. Technol. 2012, 49, 774–780. [Google Scholar] [CrossRef] [Green Version]
- Menaa, F.; Wijesinghe, P.A.U.I.; Thiripuranathar, G.; Uzair, B.; Iqbal, H.; Khan, B.A.; Menaa, B. Ecological and Industrial Implications of Dynamic Seaweed-Associated Microbiota Interactions. Mar. Drugs 2020, 18, 641. [Google Scholar] [CrossRef]
- Khotimchenko, S.V.; Vaskovsky, V.E.; Titlyanova, T.V. Fatty Acids of Marine Algae from the Pacific Coast of North California. Bot. Mar. 2002, 45. [Google Scholar] [CrossRef]
- Gosch, B.J.; Paul, N.A.; de Nys, R.; Magnusson, M. Seasonal and within-plant variation in fatty acid content and composition in the brown seaweed Spatoglossum macrodontum (Dictyotales, Phaeophyceae). J. Appl. Phycol. 2015, 27, 387–398. [Google Scholar] [CrossRef]
- Farnham, W.F.; Fletcher, R.L.; Irvine, L.M. Attached Sargassum found in Britain. Nature 1973, 243, 2. [Google Scholar] [CrossRef]
- Critchley, A.T.; Farnham, W.F.; Morrell, S.L. A chronology of new European sites of attachment for the invasive brownalga, Sargassum muticum, 1973–1981. J. Mar. Biol. Assoc. UK 1983, 63, 799–811. [Google Scholar] [CrossRef]
- Rueness, J. Sargassum muticum and other introduced Japanese macroalgae: Biological pollution of European coasts. Mar. Pollut. Bull. 1989, 20, 173–176. [Google Scholar] [CrossRef]
- Fouda, W.A.; Ibrahim, W.M.; Ellamie, A.M.; Ramadan, G. Biochemical and mineral compositions of six brown seaweeds collected from red sea at hurghada coast. Indian J. Geo-Marine Sci. 2019, 48, 484–491. [Google Scholar]
- Lodeiro, P.; Cordero, B.; Grille, Z.; Herrero, R.; Sastre de Vicente, M.E. Physicochemical studies of Cadmium(II) biosorption by the invasive alga in Europe, Sargassum muticum. Biotechnol. Bioeng. 2004, 88, 237–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, F.; Monteiro, J.P.; Duarte, D.; Melo, T.; Lopes, D.; da Costa, E.; Domingues, M.R. Unraveling the Lipidome and Antioxidant Activity of Native Bifurcaria bifurcata and Invasive Sargassum muticum Seaweeds: A Lipid Perspective on How Systemic Intrusion May Present an Opportunity. Antioxidants 2020, 9, 642. [Google Scholar] [CrossRef] [PubMed]
- Debbarma, J.; Madhusudana Rao, B.; Murthy, L.N.; Mathew, S.; Venkateshwarlu, G.; Ravishankar, C.N. Nutritional profiling of the edible seaweeds Gracilaria edulis, Ulva lactuca and Sargassum sp. Indian J. Fish. 2016, 63. [Google Scholar] [CrossRef] [Green Version]
- Ferdouse, F.; Løvstad Holdt, S.; Smith, R.; Murúa, P.; Yang, Z. The global status of seaweed production, trade and utilization. FAO Globefish Res. Program. 2018, 124, 120. [Google Scholar]
- Taboada, C.; Millan, R.; Miguez, I. Evaluation of marine algae Undaria pinnatifida and Porphyra purpurea as a food supplement: Composition, nutritional value and effect of intake on intestinal, hepatic and renal enzyme activities in rats. J. Sci. Food Agric. 2013, 93, 1863–1868. [Google Scholar] [CrossRef] [PubMed]
- Araújo, R.; Vázquez Calderón, F.; Sánchez López, J.; Azevedo, I.C.; Bruhn, A.; Fluch, S.; Garcia Tasende, M.; Ghaderiardakani, F.; Ilmjärv, T.; Laurans, M.; et al. Current Status of the Algae Production Industry in Europe: An Emerging Sector of the Blue Bioeconomy. Front. Mar. Sci. 2021, 7. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: https://www.algaebase.org/search/species/detail/?species_id=27676 (accessed on 11 January 2021).
- Dellatorre, F.G.; Avaro, M.G.; Commendatore, M.G.; Arce, L.; Díaz de Vivar, M.E. The macroalgal ensemble of Golfo Nuevo (Patagonia, Argentina) as a potential source of valuable fatty acids for nutritional and nutraceutical purposes. Algal Res. 2020, 45, 101726. [Google Scholar] [CrossRef]
- Fleurence, J.; Gutbier, G.; Mabeau, S.; Leray, C. Fatty acids from 11 marine macroalgae of the French Brittany coast. J. Appl. Phycol. 1994, 6, 527–532. [Google Scholar] [CrossRef]
- Cofrades, S.; López-Lopez, I.; Bravo, L.; Ruiz-Capillas, C.; Bastida, S.; Larrea, M.T.; Jiménez-Colmenero, F. Nutritional and Antioxidant Properties of Different Brown and Red Spanish Edible Seaweeds. Food Sci. Technol. Int. 2010, 16, 361–370. [Google Scholar] [CrossRef]
- Abbott, I.A.; Isabella, A.; Hollenberg, G.J. Marine Algae of California; Stanford University Press: Palo Alto, CA, USA, 1992. [Google Scholar]
- Blackler, H. The occurrence of Colpomenia peregrina (Sauv.) Hamel in the Mediterranean (Phaeophyta, Scytosiphonales). Blumea Biodivers. Evol. Biogeogr. Plants 1967, 15, 5–8. [Google Scholar]
- Green, L.A.; Mathieson, A.C.; Neefus, C.D.; Traggis, H.M.; Dawes, C.J. Southern expansion of the brown alga Colpomenia peregrina Sauvageau (Scytosiphonales) in the Northwest Atlantic Ocean. Bot. Mar. 2012, 55, 643–647. [Google Scholar] [CrossRef] [Green Version]
- Beacham, T.A.; Cole, I.S.; DeDross, L.S.; Raikova, S.; Chuck, C.J.; Macdonald, J.; Herrera, L.; Ali, T.; Airs, R.L.; Landels, A.; et al. Analysis of Seaweeds from South West England as a Biorefinery Feedstock. Appl. Sci. 2019, 9, 4456. [Google Scholar] [CrossRef] [Green Version]
- Francavilla, M.; Franchi, M.; Monteleone, M.; Caroppo, C. The red seaweed Gracilaria gracilis as a multi products source. Mar. Drugs 2013, 11, 3754–3776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, S.; Pereira, R.; Oliveira, B.; Baião, L.F.; Jessen, F.; Tulli, F.; Messina, M.; Silva, J.L.; Abreu, H.; Valente, L.M.P. Exploring the potential of seaweed Gracilaria gracilis and microalga Nannochloropsis oceanica, single or blended, as natural dietary ingredients for European seabass Dicentrarchus labrax. J. Appl. Phycol. 2020, 32, 2041–2059. [Google Scholar] [CrossRef]
- Wilson, A.J.; Critchley, A.T. Studies on Gracilaria gracilis (Stackhouse) Steentoft, Farnham and Irvine and Gracilaria aculeata (Hering) Papenfuss from southern Africa. I. The influence of temperature, irradiance, salinity and nitrogen-nutrition on growth. South African J. Bot. 1997, 63, 465–473. [Google Scholar] [CrossRef] [Green Version]
- Ben Said, R.; Mensi, F.; Majdoub, H.; Ben Said, A.; Ben Said, B.; Bouraoui, A. Effects of depth and initial fragment weights of Gracilaria gracilis on the growth, agar yield, quality, and biochemical composition. J. Appl. Phycol. 2018, 30, 2499–2512. [Google Scholar] [CrossRef]
- Capillo, G.; Savoca, S.; Costa, R.; Sanfilippo, M.; Rizzo, C.; Lo Giudice, A.; Albergamo, A.; Rando, R.; Bartolomeo, G.; Spanò, N.; et al. New Insights into the Culture Method and Antibacterial Potential of Gracilaria gracilis. Mar. Drugs 2018, 16, 492. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L. Edible Seaweeds of the World; Pereira, L., Ed.; CRC Press: Boca Raton, FL, USA, 2016; ISBN 9780429154041. [Google Scholar]
- Araujo, G.S.; Cotas, J.; Morais, T.; Leandro, A.; García-Poza, S.; Gonçalves, A.M.M.; Pereira, L. Calliblepharis jubata Cultivation Potential—A Comparative Study between Controlled and Semi-Controlled Aquaculture. Appl. Sci. 2020, 10, 7553. [Google Scholar] [CrossRef]
- Bert, J.J.; Dauget, J.C.; Maume, D.; Bert, M. Fatty-acids and sterols of 2 red algae Calliblepharis jubata and Solieria chordalis (Gigartinales). Cryptogam. Algol. 1991, 12, 157–162. [Google Scholar]
- Horridge, G.A. Occurrence of Asparagopsis armata harv. on the scilly Isles. Nature 1951, 167, 732–733. [Google Scholar] [CrossRef]
- Kraan, S.; Barrington, K.A. Commercial farming of Asparagopsis armata (Bonnemaisoniceae, Rhodophyta) in Ireland, maintenance of an introduced species? J. Appl. Phycol. 2005, 17, 103–110. [Google Scholar] [CrossRef]
- Mac Monagail, M.; Morrison, L. The seaweed resources of Ireland: A twenty-first century perspective. J. Appl. Phycol. 2020, 1–14. [Google Scholar] [CrossRef]
- Pellegrini, M.; Pharo, P.; Sciences, F. Contribution ä etude chimique des Algues Mediterraneennes. Botanica Marina 1967, 9–14. [Google Scholar] [CrossRef]
- Rodrigues, D.; Freitas, A.C.; Pereira, L.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition of red, brown and green macroalgae from Buarcos bay in Central West Coast of Portugal. Food Chem. 2015, 183, 197–207. [Google Scholar] [CrossRef]
- Milinovic, J.; Rodrigues, C.; Diniz, M.; Noronha, J.P. Determination of total iodine content in edible seaweeds: Application of inductively coupled plasma-atomic emission spectroscopy. Algal Res. 2021, 53, 102149. [Google Scholar] [CrossRef]
- Pereira, L.; van de Velde, F. Portuguese carrageenophytes: Carrageenan composition and geographic distribution of eight species (Gigartinales, Rhodophyta). Carbohydr. Polym. 2011, 84, 614–623. [Google Scholar] [CrossRef] [Green Version]
- Pereira, L.; Critchley, A.T.; Amado, A.M.; Ribeiro-Claro, P.J.A. A comparative analysis of phycocolloids produced by underutilized versus industrially utilized carrageenophytes (Gigartinales, Rhodophyta). J. Appl. Phycol. 2009, 21, 599–605. [Google Scholar] [CrossRef]
- Levring, T.; Hoppe, H.A.; Schmid, O.J. Marinae Algae, A Survey of Research and Utilization. Fette Seifen Anstrichm. 1969, 71, 1020. [Google Scholar] [CrossRef]
- Fujiwara-Arasaki, T.; Mino, N.; Kuroda, M. The protein value in human nutrition of edible marine algae in Japan. In Eleventh International Seaweed Symposium; Springer: Berlin, Germany, 1984; pp. 513–516. [Google Scholar]
- Denis, C.; Morançais, M.; Li, M.; Deniaud, E.; Gaudin, P.; Wielgosz-Collin, G.; Barnathan, G.; Jaouen, P.; Fleurence, J. Study of the chemical composition of edible red macroalgae Grateloupia turuturu from Brittany (France). Food Chem. 2010, 119, 913–917. [Google Scholar] [CrossRef]
- Kendel, M.; Couzinet-Mossion, A.; Viau, M.; Fleurence, J.; Barnathan, G.; Wielgosz-Collin, G. Seasonal composition of lipids, fatty acids, and sterols in the edible red alga Grateloupia turuturu. J. Appl. Phycol. 2013, 25, 425–432. [Google Scholar] [CrossRef]
- Munier, M.; Dumay, J.; Morançais, M.; Jaouen, P.; Fleurence, J. Variation in the Biochemical Composition of the Edible Seaweed Grateloupia turuturu Yamada Harvested from Two Sampling Sites on the Brittany Coast (France): The Influence of Storage Method on the Extraction of the Seaweed Pigment R-Phycoerythrin. J. Chem. 2013, 2013. [Google Scholar] [CrossRef] [Green Version]
- Kendel, M.; Barnathan, G.; Fleurence, J.; Rabesaotra, V.; Wielgosz-Collin, G. Non-methylene Interrupted and Hydroxy Fatty Acids in Polar Lipids of the Alga Grateloupia turuturu Over the Four Seasons. Lipids 2013, 48, 535–545. [Google Scholar] [CrossRef]
- Peñalver, R.; Lorenzo, J.M.; Ros, G.; Amarowicz, R.; Pateiro, M.; Nieto, G. Seaweeds as a Functional Ingredient for a Healthy Diet. Mar. Drugs 2020, 18, 301. [Google Scholar] [CrossRef] [PubMed]
- Innis, S.M. Fatty acids and early human development. Early Hum. Dev. 2007, 83, 761–766. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Omega-3 Fatty Acids in Inflammation and Autoimmune Diseases. J. Am. Coll. Nutr. 2002, 21, 495–505. [Google Scholar] [CrossRef]
- Yazici, Z.; Aysel, V.; Öksüz, E.; Köse, A.; Cumali, S.; Güven, K.C. Fatty acid composition of marine macroalgae from the Black Sea and Dardanelles. Toxicol. Environ. Chem. 2007, 89, 371–379. [Google Scholar] [CrossRef]
- Soler-Vila, A.; Coughlan, S.; Guiry, M.D.; Kraan, S. The red alga Porphyra dioica as a fish-feed ingredient for rainbow trout (Oncorhynchus mykiss): Effects on growth, feed efficiency, and carcass composition. J. Appl. Phycol. 2009, 21, 617–624. [Google Scholar] [CrossRef]
- Norambuena, F.; Hermon, K.; Skrzypczyk, V.; Emery, J.A.; Sharon, Y.; Beard, A.; Turchini, G.M. Algae in Fish Feed: Performances and Fatty Acid Metabolism in Juvenile Atlantic Salmon. PLoS ONE 2015, 10, e0124042. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion related to the Tolerable Upper Intake Level of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA). EFSA J. 2012, 10, 2815. [Google Scholar] [CrossRef]
- Institute of Medicine of the National Academies. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein and Amino Acids. 2005. Available online: https://www.nal.usda.gov/sites/default/files/fnic_uploads/energy_full_report.pdf (accessed on 30 April 2021).
- Simopoulos, A.P.; DiNicolantonio, J.J. The importance of a balanced ω-6 to ω-3 ratio in the prevention and management of obesity. Open Hear. 2016, 3, e000385. [Google Scholar] [CrossRef] [Green Version]
- DiNicolantonio, J.J.; O’Keefe, J.H. Importance of maintaining a low omega–6/omega–3 ratio for reducing inflammation. Open Hear. 2018, 5, e000946. [Google Scholar] [CrossRef]
- Joint FAO/WHO Expert Consultation. Fats and Fatty Acids in Human Nutrition. Ann. Nutr. Metab. 2008, 55. [Google Scholar]
- Simopoulos, A.P. The Importance of the Omega-6/Omega-3 Fatty Acid Ratio in Cardiovascular Disease and Other Chronic Diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Lee, J.H.; Fukumoto, M.; Nishida, H.; Ikeda, I.; Sugano, M. The Interrelated Effects of n-6/n-3 and Polyunsaturated/Saturated Ratios of Dietary Fats on the Regulation of Lipid Metabolism in Rats. J. Nutr. 1989, 119, 1893–1899. [Google Scholar] [CrossRef]
- van Ginneken, V.J.; Helsper, J.P.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Kubo, K. Relationship between tissue lipid peroxidation and peroxidizability index after α-linolenic, eicosapentaenoic, or docosahexaenoic acid intake in rats. Br. J. Nutr. 2003, 89, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Roque, B.M.; Salwen, J.K.; Kinley, R.; Kebreab, E. Inclusion of Asparagopsis armata in lactating dairy cows’ diet reduces enteric methane emission by over 50 percent. J. Clean. Prod. 2019, 234, 132–138. [Google Scholar] [CrossRef]
- Kolb, N.; Vallorani, L.; Milanovi’c, N.; Stocchi, V. Evaluation of Marine AlgaeWakame (Undaria pinnatifida) and Kombu (Laminaria digitata japonica) as Food Supplements. Food Technol. Biotechnol. 2004, 42, 57–61. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Seaweed Species | Location | GPS Location | Harvesting Date |
|---|---|---|---|
| Rhodophyta (red seaweed) | |||
| Asparagopsis armata * | Quebrado Beach | 39.368258, −9.372303 | 20/10/2020 |
| Calliblepharis jubata | Buarcos Bay | 40.165867, −8.885556 | 19/10/2020 |
| Chondracanthus teedei var. lusitanicus | Buarcos Bay | 40.165867, −8.885556 | 27/05/2020 |
| Gracilaria gracilis | Buarcos Bay | 40.165867, −8.885556 | 19/10/2020 |
| Grateloupia turuturu * | Buarcos Bay | 40.165867, −8.885556 | 13/01/2020 |
| Ochrophyta (brown seaweed) | |||
| Colpomenia peregrine * | Quebrado Beach | 39.368258, −9.372303 | 20/10/2020 |
| Sargassum muticum * | Buarcos Bay | 40.165867, −8.885556 | 19/10/2020 |
| Undaria pinnatifida * | Buarcos Bay | 40.165867, −8.885556 | 13/01/2020 |
| Species | Moisture (%) |
|---|---|
| S. muticum | 87.22 |
| U. pinnatifida | 86.84 |
| C. peregrina | 91.25 |
| G. gracilis | 49.49 |
| C. jubata | 89.13 |
| A. armata | 87.22 |
| C. teedei (MG) | 85.86 |
| C. teedei (FG) | 85.43 |
| C. teedei (T) | 84.35 |
| G. turuturu (FrG) | 86.78 |
| G. turuturu (nFrG) | 87.69 |
| G. turuturu (T) | 88.24 |
| S. muticum | U. pinnatifida | C. peregrina | G. gracilis | C. jubata | A. armata | |
|---|---|---|---|---|---|---|
| C16:0 | 20.89 ± 0.72 | 11.51 ± 0.01 | 18.24 ± 0.31 | 40.46 ± 0.23 | 15.94 ± 0.73 | 26.39 ± 1.19 |
| C17:0 | 0.29 ± 0.03 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 2.24 ± 0.05 | 0.00 ± 0.00 |
| C18:0 | 0.43 ± 0.03 | 0.64 ± 0.02 | 0.00 ± 0.00 | 1.03 ± 0.13 | 0.17 ± 0.03 | 0.71 ± 0.10 |
| C24:0 | 0.45 ± 0.05 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ∑ SFA | 22.06 | 12.15 | 18.24 | 41.49 | 18.35 | 27.10 |
| C15:1 | 3.15 ± 0.15 | 2.44 ± 0.10 | 7.72 ± 0.14 | 5.64 ± 0.47 | 3.28 ± 0.14 | 8.53 ± 0.29 |
| C16:1 | 7.43 ± 0.15 | 1.66 ± 0.06 | 2.66 ± 0.05 | 1.13 ± 0.09 | 2.14 ± 0.20 | 3.60 ± 0.18 |
| C18:1 | 7.84 ± 0.24 | 6.21 ± 0.11 | 11.24 ± 0.28 | 9.21 ± 0.23 | 4.42 ± 0.63 | 6.77 ± 0.56 |
| ∑ MUFA | 18.42 | 10.31 | 21.62 | 15.98 | 9.83 | 18.90 |
| C18:2 | 4.67 ± 0.23 | 3.87 ± 0.08 | 0.96 ± 0.06 | 0.74 ± 0.06 | 0.27 ± 0.05 | 0.78 ± 0.04 |
| C18:3 (α-LA) | 6.23 ± 0.23 | 7.51 ± 0.42 | 1.53 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.80 ± 0.09 |
| C18:3 (γ-LA) | 3.01 ± 0.24 | 12.33 ± 0.67 | 1.92 ± 0.09 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.62 ± 0.01 |
| ∑ PUFA | 13.92 | 23.71 | 4.40 | 0.74 | 0.27 | 2.21 |
| C20:4 (ARA) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.77 ± 0.05 |
| C20:5 (EPA) | 13.83 ± 0.48 | 13.15 ± 0.02 | 2.59 ± 0.00 | 5.58 ± 0.09 | 6.22 ± 0.22 | 2.43 ± 0.01 |
| C22:6 (DHA) | 7.33 ± 0.72 | 8.55 ± 0.37 | 5.80 ± 0.29 | 0.00 ± 0.00 | 22.34 ± 0.63 | 8.83 ± 0.49 |
| ∑ HUFA | 21.17 | 21.70 | 8.39 | 5.58 | 28.56 | 12.03 |
| ∑ FA | 75.56 | 67.86 | 52.65 | 63.80 | 57.01 | 60.23 |
| ω-6/ω-3 | 0.15 | 0.09 | 0.08 | 0.13 | 0.01 | 0.12 |
| N | 12 | 10 | 9 | 7 | 9 | 11 |
| C. teedei (MG) | C. teedei (FG) | C. teedei (T) | G. turuturu (FrG) | G. turuturu (nFrG) | G. turuturu (T) | |
|---|---|---|---|---|---|---|
| C16:0 | 20.29 ± 8.45 | 1.66 ± 4.36 | 17.46 ± 0.63 | 13.59 ± 14.20 | 11.84 ± 5.65 | 15.75 ± 5.41 |
| C17:0 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 1.08 |
| C18:0 | 0.50 ± 0.04 | 0.12 ± 0.07 | 0.32 ± 0.02 | 2.69 ± 0.49 | 2.34 ± 0.00 | 3.12 ± 0.10 |
| C24:0 | 0.26 ± 0.20 | 0.15 ± 0.04 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| ∑ SFA | 21.16 | 1.93 | 17.78 | 16.28 | 14.18 | 18.87 |
| C15:1 | 2.41 ± 0.93 | 0.21 ± 0.48 | 2.63 ± 0.34 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| C16:1 | 0.84 ± 0.03 | 0.12 ± 0.10 | 1.20 ± 0.06 | 0.05 ± 0.84 | 0.05 ± 0.88 | 0.06 ± 0.81 |
| C18:1 | 3.08 ± 2.71 | 0.44 ± 1.29 | 4.99 ± 0.28 | 0.23 ± 3.02 | 0.20 ± 3.51 | 0.26 ± 3.37 |
| ∑ MUFA | 6.32 | 0.77 | 8.82 | 0.28 | 0.24 | 0.33 |
| C18:2 | 0.41 ± 0.01 | 0.14 ± 0.13 | 0.38 ± 0.02 | 0.07 ± 0.27 | 0.06 ± 0.34 | 0.08 ± 0.35 |
| C18:3 (α-LA) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.02 ± 0.64 | 0.02 ± 0.52 | 0.03 ± 0.65 |
| C18:3 (γ-LA) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.77 | 0.00 ± 0.72 | 0.00 ± 0.88 |
| ∑ PUFA | 0.41 | 0.14 | 0.38 | 0.09 | 0.08 | 0.11 |
| C20:4 ω-6 (ARA) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.24 ± 0.00 | 0.21 ± 0.00 | 0.28 ± 0.00 |
| C20:5 ω-3 (EPA) | 2.66 ± 2.35 | 0.40 ± 1.11 | 6.00 ± 0.31 | 4.08 ± 2.01 | 3.55 ± 0.89 | 4.73 ± 2.33 |
| C22:6 ω-3 (DHA) | 5.16 ± 4.54 | 0.76 ± 2.42 | 10.81 ± 0.55 | 0.00 ± 2.27 | 0.00 ± 1.96 | 0.00 ± 9.07 |
| ∑ HUFA | 11.70 | 1.16 | 16.81 | 4.32 | 3.76 | 5.01 |
| ∑ FA | 39.59 | 4.01 | 43.79 | 30.21 | 26.33 | 35.01 |
| ω-6/ω-3 | 0.04 | 0.12 | 0.02 | 0.08 | 0.08 | 0.08 |
| N | 9 | 9 | 8 | 8 | 8 | 8 |
| Groups | Av. diss. between Groups | Main FA | % Contribution to diss. | % Cumulative Contribution to diss. |
|---|---|---|---|---|
| D,E | 37.98 | EPA | 19.96 | 49.75 |
| γ-LA | 15.85 | |||
| α-LA | 13.94 | |||
| D,C | 38.58 | C16:0 | 47.58 | 81.64 |
| DHA | 24.89 | |||
| C18:1 | 9.17 | |||
| E,C | 50.01 | C16:0 | 36.05 | 59.46 |
| DHA | 11.74 | |||
| EPA | 11.67 | |||
| D,B | 85.65 | C16:0 | 39.13 | 72.93 |
| DHA | 21.86 | |||
| C18:1 | 11.94 | |||
| E,B | 89.62 | C16:0 | 21.14 | 52.14 |
| EPA | 19.32 | |||
| γ-LA | 11.67 | |||
| C,B | 90.88 | C16:0 | 63.01 | 86.06 |
| C18:1 | 14.24 | |||
| C15:1 | 8.81 | |||
| D,A | 45.84 | DHA | 29.60 | 58.05 |
| C16:0 | 15.38 | |||
| EPA | 13.08 | |||
| E,A | 53.45 | EPA | 15.48 | 44.40 |
| DHA | 14.56 | |||
| γ-LA | 14.35 | |||
| C,A | 51.04 | C16:0 | 54.82 | 79.58 |
| C18:1 | 14.44 | |||
| EPA | 10.33 | |||
| B,A | 82.71 | C16:0 | 45.61 | 75.62 |
| EPA | 21.40 | |||
| ARA | 8.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rocha, C.P.; Pacheco, D.; Cotas, J.; Marques, J.C.; Pereira, L.; Gonçalves, A.M.M. Seaweeds as Valuable Sources of Essential Fatty Acids for Human Nutrition. Int. J. Environ. Res. Public Health 2021, 18, 4968. https://doi.org/10.3390/ijerph18094968
Rocha CP, Pacheco D, Cotas J, Marques JC, Pereira L, Gonçalves AMM. Seaweeds as Valuable Sources of Essential Fatty Acids for Human Nutrition. International Journal of Environmental Research and Public Health. 2021; 18(9):4968. https://doi.org/10.3390/ijerph18094968
Chicago/Turabian StyleRocha, Carolina P., Diana Pacheco, João Cotas, João C. Marques, Leonel Pereira, and Ana M. M. Gonçalves. 2021. "Seaweeds as Valuable Sources of Essential Fatty Acids for Human Nutrition" International Journal of Environmental Research and Public Health 18, no. 9: 4968. https://doi.org/10.3390/ijerph18094968
APA StyleRocha, C. P., Pacheco, D., Cotas, J., Marques, J. C., Pereira, L., & Gonçalves, A. M. M. (2021). Seaweeds as Valuable Sources of Essential Fatty Acids for Human Nutrition. International Journal of Environmental Research and Public Health, 18(9), 4968. https://doi.org/10.3390/ijerph18094968