Effect of Temperature on Growth of Vibrio paraphemolyticus and Vibrio vulnificus in Flounder, Salmon Sashimi and Oyster Meat


Abstract
:1. Introduction
2. Experimental Section
2.1. Bacteria Strains
2.2. Preparation of Inoculation in Broth and Sashimi Samples
2.3. Isolation of Natural V. parahaemolyticus and V. vulnificus in Shucked Oysters
2.4. Determination of Growth Kinetics of V. vulnificus in Shucked Oyster Meat
2.5. Effect of Temperature on Growth Kinetics of V. parahaemolyticus and V. vulnificus.



2.6. Statistical Analysis
3. Results and Discussion
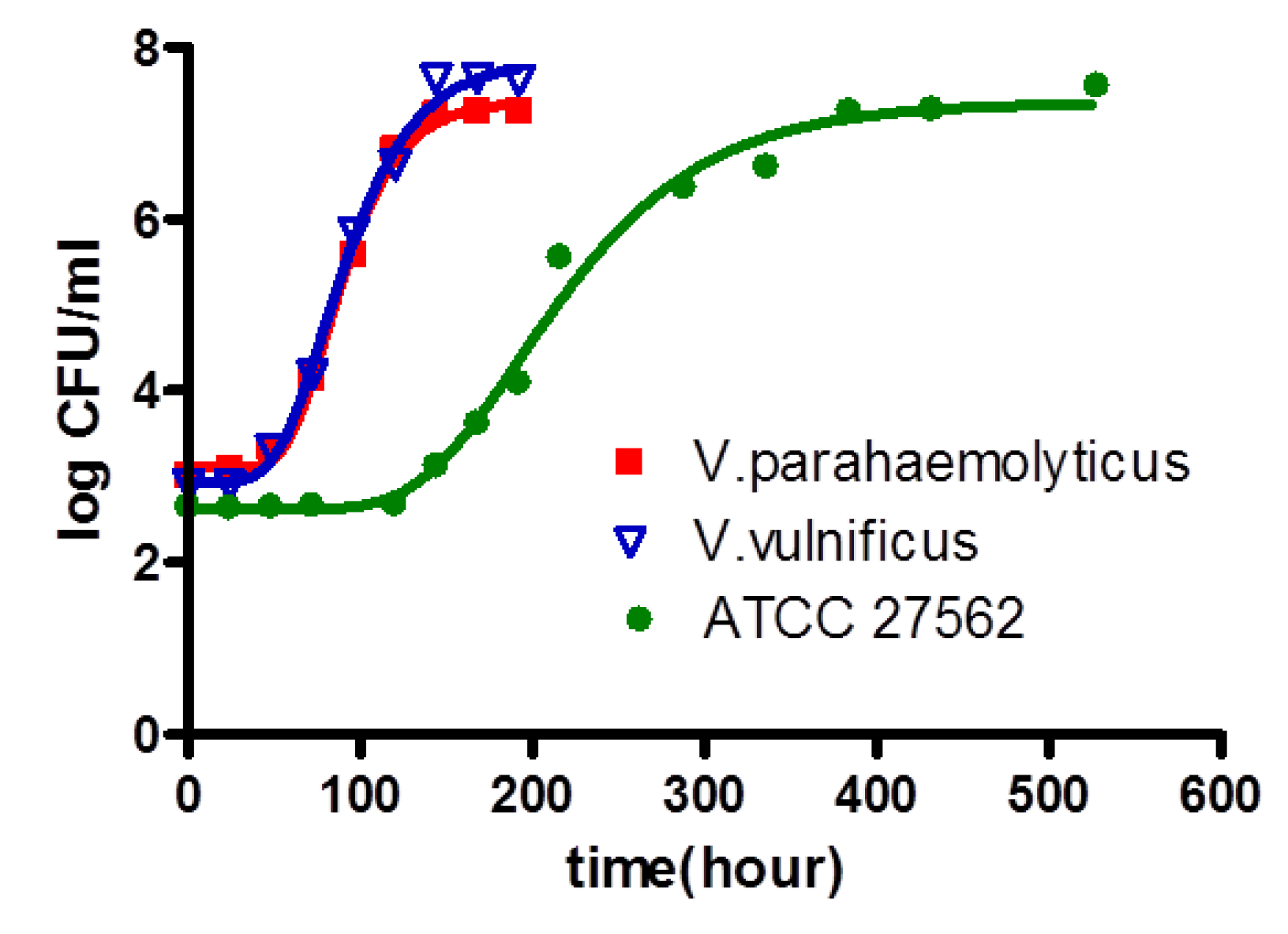
3.1. Comparison of Growth Kinetics of V. parahaemolyticus and V. vulnificus in Broth

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 11 °C | 13 °C | 18 °C | 24 °C | 30 °C | 36 °C | |
|---|---|---|---|---|---|---|---|
| LT (h) | V. parahaemolyticus | - | 48.64 ± 5.23 A | 16.94 ± 2.33 B | 2.52 ± 0.16 C | 2.22 ± 0.33 C | 1.61 ± 0.87 C.D |
| V. vulnificus | 127.80 ± 4.97 A | - | 10.49 ± 3.17 B | 4.60 ± 1.09 C | 2.40 ± 0.72 D | 1.46 ± 0.23 E | |
| SGR (log/h) | V. parahaemolyticus | - | 0.035 ± 0.04 E | 0.160 ± 0.14 D | 0.561 ± 0.02 C | 0.847 ± 0.04 B | 1.184 ± 0.35 A |
| V. vulnificus | 0.027 ± 0.08 D | - | 0.365 ± 0.21 B | 0.687 ± 0.22 C | 1.029 ± 0.31 A | 1.096 ± 0.28 A | |
| MPD (log) | V. parahaemolyticus | 9.48 ± 0.11 A | 9.66 ± 0.25 A | 9.12 ± 0.23 A.B | 8.86 ± 1.03 B | 9.74 ± 0.62 A | |
| V. vulnificus | 7.54 ± 1.34 C | - | 8.69 ± 0.77 B | 9.66 ± 0.29 A | 9.62 ± 0.32 A | 9.00 ± 0.85 A,B |
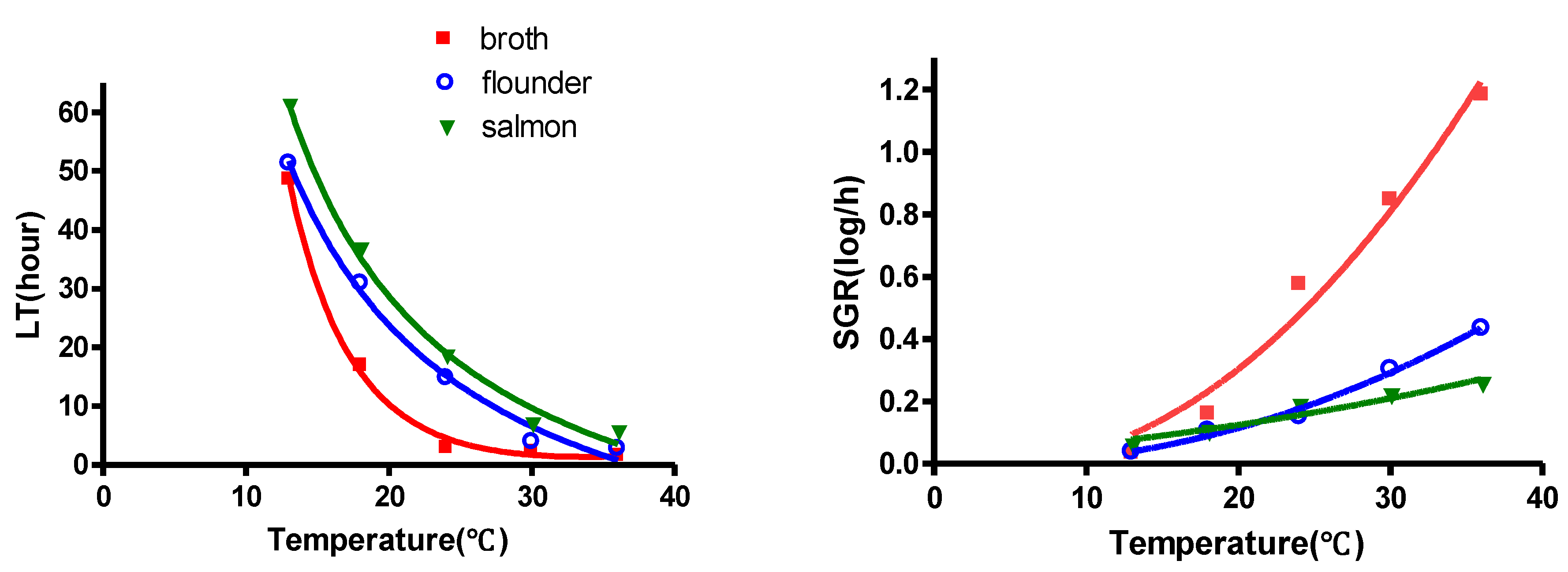
3.2. Comparison of Growth Kinetics of V. parahaemolyticus in Flounder and Salmon Sashimi
| Model | Parameter | Strain | R2 | Equation |
|---|---|---|---|---|
| Broth | LT | V. parahaemolyticus | 0.997 | Y* = 19.31 + (−1227/T) + (20945/T2) |
| V. vulnificus | 0.997 | Y = 2.784 + (−65.53/T) + (387.6/T2) | ||
| SGR | V. parahaemolyticus | 0.976 | Y = {0.03489(T−4.213)}2 | |
| V. vulnificus | 0.996 | Y = {0.03115(T + 1.316)}2 | ||
| Floundersashimi | LT | V. parahaemolyticus | 0.992 | Y † = −28.04 + (−1037/T) + (−1.867/T2) |
| SGR | V. parahaemolyticus | 0.990 | Y = {0.02017 (T−3.223)}2 | |
| Salmonsashimi | LT | V. parahaemolyticus | 0.993 | Y = −25.43 + (993.2/T) + (1792/T2) |
| SGR | V. parahaemolyticus | 0.939 | Y = {0.01052(T + 13.52)}2 | |
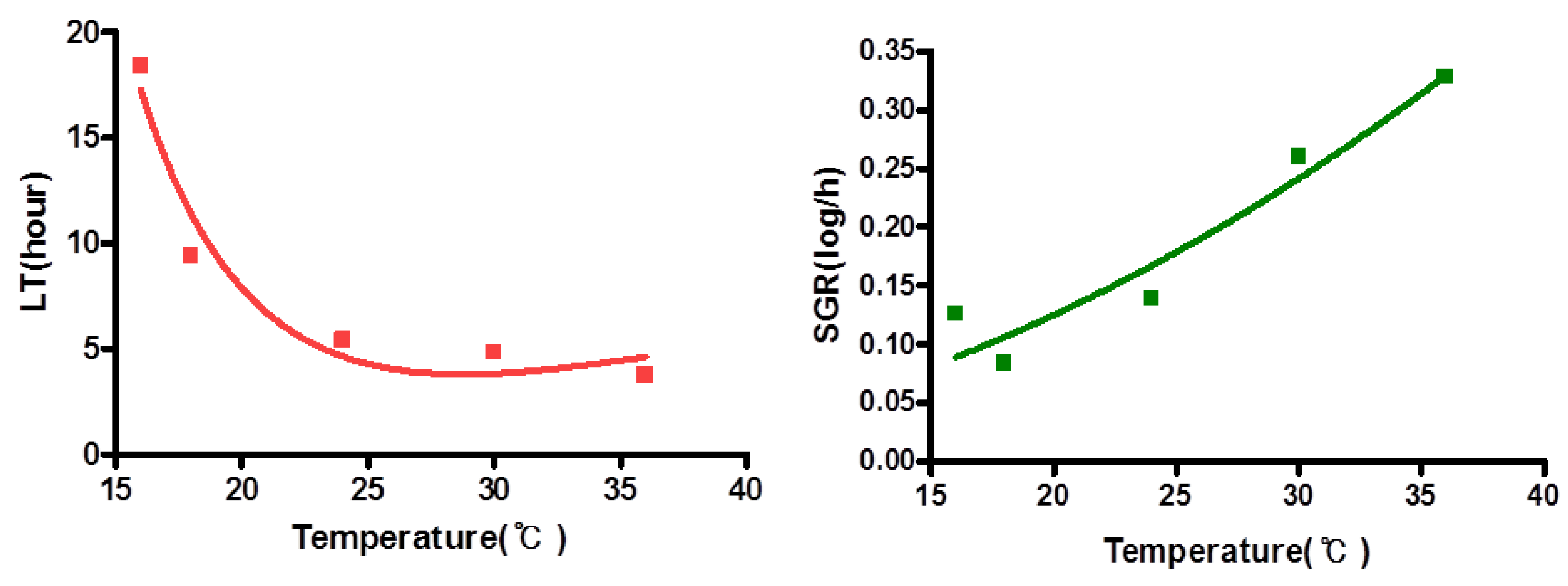
| oyster | LT | V. vulnificus | 0.950 | Y = 24.80 + (−1211/T) + (17449/T2) |
| SGR | V. vulnificus | 0.930 | Y = {0.01380 (T + 5.604)}2 |
| Sample | 13 °C | 18 °C | 24 °C | 30 °C | 36 °C | |
|---|---|---|---|---|---|---|
| LT(h) | Flouder | A 51.35 ± 1.78 a | A 30.96 ± 3.66 b | A 14.79 ± 2.99 c | A 3.90 ± 1.22 d | A 2.77 ± 1.07 d |
| Salmon | B 61.16 ± 3.11 a | B 36.81 ± 1.87 b | B 18.54 ± 3.03 c | B 6.98 ± 3.17 d | B 5.64 ± 2.12 d | |
| broth | C 48.64 ± 4.18 a | C 16.94 ± 5.15 b | C 2.52 ± 4.81 c | C 2.22 ± 1.18 c | C 1.61 ± 0.96 c,d | |
| SGR(log/h) | Flouder | A 0.037 ± 0.04 d | A 0.105 ± 0.09 c | A 0.152 ± 0.21 c | A 0.304 ± 0.02 b | A 0.435 ± 0.21 a |
| Salmon | A 0.059 ± 0.02 d | A 0.103 ± 0.12 c | A 0.185 ± 0.27 b | B 0.219 ± 0.76 a,b | B 0.256 ± 0.24 a | |
| broth | A 0.035 ± 0.03 e | B 0.160 ± 0.19 d | B 0.561 ± 0.35 c | C 0.847 ± 0.55 b | C 1.184 ± 0.59 a | |
| MPD(log) | Flouder | A 6.53 ± 0.67 b | A 7.30 ± 1.21 a | A 7.21 ± 1.09 a | A 7.60 ± 0.89 a | A 7.46 ± 0.45 a |
| Salmon | B 5.54 ± 1.65 c | B 6.51 ± 0.53 b | B 6.45 ± 0.65 b | A 7.34 ± 0.43 a | B 6.95 ± 0.66 b | |
| Broth | C 9.48 ± 0.87 a | C 9.66 ± 1.07 a | C 9.12 ± 1.19 a,b | B 8.86 ± 0.53 b | C 9.74 ± 0.32 a |

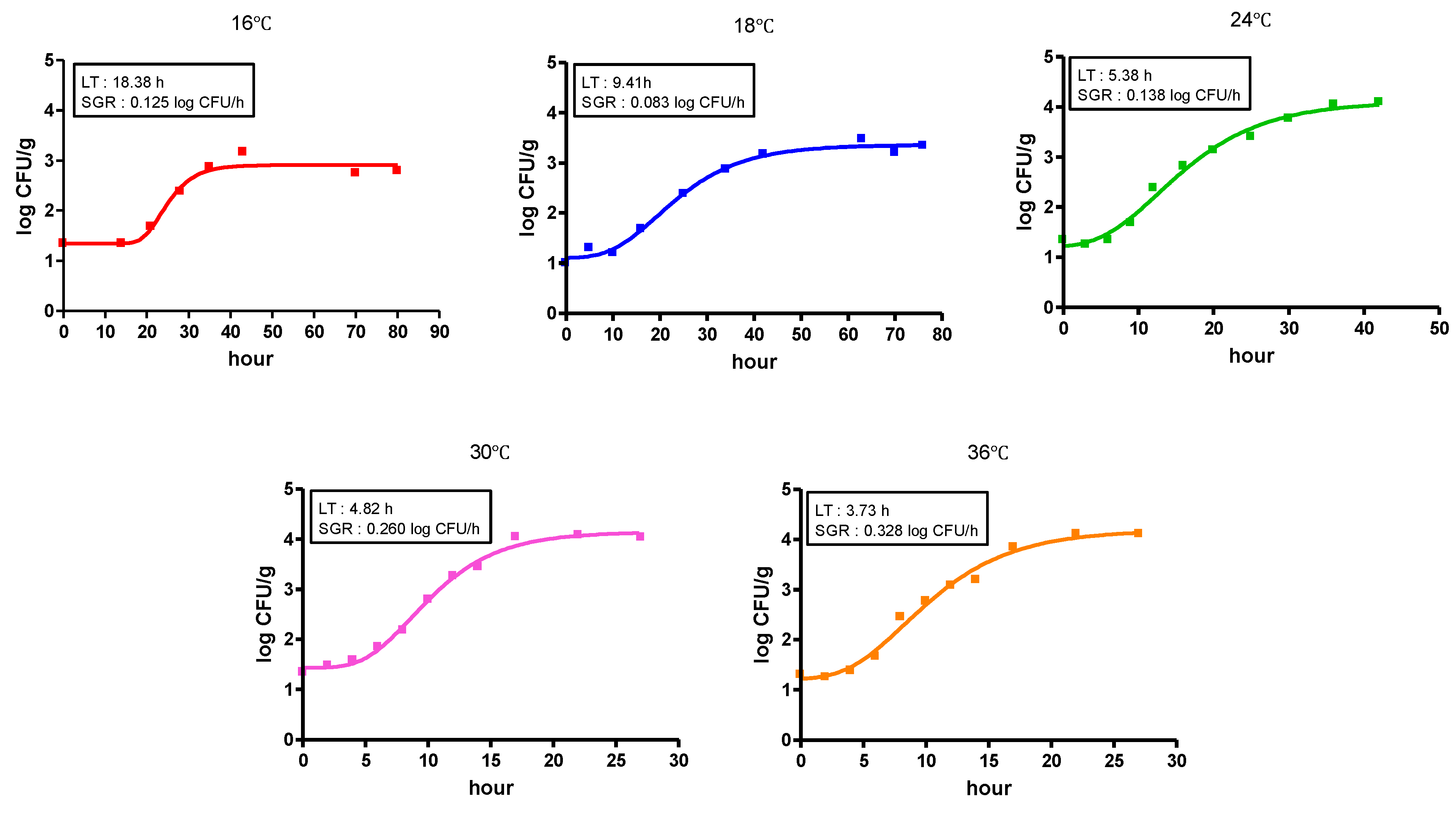
3.3. Growth Kinetics of Natural V. vulnificus in Shucked Oyster Meat


| Sample | 16 °C | 18 °C | 24 °C | 30 °C | 36 °C | |
|---|---|---|---|---|---|---|
| Broth | Initial density * | - | 2.44 ± 0.91 | 3.33 ± 0.44 | 3.32 ± 0.29 | 2.75 ± 0.45 |
| MPD | - | 8.70 ± 1.09 | 9.10 ± 0.45 | 8.90 ± 0.34 | 9.00 ± 0.89 | |
| Oyster | Initial density † | 1.34 ± 0.22 | 1.00 ± 0.32 | 1.31 ± 0.46 | 1.30 ± 0.23 | 1.34 ± 0.17 |
| MPD | 2.79 ± 0.58 | 3.47 ± 0.12 | 4.13 ± 0.54 | 4.08 ± 0.52 | 4.10 ± 0.36 |

4. Conclusions
Acknowledgments
Conflict of Interest
References
- Su, Y.C.; Liu, C. Vibrio parahaemolyticus: A concern of seafood safety. Food Microbiol. 2007, 24, 549–558. [Google Scholar] [CrossRef]
- Harth, E.; Matsuda, L.; Hernandez, C.; Rioseco, M.L.; Romero, J.; Gonzalez-Escalona, N.; Martinez-Urtaza, J.; Espejo, R.T. Epidemiology of Vibriop arahaemolyticus outbreaks, southern Chile. Emerg.Infect. Dis. 2009, 15, 163–168. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Preliminary Food Net Data on The Incidence of Infection with Pathogens Transmitted Commonly Through Food—10 States, 2009. Available online: www.cdc.gov/mmwr/preview/mmwrhtml/mm5914a2.htm (accessed on 8 May 2012).
- Chang, H.C.; Chen, M.L.; Su, Y.C.; Pai, J.Y.; Chiu, T.H. Molecular characterizations of pathogenic Vibrio parahaemolyticus isolated from Southern Taiwan oyster-growing environment. Food Control. 2011, 22, 245–251. [Google Scholar] [CrossRef]
- Kumamoto, K.S.; Vukich, D.J. Clinical infections of Vibriovulnificus: A case report and review of the literature. J. Emerg. Med. 1998, 16, 61–66. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Preliminary Food Net Data on The Incidence of Infection with Pathogens Transmitted Commonly Through Food—10 States, 2005. Available online: www.cdc.gov/mmwr/preview/mmwrhtml/mm5514a2.htm (accessed on 14 September 2011).
- Liu, F.; Guan, W.; Alam, M.J.; Shen, Z.; Zhang, S.; Li, L.; Shinoda, S.; Shi, L. Pulsed-field gel electrophoresis typing of multidrug-resistant vibrio parahaemolyticus isolated from various sources of seafood. J. Health Sci. 2009, 55, 783–789. [Google Scholar] [CrossRef]
- Korea Food and Drug Administration (KFDA). The Report of Foodborne Illness in Korea. Available online: www.kfda.go.kr (accessed on 11 December 2011. (in Korean).
- Ministry of Health & Welfare. Korea National Health and Nutrition Examination Survey (KNHNES). Available online: knhanes.cdc.go.kr (accessed on 5 July 2010). in Korean.
- Hamada-Sato, N.; Usui, K.; Kobayashi, T.; Imada, C.; Watanabe, E. Quality assurance of raw fish based on HACCP concept. Food Control 2005, 16, 301–307. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Vibrio Vulnificus Infections Associated with Raw Oyster Consumption-Florida, 1981-1992. Available online: www.cdc.gov/mmwr/preview/mmwrhtml/00020736.htm (accessed on 23 October 2011).
- Hlady, W.G. Vibrio Infections Associated with Raw Oyster Consumption in Florida, 1981–1994. J. Food Protect. 1997, 60, 353–357. [Google Scholar]
- Vasudevan, P.; Marek, P.; Daigle, S.; Hoagland, T.; Venkitanarayanan, K.S. Effect of chilling and freezing on survival of vibrio parahaemolyticuson fish fillets. J. Food Safety 2002, 22, 209–218. [Google Scholar] [CrossRef]
- Gooch, J.A.; DePaola, A.; Bowers, J.; Marshall, D.L. Growth and survival of vibrio parahaemolyticus in postharvest American oysters. J. Food Protect. 2002, 65, 970–974. [Google Scholar]
- Food and Drug Administration (FDA). Quantitative Risk Assessment on The Public Health Impact of Pathogenic Vibrio Parahaemolyticus Strains in Raw Oysters. Available online: www.fda.gov/Food/ScienceResearch/ResearchAreas/RiskAssessmentSafetyAssessment/ucm050421.htm (accessed on 19 July 2005).
- Burnham, V.E.; Janes, M.E.; Jakus, L.A.; Supan, J.; DePaola, A.; Bell, J. Growth and survival differences of Vibrio vulnificus and Vibrio parahaemolyticus strains during cold storage. J. Food Sci. 2009, 74, 314–318. [Google Scholar] [CrossRef]
- Yang, Z.Q.; Jiao, X.A.; Li, P.; Pan, Z.M.; Huang, J.L.; Gu, R.X.; Fang, W.M.; Chao, G.X. Predictive model of Vibrio parahaemolyticus growth and survival on salmon meat as a function of temperature. Food Microbiol. 2009, 26, 606–614. [Google Scholar] [CrossRef]
- Miles, D.W.; Ross, T.; Olley, J.; McMeekin, T.A. Development and evaluation of a predictive model for the effect of temperature and water activity on the growth rate of Vibrio parahaemolyticus. Int. J. Food Microbiol. 1997, 38, 133–142. [Google Scholar] [CrossRef]
- Yoon, K.S.; Min, K.J.; Jung, Y.J.; Kwon, K.Y.; Lee, J.K.; Oh, S.W. A model of the effect of temperature on the growth of pathogenic and nonpathogenic Vibrio parahaemolyticus isolated from oysters in Korea. Food Microbiol. 2008, 25, 635–641. [Google Scholar] [CrossRef]
- Fernandez-Piquer, J.; Bowman, J.P.; Ross, T.; Tamplin, M.L. Predictive models for the effect of storage temperature on vibrio parahaemolyticus viability and counts of total viable bacteria in pacific oysters (crassostrea gigas). Appl. Environ. Microb. 2011, 77, 8687–8695. [Google Scholar] [CrossRef]
- DaSilva, L.; Parveen, S.; DePaola, A.; Bowers, J.; Brohawn, K.; Tamplin, M.L. Development and validation of a predictive model for the growth of vibrio vulnificus in postharvest shellstock oysters. Appl. Environ. Microb. 2012, 78, 1675–1681. [Google Scholar] [CrossRef]
- Lee, J.K.; Jung, D.W.; Eom, S.Y.; Oh, S.W.; Kim, Y.; Kwak, H.S.; Kim, Y.H. Occurrence of Vibrio parahaemolyticus in oysters from Korean retail outlets. Food Control. 2008, 19, 990–994. [Google Scholar] [CrossRef]
- Davey, K.R. A predictive model for combined temperature and water activity on microbial growth during the growth phase. J. Appl. Microbiol. 1989, 67, 483–488. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Olley, J.; McMeekin, T.A.; Ball, A. Relationship between temperature and growth rate of bacterial cultures. J. Bacteriol. 1982, 149, 1–5. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, Y.W.; Lee, S.H.; Hwang, I.G.; Yoon, K.S. Effect of Temperature on Growth of Vibrio paraphemolyticus and Vibrio vulnificus in Flounder, Salmon Sashimi and Oyster Meat. Int. J. Environ. Res. Public Health 2012, 9, 4662-4675. https://doi.org/10.3390/ijerph9124662
Kim YW, Lee SH, Hwang IG, Yoon KS. Effect of Temperature on Growth of Vibrio paraphemolyticus and Vibrio vulnificus in Flounder, Salmon Sashimi and Oyster Meat. International Journal of Environmental Research and Public Health. 2012; 9(12):4662-4675. https://doi.org/10.3390/ijerph9124662
Chicago/Turabian StyleKim, Yoo Won, Soon Ho Lee, In Gun Hwang, and Ki Sun Yoon. 2012. "Effect of Temperature on Growth of Vibrio paraphemolyticus and Vibrio vulnificus in Flounder, Salmon Sashimi and Oyster Meat" International Journal of Environmental Research and Public Health 9, no. 12: 4662-4675. https://doi.org/10.3390/ijerph9124662
APA StyleKim, Y. W., Lee, S. H., Hwang, I. G., & Yoon, K. S. (2012). Effect of Temperature on Growth of Vibrio paraphemolyticus and Vibrio vulnificus in Flounder, Salmon Sashimi and Oyster Meat. International Journal of Environmental Research and Public Health, 9(12), 4662-4675. https://doi.org/10.3390/ijerph9124662