Community Structure Analyses of Anodic Biofilms in a Bioelectrochemical System Combined with an Aerobic Reactor

Abstract
:1. Introduction
2. Materials and Methods
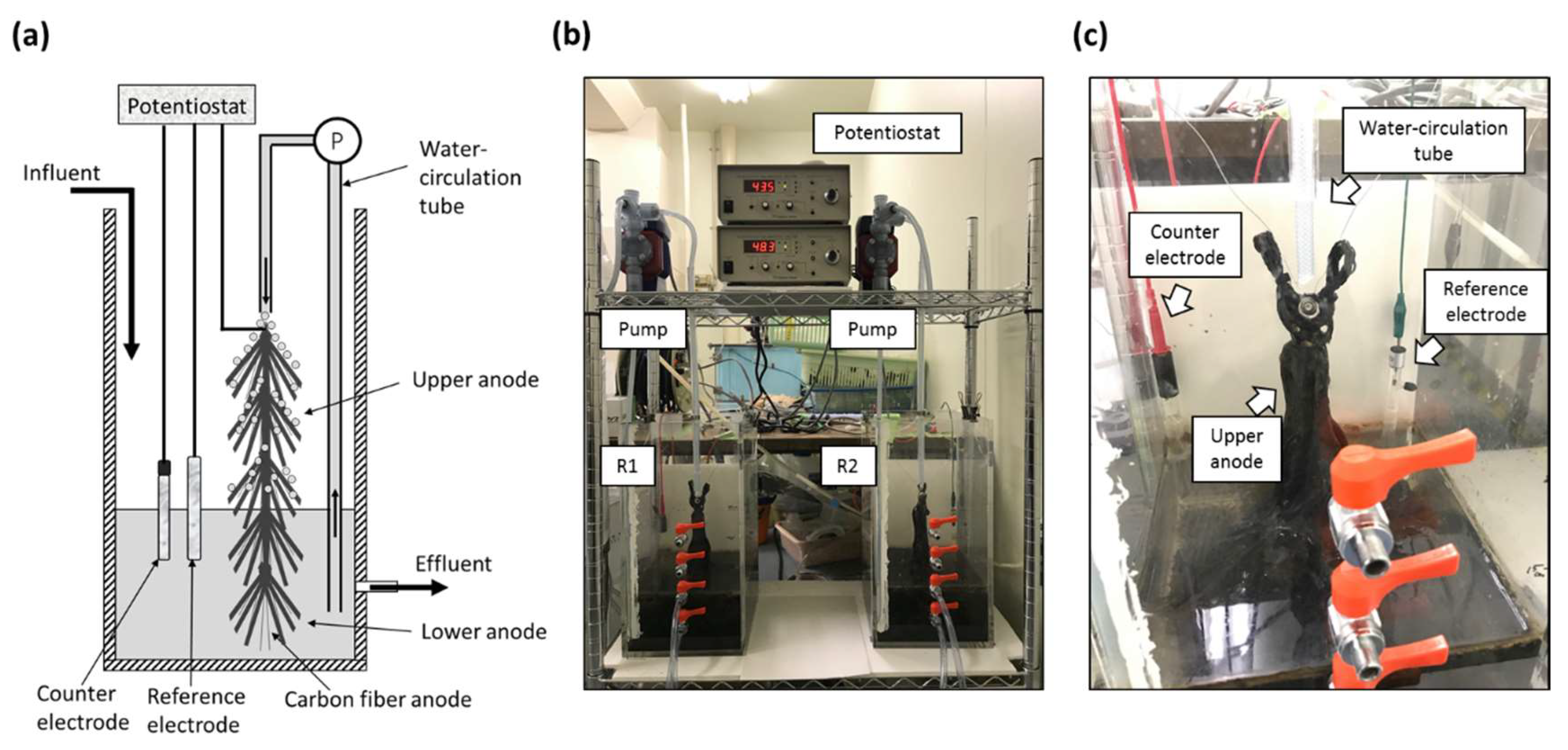
2.1. Reactor Construction and Operation
2.2. Microbial Community Analysis
3. Results and Discussion
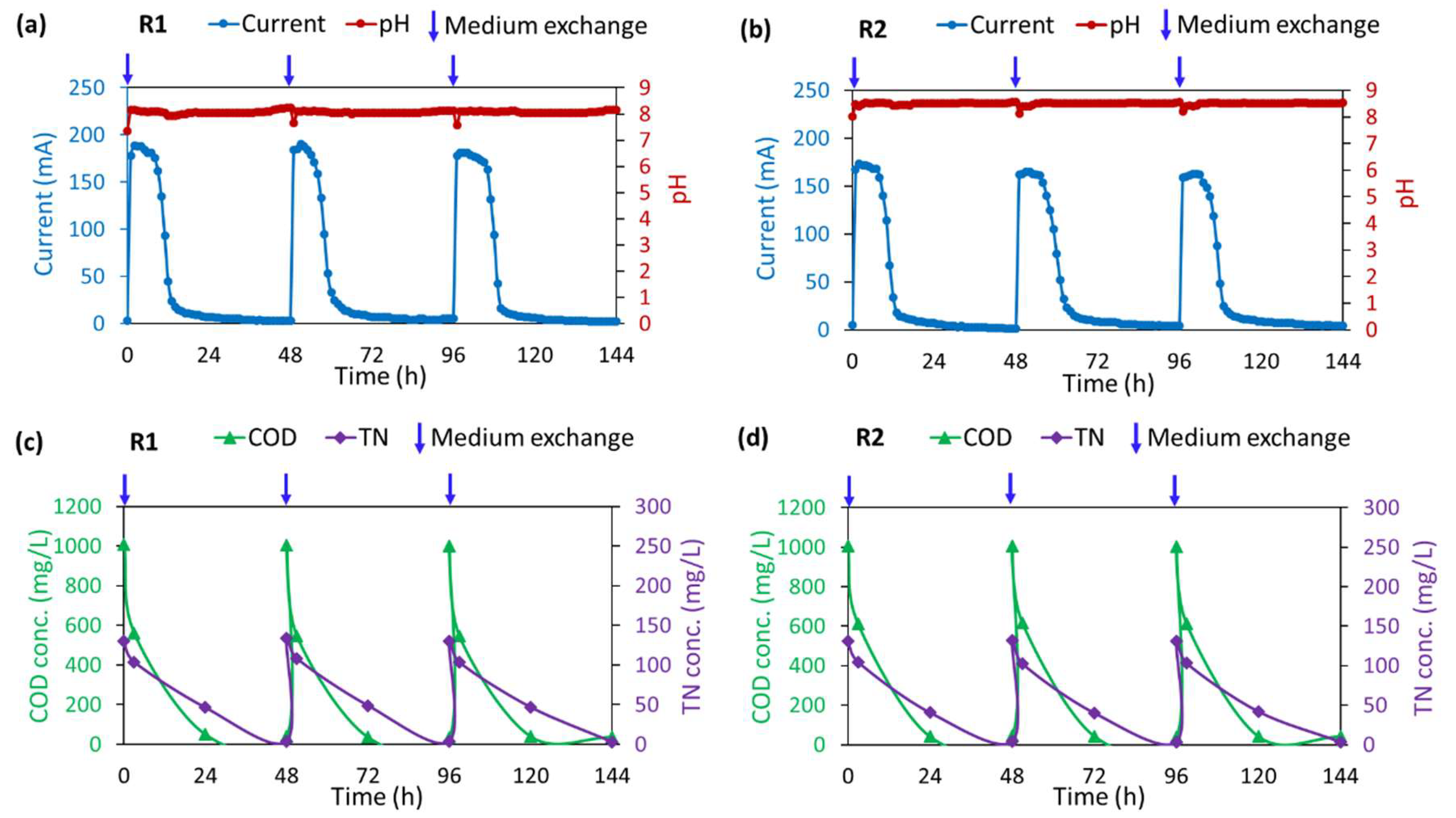
3.1. Decomposition of Organic Matter and Soluble Nitrogen Compounds
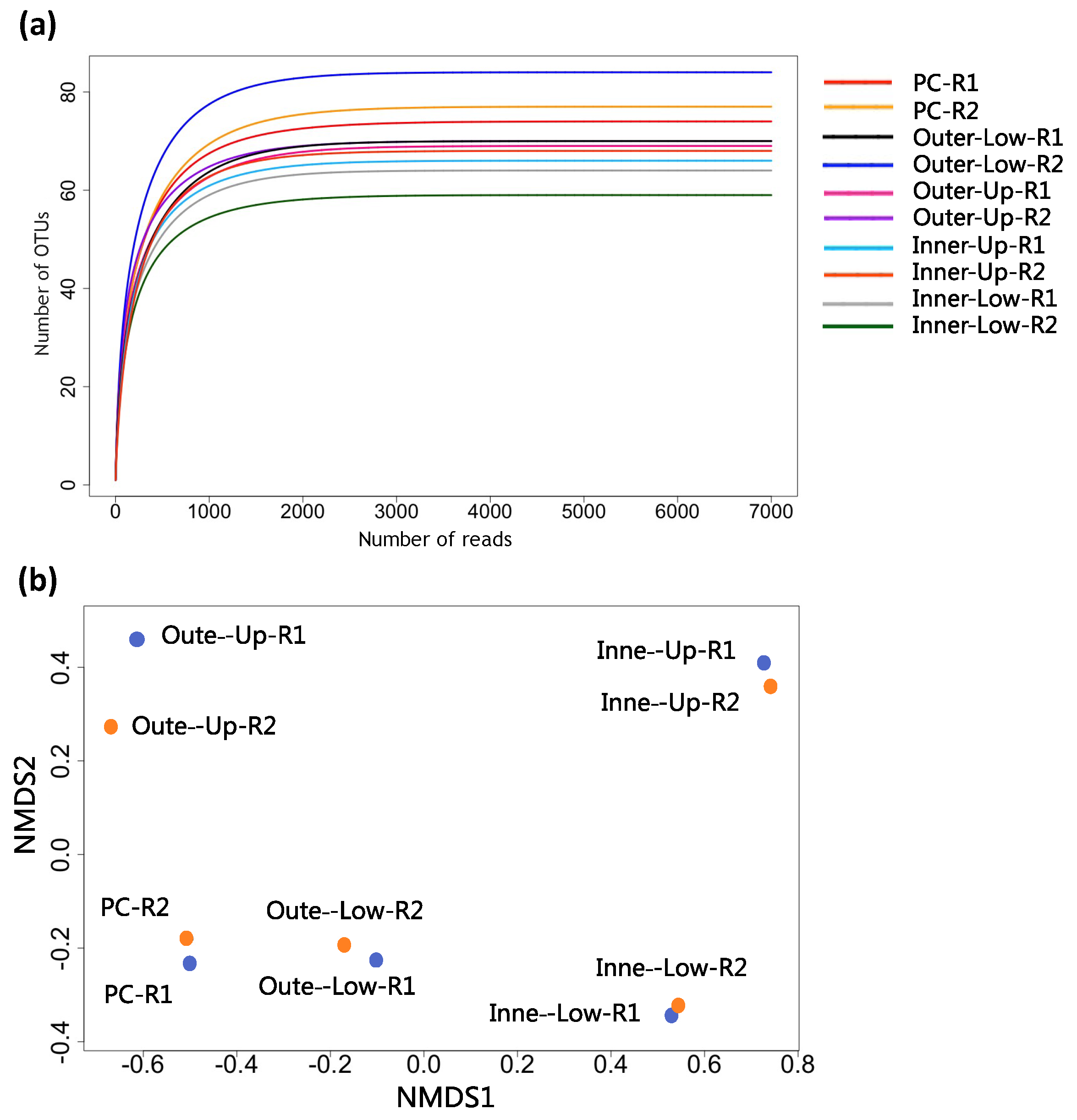
3.2. Analyses of Alpha and Beta Diversities
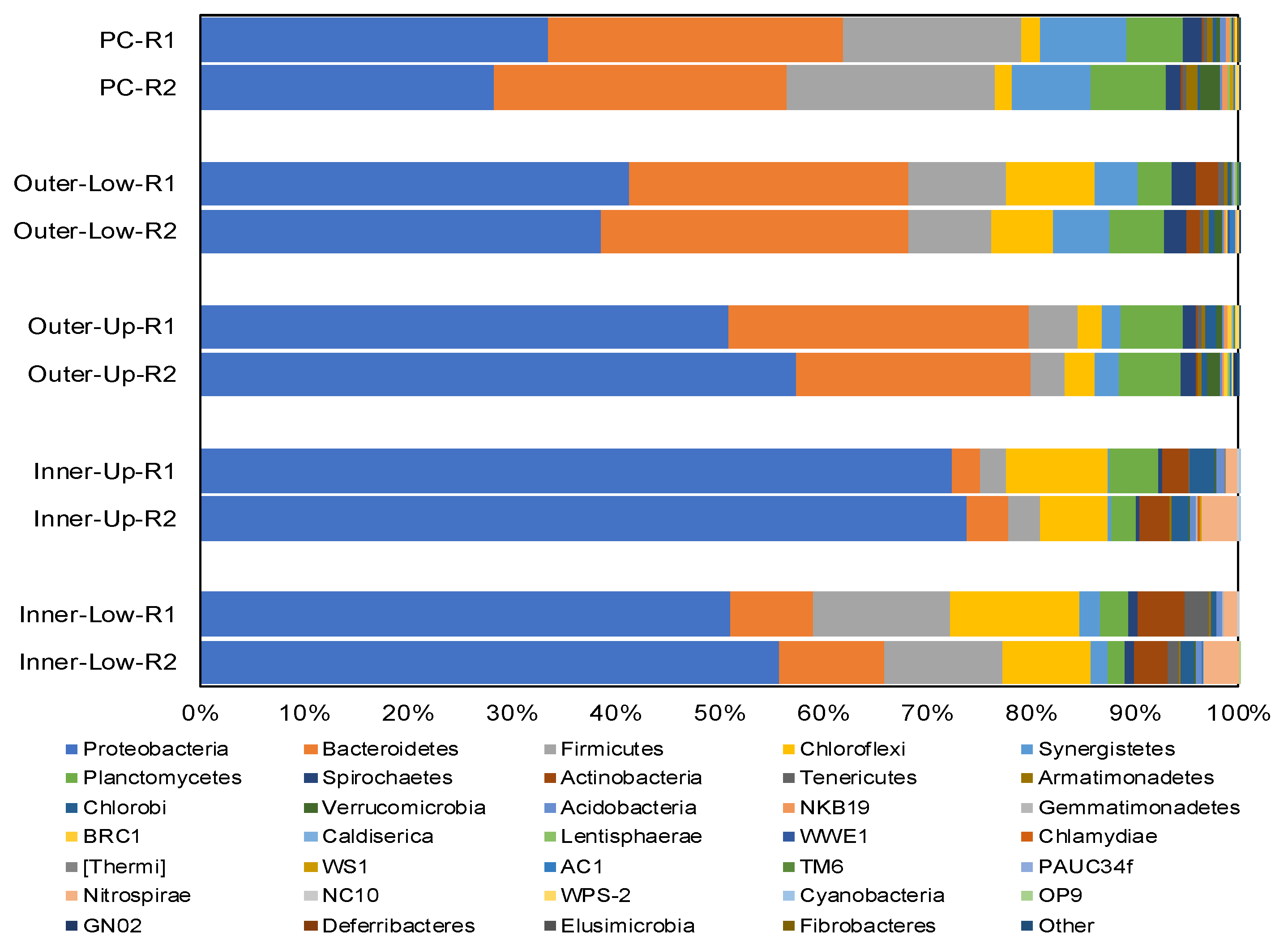
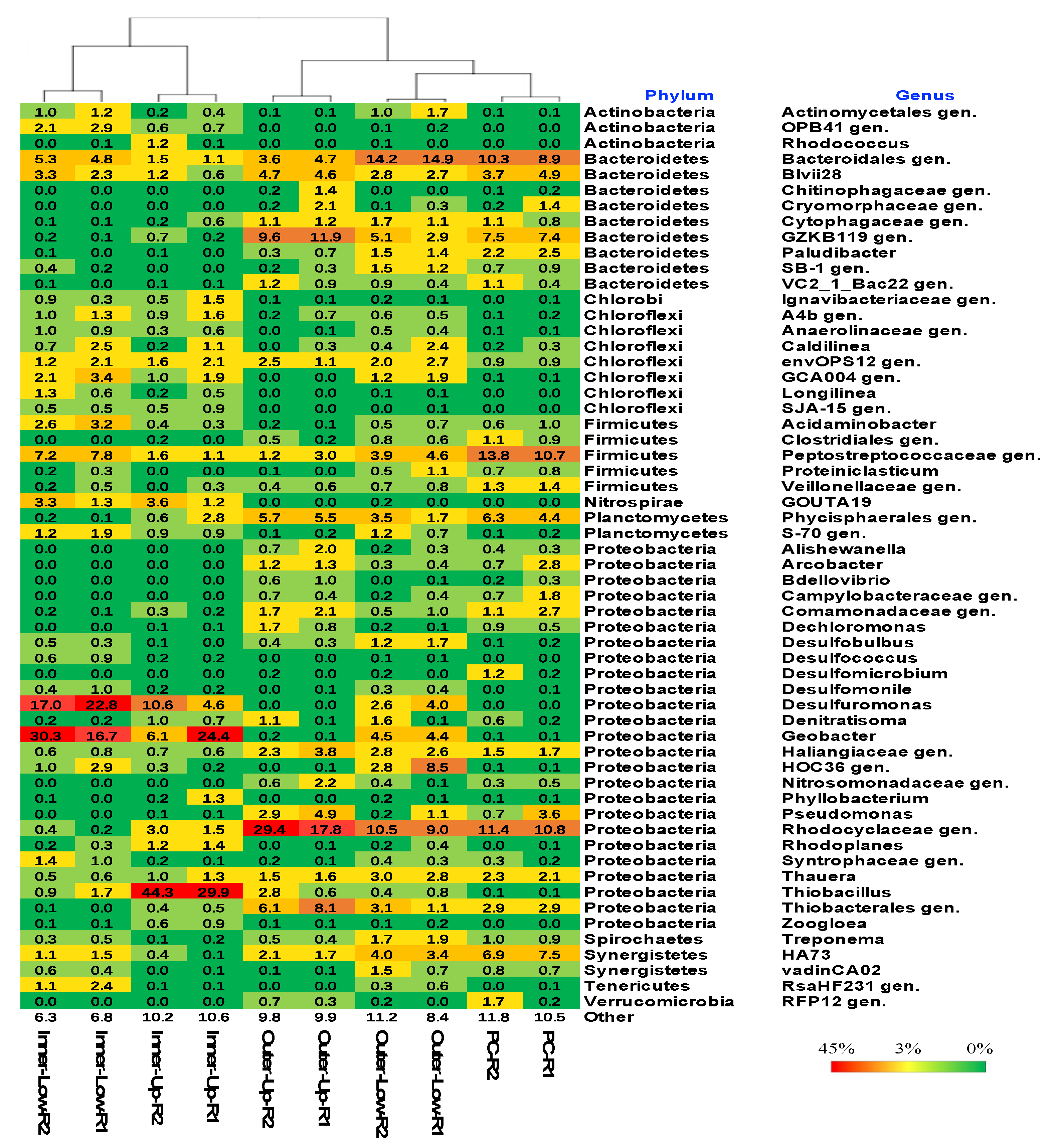
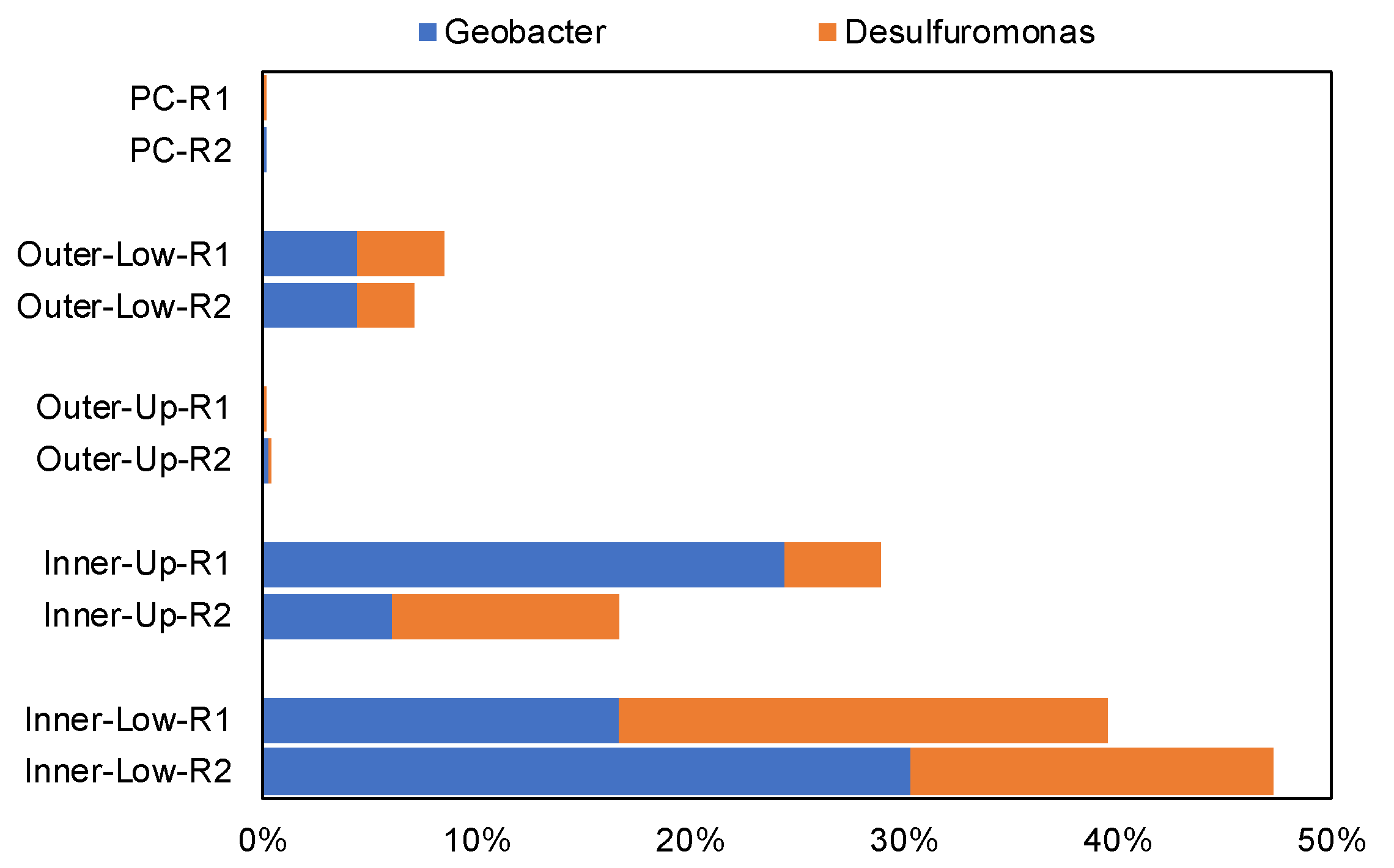
3.3. Structures of the Microbial Community in the Anodic Biofilms of BES-TF Reactors at the Phylum and Genus Levels
3.4. Application of BES Technology to Aerobic Bioreactors
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, H.; Ramnarayanan, R.; Logan, B.E. Production of Electricity during Wastewater Treatment Using a Single Chamber Microbial Fuel Cell. Environ. Sci. Technol. 2004, 38, 2281–2285. [Google Scholar] [CrossRef] [PubMed]
- Call, D.; Logan, B.E. Hydrogen Production in a Single Chamber Microbial Electrolysis Cell Lacking a Membrane. Environ. Sci. Technol. 2008, 42, 3401–3406. [Google Scholar] [CrossRef] [PubMed]
- Sleutels, T.H.J.A.; Ter Heijne, A.; Buisman, C.J.N.; Hamelers, H.V.M. Bioelectrochemical systems: An outlook for practical applications. ChemSusChem 2012, 5, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Luo, H.; Fallgren, P.H.; Jin, S.; Ren, Z.J. Bioelectrochemical system platform for sustainable environmental remediation and energy generation. Biotechnol. Adv. 2015, 33, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yu, W.; Graham, N.; Zhao, Y.; Qu, J. Application of Integrated Bioelectrochemical-Wetland Systems for Future Sustainable Wastewater Treatment. Environ. Sci. Technol. 2019, 53, 1741–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, B.E. Exoelectrogenic bacteria that power microbial fuel cells. Nat. Rev. Microbiol. 2009, 7, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R. Electromicrobiology. Annu. Rev. Microbiol. 2012, 66, 391–409. [Google Scholar] [CrossRef]
- Wang, H.; Ren, Z.J. A comprehensive review of microbial electrochemical systems as a platform technology. Biotechnol. Adv. 2013, 31, 1796–1807. [Google Scholar] [CrossRef]
- Yamashita, T.; Ookawa, N.; Ishida, M.; Kanamori, H.; Sasaki, H.; Katayose, Y.; Yokoyama, H. A novel open-type biosensor for the in-situ monitoring of biochemical oxygen demand in an aerobic environment. Sci. Rep. 2016, 6, 38552. [Google Scholar] [CrossRef]
- Clauwaert, P.; Aelterman, P.; Pham, T.H.; De Schamphelaire, L.; Carballa, M.; Rabaey, K.; Verstraete, W. Minimizing losses in bio-electrochemical systems: The road to applications. Appl. Microbiol. Biotechnol. 2008, 79, 901–913. [Google Scholar] [CrossRef]
- Lovley, D.R.; Giovannoni, S.J.; White, D.C.; Champine, J.E.; Phillips, E.J.P.; Gorby, Y.A.; Goodwin, S. Geobacter metallireducens gen. nov. sp. nov., a microorganism capable of coupling the complete oxidation of organic compounds to the reduction of iron and other metals. Arch. Microbiol. 1993, 159, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, S.; Cord-Ruwisch, R.; Schink, B. A periplasmic and extracellular c-type cytochrome of Geobacter sulfurreducens acts as a ferric iron reductase and as an electron carrier to other acceptors or to partner bacteria. J. Bacteriol. 1998, 180, 3686–3691. [Google Scholar] [PubMed]
- Reguera, G.; McCarthy, K.D.; Mehta, T.; Nicoll, J.S.; Tuominen, M.T.; Lovley, D.R. Extracellular electron transfer via microbial nanowires. Nature 2005, 435, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.-J.; Choi, M.-J.; Lee, J.-W.; Kim, K.-Y.; Kim, I.S. Effect of different substrates on the performance, bacterial diversity, and bacterial viability in microbial fuel cells. Bioresour. Technol. 2009, 100, 3518–3525. [Google Scholar] [CrossRef] [PubMed]
- Logan, B.E.; Wallack, M.J.; Kim, K.-Y.; He, W.; Feng, Y.; Saikaly, P.E. Assessment of Microbial Fuel Cell Configurations and Power Densities. Environ. Sci. Technol. Lett. 2015, 2, 206–214. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Liu, L.; Yang, F. Development of a novel proton exchange membrane-free integrated MFC system with electric membrane bioreactor and air contact oxidation bed for efficient and energy-saving wastewater treatment. Bioresour. Technol. 2017, 238, 472–483. [Google Scholar] [CrossRef]
- Ren, L.; Ahn, Y.; Logan, B.E. A Two-Stage Microbial Fuel Cell and Anaerobic Fluidized Bed Membrane Bioreactor (MFC-AFMBR) System for Effective Domestic Wastewater Treatment. Environ. Sci. Technol. 2014, 48, 4199–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Zhu, X.; Ni, J.; Borthwick, A. Palm oil mill effluent treatment using a two-stage microbial fuel cells system integrated with immobilized biological aerated filters. Bioresour. Technol. 2010, 101, 2729–2734. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Feng, Y.; Qu, Y.; Du, Y.; Zhou, X.; Liu, J. A combined system of microbial fuel cell and intermittently aerated biological filter for energy self-sufficient wastewater treatment. Sci. Rep. 2015, 5, 18070. [Google Scholar] [CrossRef] [PubMed]
- Wik, T. Trickling Filters and Biofilm Reactor Modelling. Rev. Environ. Sci. Biotechnol. 2003, 2, 193–212. [Google Scholar] [CrossRef]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial Fuel Cells: Methodology and Technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- Hou, D.; Lu, L.; Ren, Z.J. Microbial fuel cells and osmotic membrane bioreactors have mutual benefits for wastewater treatment and energy production. Water Res. 2016, 98, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 September 2019).
- Yokoyama, H.; Waki, M.; Moriya, N.; Yasuda, T.; Tanaka, Y.; Haga, K. Effect of fermentation temperature on hydrogen production from cow waste slurry by using anaerobic microflora within the slurry. Appl. Microbiol. Biotechnol. 2007, 74, 474–483. [Google Scholar] [CrossRef]
- Yokoyama, H.; Ohmori, H.; Ishida, M.; Waki, M.; Tanaka, Y. Treatment of cow-waste slurry by a microbial fuel cell and the properties of the treated slurry as a liquid manure. Anim. Sci. J. 2006, 77, 634–638. [Google Scholar] [CrossRef]
- Mulder, A.; van de Graaf, A.A.; Robertson, L.A.; Kuenen, J.G. Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed reactor. FEMS Microbiol. Ecol. 1995, 16, 177–183. [Google Scholar] [CrossRef]
- Yamashita, T.; Ishida, M.; Asakawa, S.; Kanamori, H.; Sasaki, H.; Ogino, A.; Katayose, Y.; Hatta, T.; Yokoyama, H. Enhanced electrical power generation using flame-oxidized stainless steel anode in microbial fuel cells and the anodic community structure. Biotechnol. Biofuels 2016, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Pfennig, N.; Biebl, H. Desulfuromonas acetoxidans gen. nov. and sp. nov., a new anaerobic, sulfur-reducing, acetate-oxidizing bacterium. Arch. Microbiol. 1976, 110, 3–12. [Google Scholar] [CrossRef]
- Roden, E.E.; Lovley, D.R. Dissimilatory Fe(III) Reduction by the Marine Microorganism Desulfuromonas acetoxidans. Appl. Environ. Microbiol. 1993, 59, 734–742. [Google Scholar]
- An, T.T.; Picardal, F.W. Desulfuromonas carbonis sp. nov., an Fe(III)-, S0- and Mn(IV)-reducing bacterium isolated from an active coalbed methane gas well. Int. J. Syst. Evol. Microbiol. 2015, 65, 1686–1693. [Google Scholar] [CrossRef]
- Finster, K.; Bak, F.; Pfennig, N. Desulfuromonas acetexigens sp. nov., a dissimilatory sulfur-reducing eubacterium from anoxic freshwater sediments. Arch. Microbiol. 1994, 161, 328–332. [Google Scholar] [CrossRef]
- Yamashita, T.; Yokoyama, H. Molybdenum anode: A novel electrode for enhanced power generation in microbial fuel cells, identified via extensive screening of metal electrodes. Biotechnol. Biofuels 2018, 11, 39. [Google Scholar] [CrossRef]
- Eyiuche, N.J.; Asakawa, S.; Yamashita, T.; Ikeguchi, A.; Kitamura, Y.; Yokoyama, H. Community analysis of biofilms on flame-oxidized stainless steel anodes in microbial fuel cells fed with different substrates. BMC Microbiol. 2017, 17, 145. [Google Scholar] [CrossRef]
- Sun, G.; Thygesen, A.; Meyer, A.S. Acetate is a superior substrate for microbial fuel cell initiation preceding bioethanol effluent utilization. Appl. Microbiol. Biotechnol. 2015, 99, 4905–4915. [Google Scholar] [CrossRef]
- Bond, D.R.; Holmes, D.E.; Tender, L.M.; Lovley, D.R. Electrode-reducing microorganisms that harvest energy from marine sediments. Science 2002, 295, 483–485. [Google Scholar] [CrossRef]
- Oren, A. The Family Rhodocyclaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 975–998. ISBN 978-3-642-30197-1. [Google Scholar]
- Singleton, D.R.; Dickey, A.N.; Scholl, E.H.; Wright, F.A.; Aitken, M.D. Complete Genome Sequence of a Novel Bacterium within the Family Rhodocyclaceae That Degrades Polycyclic Aromatic Hydrocarbons. Genome Announc. 2015, 3, e00251-15. [Google Scholar] [CrossRef]
- Zhang, G.; Zhao, Q.; Jiao, Y.; Wang, K.; Lee, D.-J.; Ren, N. Efficient electricity generation from sewage sludge using biocathode microbial fuel cell. Water Res. 2012, 46, 43–52. [Google Scholar] [CrossRef]
- Song, Y.; Xiao, L.; Jayamani, I.; He, Z.; Cupples, A.M. A novel method to characterize bacterial communities affected by carbon source and electricity generation in microbial fuel cells using stable isotope probing and Illumina sequencing. J. Microbiol. Methods 2015, 108, 4–11. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, J.; Xu, Y.; Yu, H.; Wu, Z. Power production from different types of sewage sludge using microbial fuel cells: A comparative study with energetic and microbiological perspectives. J. Power Sources 2013, 235, 280–288. [Google Scholar] [CrossRef]
- Baalsrud, K.; Baalsrud, K.S. Studies on thiobacillus denitrificans. Arch. Mikrobiol. 1954, 20, 34–62. [Google Scholar] [CrossRef]
- Aminuddin, M.; Nicholas, D.J.D. Sulphide oxidation linked to the reduction of nitrate and nitrite in Thiobacillus denitrificans. Biochim. Biophys. Acta BBA Bioenerg. 1973, 325, 81–93. [Google Scholar] [CrossRef]
- Müller, J.B.; Ramos, D.T.; Larose, C.; Fernandes, M.; Lazzarin, H.S.C.; Vogel, T.M.; Corseuil, H.X. Combined iron and sulfate reduction biostimulation as a novel approach to enhance BTEX and PAH source-zone biodegradation in biodiesel blend-contaminated groundwater. J. Hazard. Mater. 2017, 326, 229–236. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Performance Indicators | R1 | R2 |
|---|---|---|
| Coulombic efficiency (%) at 24 h | 16.8 ± 0.7 | 14.1 ± 0.4 |
| COD decomposition efficiency (%) at 48 h | 99.7 ± 0.03 | 96.2 ± 0.6 |
| TN decomposition efficiency (%) at 48 h | 97.5 ± 0.1 | 97.2 ± 0.4 |
| Samples | No. of Reads | No. of OTUs | Shannon Index | Simpson Index |
|---|---|---|---|---|
| PC-R1 | 16449 | 189 | 3.342 | 0.943 |
| PC-R2 | 18835 | 190 | 3.278 | 0.934 |
| Outer-Low-R1 | 10280 | 171 | 3.428 | 0.944 |
| Outer-Low-R2 | 17104 | 201 | 3.548 | 0.948 |
| Outer-Up-R1 | 8754 | 157 | 3.215 | 0.928 |
| Outer-Up-R2 | 18199 | 183 | 2.978 | 0.881 |
| Inner-Up-R1 | 7424 | 151 | 2.695 | 0.834 |
| Inner-Up-R2 | 17343 | 203 | 2.512 | 0.771 |
| Inner-Low-R1 | 8515 | 134 | 3.007 | 0.9 |
| Inner-Low-R2 | 20362 | 168 | 2.725 | 0.856 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Q.; Yamashita, T.; Matsuura, N.; Yamamoto-Ikemoto, R.; Yokoyama, H. Community Structure Analyses of Anodic Biofilms in a Bioelectrochemical System Combined with an Aerobic Reactor. Energies 2019, 12, 3643. https://doi.org/10.3390/en12193643
Liang Q, Yamashita T, Matsuura N, Yamamoto-Ikemoto R, Yokoyama H. Community Structure Analyses of Anodic Biofilms in a Bioelectrochemical System Combined with an Aerobic Reactor. Energies. 2019; 12(19):3643. https://doi.org/10.3390/en12193643
Chicago/Turabian StyleLiang, Qiaochu, Takahiro Yamashita, Norihisa Matsuura, Ryoko Yamamoto-Ikemoto, and Hiroshi Yokoyama. 2019. "Community Structure Analyses of Anodic Biofilms in a Bioelectrochemical System Combined with an Aerobic Reactor" Energies 12, no. 19: 3643. https://doi.org/10.3390/en12193643
APA StyleLiang, Q., Yamashita, T., Matsuura, N., Yamamoto-Ikemoto, R., & Yokoyama, H. (2019). Community Structure Analyses of Anodic Biofilms in a Bioelectrochemical System Combined with an Aerobic Reactor. Energies, 12(19), 3643. https://doi.org/10.3390/en12193643