Bioinspired Materials: From Living Systems to New Concepts in Materials Chemistry


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. From Creatures to Concepts
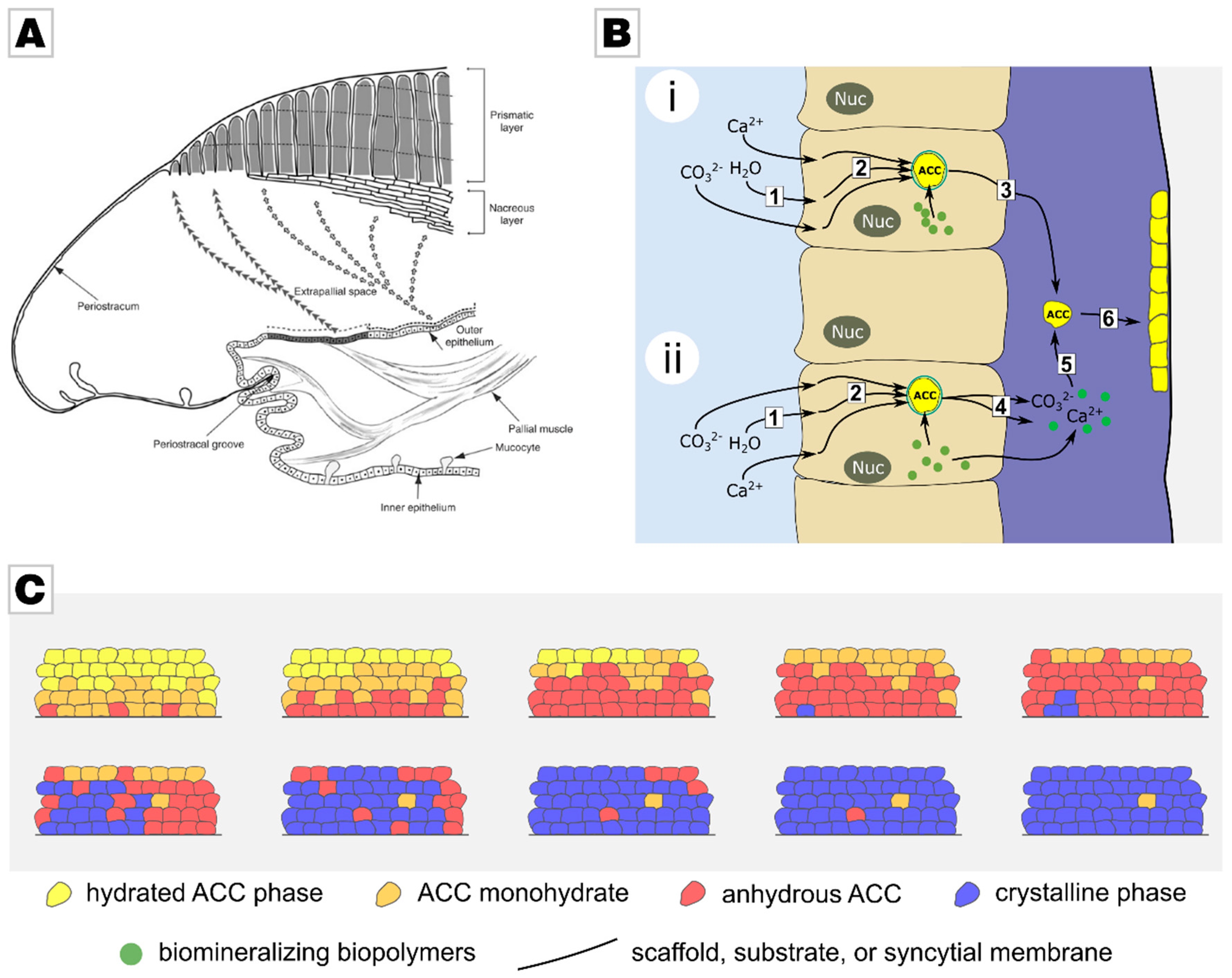
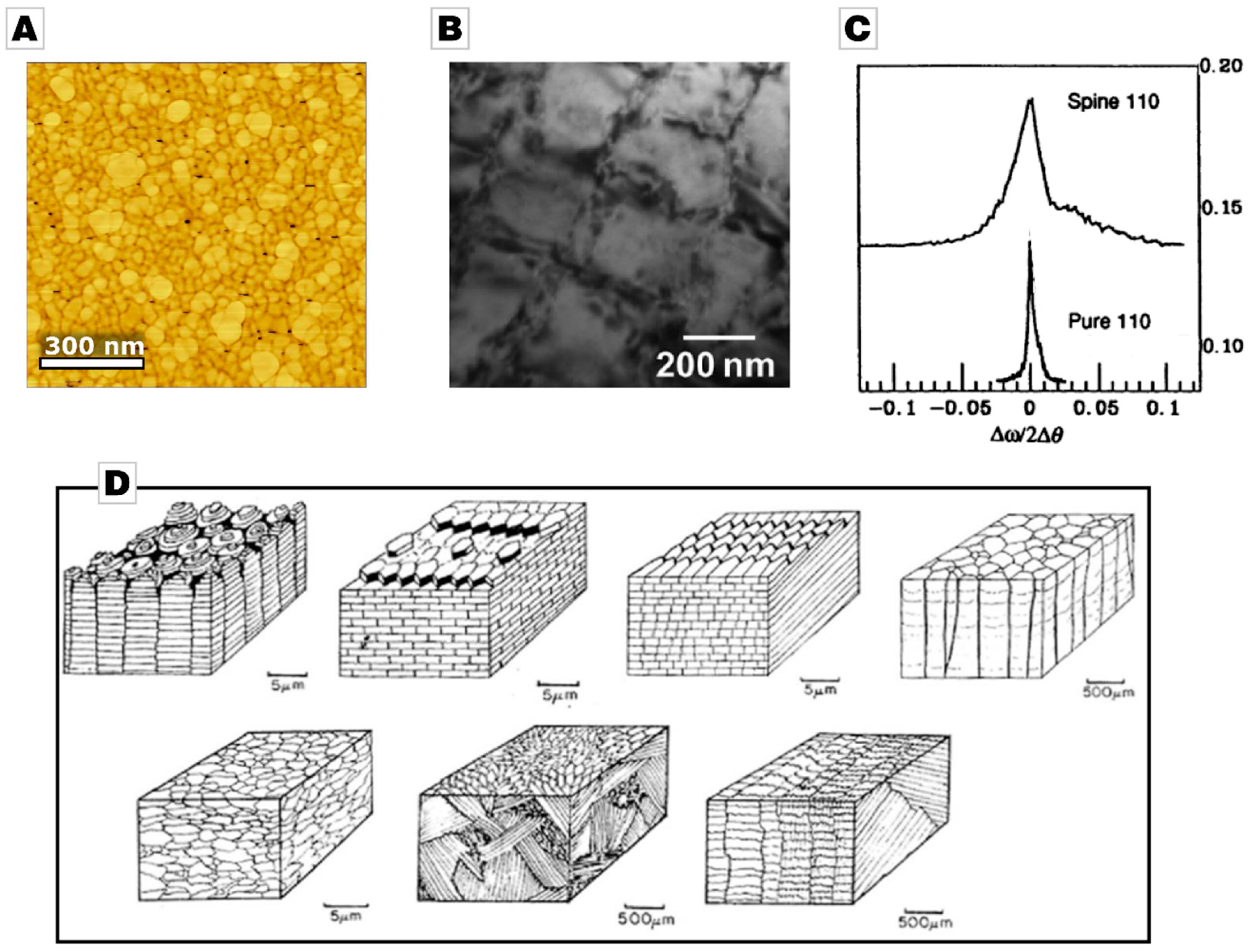
2.1. From Calcareous Creatures to New Concepts in Crystallization
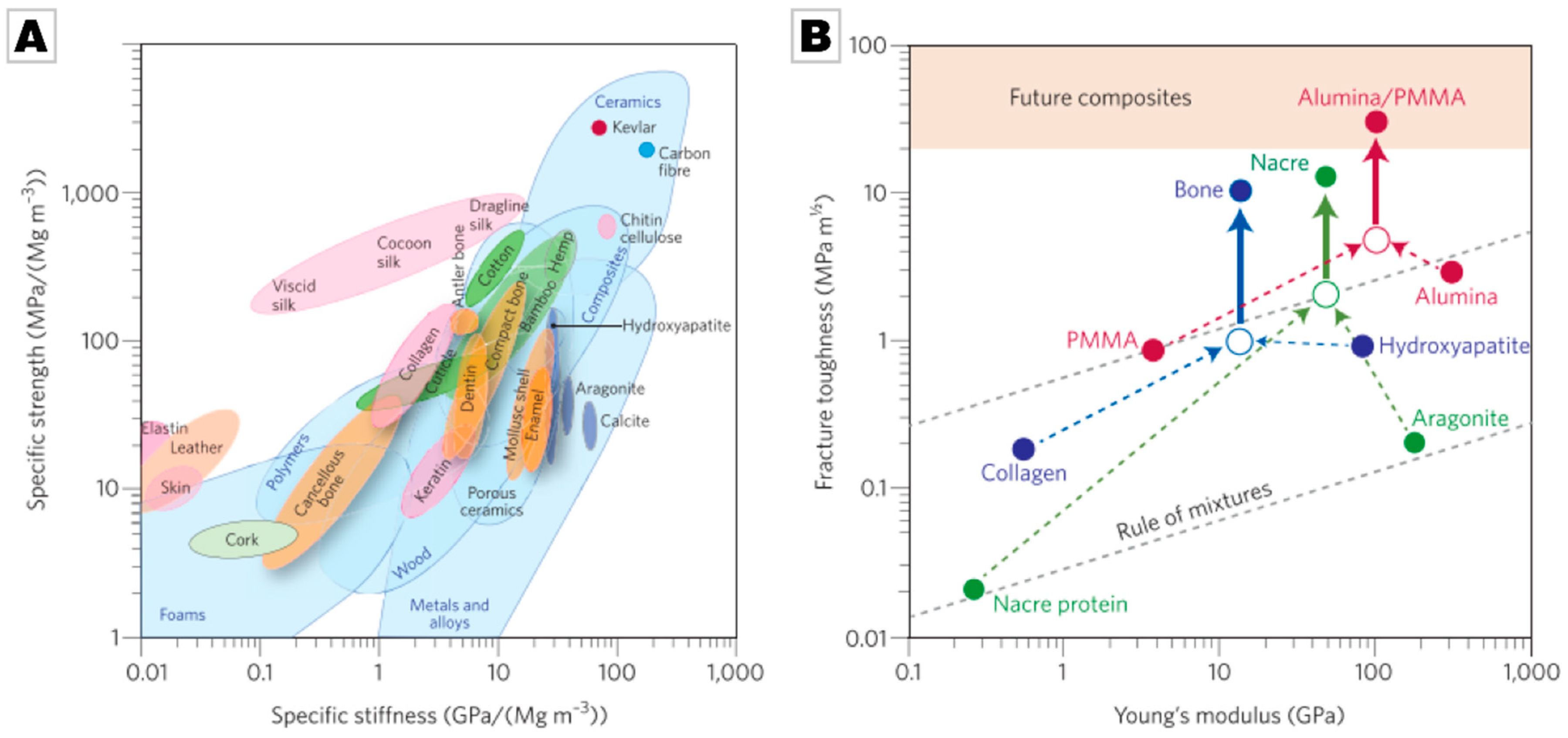
2.2. Biominerals as an Evolutionarily-Tested Archive of Functional Material Design Motifs
2.3. Biogenic Silica Formation Demonstrates How to Mildly Drive Metal Oxide Formation
3. Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Mann, S. Biomineralization: Principles and Concepts in Bioinorganic Materials Chemistry; Compton, R., Davies, S.G., Evans, J., Eds.; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Meldrum, F.C.; Cölfen, H.; Cölfen, H. Controlling Mineral Morphologies and Structures in Biological and Synthetic Systems. Chem. Rev. 2008, 108, 4332–4432. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, H.D.; Rim, J.E.; Barthelat, F.F.; Buehler, M.J. Merger of Structure and Material in Nacre and Bone—Perspectives on de Novo Biomimetic Materials. Prog. Mater. Sci. 2009, 54, 1059–1100. [Google Scholar] [CrossRef]
- Barthelat, F.; Rabiei, R. Toughness Amplification in Natural Composites. J. Mech. Phys. Solids 2011, 59, 829–840. [Google Scholar] [CrossRef]
- Wegst, U.G.K.; Bai, H.; Saiz, E.; Tomsia, A.P.; Ritchie, R.O. Bioinspired Structural Materials. Nat. Mater. 2014, 14, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Marin, F.; Luquet, G.; Marie, B.; Medakovic, D. Molluscan Shell Proteins: Primary Structure, Origin, and Evolution. Curr. Top. Dev. Biol. 2008, 80, 209–276. [Google Scholar] [CrossRef] [PubMed]
- Wilbur, K.M.; Saleuddin, A.S.M. (Eds.) Shell Formation. In The Mollusca, Volume 4; Academic Press: New York, NY, USA, 1983; pp. 235–287. [Google Scholar]
- Marie, B.; Joubert, C.; Tayalé, A.; Zanella-Cléon, I.; Belliard, C.; Piquemal, D.; Cochennec-Laureau, N.; Marin, F.; Gueguen, Y.; Montagnani, C. Different Secretory Repertoires Control the Biomineralization Processes of Prism and Nacre Deposition of the Pearl Oyster Shell. Proc. Natl. Acad. Sci. USA 2012, 109, 20986–20991. [Google Scholar] [CrossRef] [PubMed]
- Mourea, G.; Vilarinho, L.; Santos, A.C.; Machado, J. Organic Compounds in the Extrapalial Fluid and Haemolymph of Anodonta Cygnea (L.) with Emphasis on the Seasonal Biomineralization Process. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2000, 125, 293–306. [Google Scholar] [CrossRef]
- Marin, F.; Narayanappa, P.; Motreuil, S. Acidic Shell Proteins of the Mediterranean Fan Mussel Pinna nobilis. In Molecular Biomineralization. Progress in Molecular and Subcellular Biology; Müller, W.E.G., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 52, pp. 353–396. [Google Scholar]
- Thula, T.T.; Svedlund, F.; Rodriguez, D.E.; Podschun, J.; Pendi, L.; Gower, L.B. Mimicking the Nanostructure of Bone: Comparison of Polymeric Process-Directing Agents. Polymers 2011, 3, 10–35. [Google Scholar] [CrossRef] [PubMed]
- Marin, F.; Amons, R.; Guichard, N.; Stigter, M.; Hecker, A.; Luquet, G.; Layrolle, P.; Alcaraz, G.; Riondet, C.; Westbroek, P. Caspartin and Calprismin, Two Proteins of the Shell Calcitic Prisms of the Mediterranean Fan Mussel Pinna nobilis. J. Biol. Chem. 2005, 280, 33895–33908. [Google Scholar] [CrossRef]
- Marin, F.; Luquet, G. Unusually Acidic Proteins in Biomineralization. In Handbook of Biomineralization: Biological Aspects and Structure Formation; Bäuerlein, E., Ed.; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2007; Volume 1. [Google Scholar]
- Hovden, R.; Wolf, S.E.; Holtz, M.E.; Marin, F.; Muller, D.A.; Estroff, L.A. Nanoscale Assembly Processes Revealed in the Nacroprismatic Transition Zone of Pinna nobilis Mollusc Shells. Nat. Commun. 2015, 6, 10097. [Google Scholar] [CrossRef]
- Teng, H.H.; Dove, P.M.; De Yoreo, J.J. Kinetics of Calcite Growth: Surface Processes and Relationships to Macroscopic Rate Laws. Geochim. Cosmochim. Acta 2000, 64, 2255–2266. [Google Scholar] [CrossRef]
- Nielsen, A.E. Rate Laws and Rate Constants in Crystal Growth. Croat. Chim. Acta 1987, 60, 531–539. [Google Scholar]
- Vielzeuf, D.; Garrabou, J.; Baronnet, A.; Grauby, O.; Marschal, C. Nano to Macroscale Biomineral Architecture of Red Coral (Corallium Rubrum). Am. Mineral. 2008, 93, 1799–1815. [Google Scholar] [CrossRef]
- Addadi, L.; Joester, D.; Nudelman, F.; Weiner, S. Mollusk Shell Formation: A Source of New Concepts for Understanding Biomineralization Processes. Chem. Eur. J. 2006, 12, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.E.; Böhm, C.F.; Harris, J.; Demmert, B.; Jacob, D.E.; Mondeshki, M.; Ruiz-Agudo, E.E.; Rodriguez-Navarro, C.; Rodríguez-Navarro, C. Nonclassical Crystallization in vivo et in vitro (I): Process-Structure-Property Relationships of Nanogranular Biominerals. J. Struct. Biol. 2016, 196, 260–287. [Google Scholar] [CrossRef] [PubMed]
- Weiner, S.; Addadi, L. Crystallization Pathways in Biomineralization. Annu. Rev. Mater. Res. 2011, 41, 21–40. [Google Scholar] [CrossRef]
- Watabe, N. Shell Repair. In The Mollusca, Physiology, Volume 4, Part 1; Wilbur, K.M., Saleuuddin, A., Eds.; Academic Press, Inc. (London) LTD: New York, NY, USA, 1983; pp. 289–310. [Google Scholar]
- Gower, L.B. Biomimetic Model Systems for Investigating the Amorphous Precursor Pathway and Its Role in Biomineralization. Chem. Rev. 2008, 108, 4551–4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, S.E.; Lieberwirth, I.; Natalio, F.; Bardeau, J.-F.; Delorme, N.; Emmerling, F.; Barrea, R.; Kappl, M.; Marin, F. Merging Models of Biomineralisation with Concepts of Nonclassical Crystallisation: Is a Liquid Amorphous Precursor Involved in the Formation of the Prismatic Layer of the Mediterranean Fan Mussel Pinna nobilis? Faraday Discuss. 2012, 159, 433–448. [Google Scholar] [CrossRef]
- Beniash, E.; Aizenberg, J.; Addadi, L.; Weiner, S. Amorphous calcium carbonate transforms into calcite during sea urchin larval spicule growth. Proc. R. Soc. Lond. B Biol. Sci. 1997, 264, 461–465. [Google Scholar] [CrossRef]
- Li, H.; Xin, H.L.; Kunitake, M.E.; Keene, E.C.; Muller, D.A.; Estroff, L.A.; Muller, A.; Estroff, L.A. Calcite Prisms from Mollusk Shells (Atrina Rigida): Swiss-Cheese-like Organic-Inorganic Single-Crystal Composites. Adv. Funct. Mater. 2011, 21, 2028–2034. [Google Scholar] [CrossRef]
- Falini, G.; Albeck, S.; Weiner, S.; Addadi, L.; Falini, G.; Albeck, S.; Weiner, S.; Addadit, L. Control of Aragonite or Calcite Polymorphism by Mollusk Shell Macromolecules. Science 1996, 271, 67–69. [Google Scholar] [CrossRef]
- Gong, Y.U.T.; Killian, C.E.; Olson, I.C.; Appathurai, N.P.; Amasino, A.L.; Martin, M.C.; Holt, L.J.; Wilt, F.H.; Gilbert, P.U.P.A. Phase Transitions in Biogenic Amorphous Calcium Carbonate. Proc. Natl. Acad. Sci. USA 2012, 109, 6088–6093. [Google Scholar] [CrossRef] [PubMed]
- Radha, A.V.; Forbes, T.Z.T.Z.; Killian, C.E.; Gilbert, P.U.P.A.; Navrotsky, A. Transformation and Crystallization Energetics of Synthetic and Biogenic Amorphous Calcium Carbonate. Proc. Natl. Acad. Sci. USA 2010, 107, 16438–16443. [Google Scholar] [CrossRef] [PubMed]
- De Yoreo, J.J.; Gilbert, P.U.P.A.; Sommerdijk, N.A.J.M.; Penn, R.L.; Whitelam, S.; Joester, D.; Zhang, H.; Rimer, J.D.; Navrotsky, A.; Banfield, J.F.; et al. Crystallization by Particle Attachment in Synthetic, Biogenic, and Geologic Environments. Science 2015, 349, aaa6760. [Google Scholar] [CrossRef] [PubMed]
- Albéric, M.; Bertinetti, L.; Zou, Z.; Fratzl, P.; Habraken, W.; Politi, Y. The Crystallization of Amorphous Calcium Carbonate Is Kinetically Governed by Ion Impurities and Water. Adv. Sci. 2017, 5, 1701000. [Google Scholar] [CrossRef]
- Killian, C.E.; Metzler, R.; Gong, Y.U.T.; Olson, I.C.; Aizenberg, J.; Politi, Y.; Wilt, F.H.; Scholl, A.; Young, A.; Doran, A.; et al. Mechanism of Calcite Co-Orientation in the Sea Urchin Tooth. J. Am. Chem. Soc. 2009, 131, 18404–18409. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-Y.; Schenk, A.S.; Ihli, J.; Kulak, A.N.; Hetherington, N.B.J.; Tang, C.C.; Schmahl, W.W.; Griesshaber, E.; Hyett, G.; Meldrum, F.C. A Critical Analysis of Calcium Carbonate Mesocrystals. Nat. Commun. 2014, 5, 4341. [Google Scholar] [CrossRef]
- Gal, A.; Kahil, K.; Vidavsky, N.; DeVol, R.T.; Gilbert, P.U.P.A.; Fratzl, P.; Weiner, S.; Addadi, L. Particle Accretion Mechanism Underlies Biological Crystal Growth from an Amorphous Precursor Phase. Adv. Funct. Mater. 2014, 24, 5420–5426. [Google Scholar] [CrossRef]
- Gal, A.; Weiner, S.; Addadi, L. A Perspective on Underlying Crystal Growth Mechanisms in Biomineralization: Solution Mediated Growth versus Nanosphere Particle Accretion. CrystEngComm 2015, 17, 2606–2615. [Google Scholar] [CrossRef]
- Harris, J.; Mey, I.; Hajir, M.; Mondeshki, M.; Wolf, S.E. Pseudomorphic Transformation of Amorphous Calcium Carbonate Films Follows Spherulitic Growth Mechanisms and Can Give Rise to Crystal Lattice Tilting. CrystEngComm 2015, 17, 6831–6837. [Google Scholar] [CrossRef]
- Gower, L.B.; Odom, D. Deposition of Calcium Carbonate Films by a Polymer-Induced Liquid-Precursor (PILP) Process. J. Cryst. Growth 2000, 210, 719–734. [Google Scholar] [CrossRef]
- Politi, Y.; Metzler, R.A.; Abrecht, M.; Gilbert, B.; Wilt, F.H.; Sagi, I.; Addadi, L.; Weinfurter, H.; Gilbert, P.U.P.A. Transformation Mechanism of Amorphous Calcium Carbonate into Calcite in the Sea Urchin Larval Spicule. Proc. Natl. Acad. Sci. USA 2008, 105, 17362–17366. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, P.U.P.A.; Metzler, R.A.; Zhou, D.; Scholl, A.; Doran, A.; Young, A.; Kunz, M.; Tamura, N.; Coppersmith, S.N. Gradual Ordering in Red Abalone Nacre. J. Am. Chem. Soc. 2008, 130, 17519–17527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Nielsen, M.H.; Freeman, C.L.; Hamm, L.M.; Tao, J.; Lee, J.R.I.; Han, T.Y.J.; Becker, U.; Harding, J.H.; Dove, P.M.; et al. The Thermodynamics of Calcite Nucleation at Organic Interfaces: Classical vs. Non-Classical Pathways. Faraday Discuss. 2012, 159, 509–523. [Google Scholar] [CrossRef]
- Cölfen, H.; Mann, S. Higher-Order Organization by Mesoscale Self-Assembly and Transformation of Hybrid Nanostructures. Angew. Chem. Int. Ed. 2003, 42, 2350–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergström, L.; Sturm Née Rosseeva, E.V.; Salazar-Alvarez, G.; Cölfen, H. Mesocrystals in Biominerals and Colloidal Arrays. Acc. Chem. Res. 2015, 48, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Cölfen, H. Bio-Inspired Mineralization Using Hydrophilic Polymers. Top. Curr. Chem. 2006, 271, 1–77. [Google Scholar] [CrossRef]
- Robb, D.T.; Privman, V. Model of Nanocrystal Formation in Solution by Burst Nucleation and Diffusional Growth. Langmuir 2008, 24, 26–35. [Google Scholar] [CrossRef]
- Gower, L.B.; Tirrell, D. Calcium Carbonate Films and Helices Grown in Solutions of Poly(Aspartate). J. Cryst. Growth 1998, 191, 153–160. [Google Scholar] [CrossRef]
- Rodríguez-Navarroa, C.; Ruiz-Agudo, E.; Harris, J.; Wolf, S.E. Nonclassical crystallization in vivo et in vitro (II): Nanogranular features in biomimetic minerals disclose a general colloid-mediated crystal growth mechanism. J. Struct. Biol. 2016, 196, 260–287. [Google Scholar] [CrossRef]
- Wolf, S.E.; Leiterer, J.; Kappl, M.; Emmerling, F.; Tremel, W. Early Homogenous Amorphous Precursor Stages of Calcium Carbonate and Subsequent Crystal Growth in Levitated Droplets. J. Am. Chem. Soc. 2008, 130, 12342–12347. [Google Scholar] [CrossRef] [PubMed]
- Tartaj, P.; Amarilla, J.M. Multifunctional Response of Anatase Nanostructures Based on 25 Nm Mesocrystal-Like Porous Assemblies. Adv. Mater. 2011, 23, 4904–4907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.-Y.; Long, L.-L.; Li, W.-W.; Wang, W.-K.; Yu, H.-Q. Hexagonal Microrods of Anatase Tetragonal TiO2: Self-Directed Growth and Superior Photocatalytic Performance. Chem. Commun. 2013, 49, 6075. [Google Scholar] [CrossRef] [PubMed]
- Song, R.Q.; Krasia-Christoforou, T.; Debus, C.; Cölfen, H. Structure and Magnetic Property Control of Copper Hydroxide Acetate by Non-Classical Crystallization. Small 2017, 13, 1602702. [Google Scholar] [CrossRef] [PubMed]
- Song, R.-Q.; Cölfen, H. Mesocrystals-Ordered Nanoparticle Superstructures. Adv. Mater. 2010, 22, 1301–1330. [Google Scholar] [CrossRef] [PubMed]
- Song, R.-Q.; Cölfen, H.; Xu, A.-W.; Hartmann, J.; Antonietti, M. Polyelectrolyte-Directed Nanoparticle Aggregation: Systematic Morphogenesis of Calcium Carbonate by Nonclassical Crystallization. ACS Nano 2009, 3, 1966–1978. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xie, T.; Peng, Q.; Li, Y. Ag, Ag2S, and Ag2Se Nanocrystals: Synthesis, Assembly, and Construction of Mesoporous Structures. J. Am. Chem. Soc. 2008, 130, 4016–4022. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Antonietti, M.; Cölfen, H. Calcite Mesocrystals: “Morphing” Crystals by a Polyelectrolyte. Chemistry 2006, 12, 5722–5730. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cölfen, H.; Antonietti, M. Nonclassical Crystallization: Mesocrystals and Morphology Change of CaCO3 Crystals in the Presence of a Polyelectrolyte Additive. J. Am. Chem. Soc. 2005, 127, 3246–3247. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, Y.; Wu, H.; Guo, S. Control of the Formation of Rod-like ZnO Mesocrystals and Their Photocatalytic Properties. CrystEngComm 2013, 15, 2608–2615. [Google Scholar] [CrossRef]
- Da Silva, R.O.; Gonçalves, R.H.; Stroppa, D.G.; Ramirez, A.J.; Leite, E.R. Synthesis of Recrystallized Anatase TiO2 Mesocrystals with Wulff Shape Assisted by Oriented Attachment. Nanoscale 2011, 3, 1910–1916. [Google Scholar] [CrossRef] [PubMed]
- Schenk, A.S.; Eiben, S.; Goll, M.; Reith, L.; Kulak, A.N.; Meldrum, F.C.; Jeske, H.; Wege, C.; Ludwigs, S. Virus-Directed Formation of Electrocatalytically Active Nanoparticle-Based Co3O4tubes. Nanoscale 2017, 9, 6334–6345. [Google Scholar] [CrossRef] [PubMed]
- Ross, M.B.; Ku, J.C.; Vaccarezza, V.M.; Schatz, G.C.; Mirkin, C.A. Nanoscale Form Dictates Mesoscale Function in Plasmonic DNA–Nanoparticle Superlattices. Nat. Nanotechnol. 2015, 10, 453–458. [Google Scholar] [CrossRef]
- Ortega, S.; Ibáñez, M.; Liu, Y.; Zhang, Y.; Kovalenko, M.V.; Cadavid, D.; Cabot, A. Bottom-up Engineering of Thermoelectric Nanomaterials and Devices from Solution-Processed Nanoparticle Building Blocks. Chem. Soc. Rev. 2017, 46, 3510–3528. [Google Scholar] [CrossRef] [PubMed]
- Sang, L.; Zhao, Y.; Burda, C. TiO2 Nanoparticles as Functional Building Blocks. Chem. Rev. 2014, 114, 9283–9318. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-W.; Christenson, H.K.; Meldrum, F.C. Confinement Leads to Control over Calcium Sulfate Polymorph. Adv. Funct. Mater. 2013, 23, 5615–5623. [Google Scholar] [CrossRef]
- Uebe, R.; Schüler, D. Magnetosome Biogenesis in Magnetotactic Bacteria. Nat. Rev. Microbiol. 2016, 14, 621–637. [Google Scholar] [CrossRef]
- Smeets, P.J.M.; Finney, A.R.; Habraken, W.J.E.M.; Nudelman, F.; Friedrich, H. A Classical View on Nonclassical Nucleation. Proc. Natl. Acad. Sci. USA 2017, 114, E7882–E7890. [Google Scholar] [CrossRef]
- Wolf, S.E.; Gower, L.B. Challenges and Perspectives of the Polymer-Induced Liquid-Precursor Process: The Pathway from Liquid-Condensed Mineral Precursors to Mesocrystalline Products. In New Perspectives on Mineral Nucleation and Growth; van Driessche, A.E.S., Kellermeier, M., Benning, L.G., Gebauer, D., Eds.; Springer: Cham, Switzerland, 2017; pp. 43–75. [Google Scholar]
- Cölfen, H.; Antonietti, M. Mesocrystals and Nonclassical Crystallization; Cölfen, H., Antonietti, M., Eds.; Wiley-VCH Verlag GmbH: Berlin, Germany, 2008. [Google Scholar]
- Niederberger, M.; Cölfen, H. Oriented Attachment and Mesocrystals: Non-Classical Crystallization Mechanisms Based on Nanoparticle Assembly. Phys. Chem. Chem. Phys. 2006, 8, 3271–3287. [Google Scholar] [CrossRef]
- Wallace, A.F.; Hedges, L.O.; Fernandez-Martinez, A.; Raiteri, P.; Gale, J.D.; Waychunas, G.A.; Whitelam, S.; Banfield, J.F.; De Yoreo, J.J. Microscopic Evidence for Liquid-Liquid Separation in Supersaturated CaCO3 Solutions. Science 2013, 341, 885–889. [Google Scholar] [CrossRef]
- Kim, Y.-Y.; Schenk, A.S.; Walsh, D.; Kulak, A.N.; Cespedes, O.; Meldrum, F.C. Bio-Inspired Formation of Functional Calcite/Metal Oxide Nanoparticle Composites. Nanoscale 2014, 6, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Schenk, A.S.; Zlotnikov, I.; Pokroy, B.; Gierlinger, N.; Masic, A.; Zaslansky, P.; Fitch, A.N.; Paris, O.; Metzger, T.H.; Cölfen, H.; et al. Hierarchical Calcite Crystals with Occlusions of a Simple Polyelectrolyte Mimic Complex Biomineral Structures. Adv. Funct. Mater. 2012, 22, 4668–4676. [Google Scholar] [CrossRef] [Green Version]
- Barthelat, F.; Rim, J.E.; Espinosa, H.D. A Review on the Structure and Mechanical Properties of Mollusk Shells—Perspectives on Synthetic Biomimetic Materials. In Applied Scanning Probe Methods XIII, Biomimetics and Industrial Applications; Bhushan, B., Fuchs, H., Eds.; Springer: Berlin, Germany, 2009; pp. 17–41. [Google Scholar]
- Villanova, J.; Kozachkevich, S.; Zaslansky, P.; Kundanati, L.; Bracha, A.A.; Polishchuk, I.; Bloch, L.; Levy, D.; Katsman, A.; Giacobbe, C.; et al. Coherently Aligned Nanoparticles within a Biogenic Single Crystal: A Biological Prestressing Strategy. Science 2017, 358, 1294–1298. [Google Scholar] [CrossRef]
- Sethmann, I.; Hinrichs, R.; Wörheide, G.; Putnis, A. Nano-Cluster Composite Structure of Calcitic Sponge Spicules—A Case Study of Basic Characteristics of Biominerals. J. Inorg. Biochem. 2006, 100, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Addadi, L.; Weiner, S. Biomineralization: Mineral Formation by Organisms. Phys. Scr. 2014, 89, 098003. [Google Scholar] [CrossRef]
- Weiner, S.; Addadi, L. Design Strategies in Mineralized Biological Materials. J. Mater. Chem. 1997, 7, 689–702. [Google Scholar] [CrossRef]
- Gebauer, D.; Wolf, S.E. Designing Solid Materials from their Solute State: a Shift in Paradigms toward a Holistic Approach in Functional Materials Chemistry. J. Am. Chem. Soc. 2019, 141, 4490–4504. [Google Scholar] [CrossRef]
- Wegst, U.G.K.; Ashby, M.F. The Mechanical Efficiency of Natural Materials. Philos. Mag. 2004, 84, 2167–2186. [Google Scholar] [CrossRef]
- Meyers, M.A.; Chen, P.-Y.; Lin, A.Y.-M.; Seki, Y. Biological Materials: Structure and Mechanical Properties. Prog. Mater. Sci. 2008, 53, 1–206. [Google Scholar] [CrossRef]
- Pokroy, B.; Zolotoyabko, E. Microstructure of Natural Plywood-like Ceramics: A Study by High-Resolution Electron Microscopy and Energy-Variable X-Ray Diffraction. J. Mater. Chem. 2003, 13, 682–688. [Google Scholar] [CrossRef]
- Tai, K.; Ulm, F.J.; Ortiz, C. Nanogranular Origins of the Strength of Bone. Nano Lett. 2006, 6, 2520–2525. [Google Scholar] [CrossRef] [PubMed]
- Ryall, R.L.; Fleming, D.E.; Doyle, I.R.; Evans, N.A.; Dean, C.J.; Marshall, V.R. Intracrystalline Proteins and the Hidden Ultrastructure of Calcium Oxalate Urinary Crystals: Implications for Kidney Stone Formation. J. Struct. Biol. 2001, 134, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Böhm, C.F.; Demmert, B.; Harris, J.; Fey, T.; Marin, F.; Wolf, S.E. Structural Commonalities and Deviations in the Hierarchical Organization of Crossed-Lamellar Shells: A Case Study on the Shell of the Bivalve Glycymeris Glycymeris. J. Mater. Res. 2016, 31, 536–546. [Google Scholar] [CrossRef]
- Schenk, A.S.; Kim, Y.Y. Unraveling the Internal Microstructure of Biogenic and Bioinspired Calcite Single Crystals. MRS Bull. 2015, 40, 499–508. [Google Scholar] [CrossRef]
- Mutvei, H.; Dunca, E. Crystalline Structure, Orientation and Nucleation of the Nacreous Tablets in the Cephalopod Nautilus. Paläontologische Zeitschrift 2010, 84, 457–465. [Google Scholar] [CrossRef]
- Checa, A.G.; Mutvei, H.; Osuna-Mascaró, A.J.; Bonarski, J.T.; Faryna, M.; Berent, K.; Pina, C.M.; Rousseau, M.; Macías-Sánchez, E. Crystallographic Control on the Substructure of Nacre Tablets. J. Struct. Biol. 2013, 183, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Athanasiadou, D.; Jiang, W.; Goldbaum, D.; Saleem, A.; Basu, K.; Pacella, M.S.; Böhm, C.F.; Chromik, R.R.; Hincke, M.T.; Rodríguez-Navarro, A.B.; et al. Nanostructure, Osteopontin, and Mechanical Properties of Calcitic Avian Eggshell. Sci. Adv. 2018, 4, eaar3219. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A.B.; Marie, P.; Nys, Y.; Hincke, M.T.; Gautron, J. Amorphous Calcium Carbonate Controls Avian Eggshell Mineralization: A New Paradigm for Understanding Rapid Eggshell Calcification. J. Struct. Biol. 2015, 190, 291–303. [Google Scholar] [CrossRef]
- Seto, J.; Ma, Y.; Davis, S.A.; Meldrum, F.C.; Gourrier, A.; Kim, Y.-Y.; Schilde, U.; Sztucki, M.; Burghammer, M.; Maltsev, S.; et al. Structure-Property Relationships of a Biological Mesocrystal in the Adult Sea Urchin Spines. Proc. Natl. Acad. Sci. USA 2012, 109, 3699–3704. [Google Scholar] [CrossRef]
- Dauphin, Y.; Cuif, J.P.; Doucet, J.; Salomé, M.; Susini, J.; Willams, C.T. In Situ Chemical Speciation of Sulfur in Calcitic Biominerals and the Simple Prism Concept. J. Struct. Biol. 2003, 142, 272–280. [Google Scholar] [CrossRef]
- Dauphin, Y. The Nanostructural Unity of Mollusc Shells. Mineral. Mag. 2008, 72, 243–246. [Google Scholar] [CrossRef]
- Jacob, D.E.; Soldati, A.; Wirth, R.; Huth, J.; Wehrmeister, U.; Hofmeister, W. Nanostructure, Composition and Mechanisms of Bivalve Shell Growth. Geochim. Cosmochim. Acta 2008, 72, 5401–5415. [Google Scholar] [CrossRef]
- Checa, A.G.; Bonarski, J.T.; Willinger, M.G.; Faryna, M.; Berent, K.; Kania, B.; González-Segura, A.; Pina, C.M.; Pospiech, J.; Morawiec, A. Crystallographic Orientation Inhomogeneity and Crystal Splitting in Biogenic Calcite. J. R. Soc. Interface 2013, 10, 20130425. [Google Scholar] [CrossRef]
- Okumura, T.; Suzuki, M.; Nagasawa, H.; Kogure, T. Microstructural Variation of Biogenic Calcite with Intracrystalline Organic Macromolecules. Cryst. Growth Des. 2012, 12, 224–230. [Google Scholar] [CrossRef]
- Suzuki, M.; Kameda, J.; Sasaki, T.; Saruwatari, K.; Nagasawa, H.; Kogure, T. Characterization of the Multilayered Shell of a Limpet, Lottia Kogamogai (Mollusca: Patellogastropoda), Using SEM–EBSD and FIB–TEM Techniques. J. Struct. Biol. 2010, 171, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Aizenberg, J.; Hanson, J.; Koetzle, T.F.; Weinfurter, H.; Addadi, L.; York, N.; August, R.V. Control of Macromolecule Distribution within Synthetic and Biogenic Single Calcite Crystals. J. Am. Chem. Soc. 1997, 119, 881–886. [Google Scholar] [CrossRef]
- Berman, A.; Addadi, L.; Kvick, A.Y.E.; Leiserowitz, L.; Nelson, M.; Weinfurter, H. Intercalation of Sea Urchin Proteins in Calcite: Study of a Crystalline Composite Material. Science 1990, 250, 664–667. [Google Scholar] [CrossRef] [Green Version]
- Pisklak, D.M.; Szeleszczuk, Ł.; Wawer, I. 1H and 13C Magic-Angle Spinning Nuclear Magnetic Resonance Studies of the Chicken Eggshell. J. Agric. Food Chem. 2012, 60, 12254–12259. [Google Scholar] [CrossRef]
- Ben Shir, I.; Kababya, S.; Katz, I.; Pokroy, B.; Schmidt, A. Exposed and Buried Biomineral Interfaces in the Aragonitic Shell of Perna canaliculus Revealed by Solid-State NMR. Chem. Mater. 2013, 25, 4595–4602. [Google Scholar] [CrossRef]
- Lee, D.; Leroy, C.; Crevant, C.; Bonhomme-Coury, L.; Babonneau, F.; Laurencin, D.; Bonhomme, C.; De Paëpe, G.; Bryce, D.L.; Laurencin, D.; et al. Interfacial Ca2+ Environments in Nanocrystalline Apatites Revealed by Dynamic Nuclear Polarization Enhanced 43Ca NMR Spectroscopy. Nat. Commun. 2017, 8, 14104. [Google Scholar] [CrossRef]
- Agbaje, O.B.A.; Ben Shir, I.; Zax, D.B.; Schmidt, A.; Jacob, D.E. Biomacromolecules within Bivalve Shells: Is Chitin Abundant? Acta Biomater. 2018, 80, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Bar-On, B.; Wagner, H.D. Structural Motifs and Elastic Properties of Hierarchical Biological Tissues—A Review. J. Struct. Biol. 2013, 183, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Meyers, M.A.; Lin, A.; Chen, P.; Muyco, J. Mechanical Strength of Abalone Nacre: Role of the Soft Organic Layer. J. Mech. Behav. Biomed. Mater. 2008, 1, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Meyers, M.A.; Zhang, Z.; Ritchie, R.O. Functional Gradients and Heterogeneities in Biological Materials: Design Principles, Functions, and Bioinspired Applications. Prog. Mater. Sci. 2017, 88, 467–498. [Google Scholar] [CrossRef]
- Currey, J.D.; Taylor, J.D. The Mechanical Behaviour of Some Molluscan Hard Tissues. J. Zool. 1974, 173, 395–406. [Google Scholar] [CrossRef]
- Chen, P.-Y.; McKittrick, J.; Meyers, M.A. Biological Materials: Functional Adaptations and Bioinspired Designs. Prog. Mater. Sci. 2012, 57, 1492–1704. [Google Scholar] [CrossRef]
- Bayerlein, B.; Bertinetti, L.; Bar-on, B.; Blumtritt, H.; Fratzl, P. Inherent Role of Water in Damage Tolerance of the Prismatic Mineral-Organic Biocomposite in the Shell of Pinna nobilis. Adv. Fcunt. Mater. 2016, 26, 3663–3669. [Google Scholar] [CrossRef]
- Cuif, J.-P.; Burghammer, M.; Chamard, V.; Dauphin, Y.; Godard, P.; Moullac, G.; Nehrke, G.; Perez-Huerta, A. Evidence of a Biological Control over Origin, Growth and End of the Calcite Prisms in the Shells of Pinctada Margaritifera (Pelecypod, Pterioidea). Minerals 2014, 4, 815–834. [Google Scholar] [CrossRef]
- Nakahara, H.; Kakei, M.; Bevelander, G. Fine Structure and Amino Acid Composition of the Organic “Envelope” in the Prismatic Layer of Some Bivlave Shells. Jpn. J. Malacol. 1980, 39, 167–177. [Google Scholar]
- Gilbert, P.U.P.A.; Young, A.; Coppersmith, S.N. Measurement of C-Axis Angular Orientation in Calcite (CaCO3) Nanocrystals Using X-Ray Absorption Spectroscopy. Proc. Natl. Acad. Sci. USA 2011, 108, 11350–11355. [Google Scholar] [CrossRef]
- Kunitake, M.E.; Mangano, L.M.; Peloquin, J.M.; Baker, S.P.; Estroff, L.A. Evaluation of Strengthening Mechanisms in Calcite Single Crystals from Mollusk Shells. Acta Biomater. 2013, 9, 5353–5359. [Google Scholar] [CrossRef] [PubMed]
- Kuhn-Spearing, L.T.; Kessler, H.; Chateau, E.; Ballarini, R.; Heuer, A.H.; Spearing, S.M. Fracture Mechanisms of the Strombus Gigas Conch Shell: Implications for the Design of Brittle Laminates. J. Mater. Sci. 1996, 31, 6583–6594. [Google Scholar] [CrossRef]
- Kamat, S.; Su, X.; Ballarini, R.; Heuer, A. Structural Basis for the Fracture Toughness of the Shell of the Conch Strombus Gigas. Nature 2000, 405, 1036–1040. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.C.; Milliron, G.W.; Miserez, A.; Evans-Lutterodt, K.; Herrera, S.; Gallana, I.; Mershon, W.J.; Swanson, B.; Zavattieri, P.; DiMasi, E.; et al. The Stomatopod Dactyl Club: A Formidable Damage-Tolerant Biological Hammer. Science 2012, 336, 1275–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunenfelder, L.K.; Suksangpanya, N.; Salinas, C.; Milliron, G.; Yaraghi, N.; Herrera, S.; Evans-Lutterodt, K.; Nutt, S.R.; Zavattieri, P.; Kisailus, D. Bio-Inspired Impact-Resistant Composites. Acta Biomater. 2014, 10, 3997–4008. [Google Scholar] [CrossRef] [PubMed]
- Guarín-Zapata, N.; Gomez, J.; Yaraghi, N.; Kisailus, D.; Zavattieri, P.D. Shear Wave Filtering in Naturally-Occurring Bouligand Structures. Acta Biomater. 2015, 23, 11–20. [Google Scholar] [CrossRef] [PubMed]
- de Obaldia, E.E.; Jeong, C.; Grunenfelder, L.K.; Kisailus, D.; Zavattieri, P. Analysis of the Mechanical Response of Biomimetic Materials with Highly Oriented Microstructures through 3D Printing, Mechanical Testing and Modeling. J. Mech. Behav. Biomed. Mater. 2015, 48, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Mirkhalaf, M.; Dastjerdi, A.K.; Barthelat, F. Overcoming the Brittleness of Glass through Bio-Inspiration and Micro-Architecture. Nat. Commun. 2014, 5, 3166. [Google Scholar] [CrossRef]
- Ritchie, R.O. The Conflicts between Strength and Toughness. Nat. Mater. 2011, 10, 817–822. [Google Scholar] [CrossRef]
- Weißbach, W.; Dahms, M.; Jaroschek, C. Werkstoffkunde; Springer: Wiesbaden, Germany, 2015. [Google Scholar]
- Turner, F.J.; Griffs, D.T.; Heard, H. Experimental Deformation of Calcite Crystals. Geol. Soc. Am. Bull. 1954, 65, 883–934. [Google Scholar] [CrossRef]
- Carlton, C.E.; Ferreira, P.J. What Is behind the Inverse Hall-Petch Effect in Nanocrystalline Materials? Acta Mater. 2007, 55, 3749–3756. [Google Scholar] [CrossRef]
- Wiederhorn, S.M. Brittle Fracture and Toughening Mechanisms in Ceramics. Annu. Rev. Mater. Sci. 1984, 14, 373–403. [Google Scholar] [CrossRef]
- Salmang, H.; Scholze, H. Keramik; Springer-Verlag: Berlin/Heidelberg, Germany, 2007. [Google Scholar] [CrossRef]
- Schamel, M.; Barralet, J.E.; Gelinsky, M.; Groll, J.; Gbureck, U. Intrinsic 3D Prestressing: A New Route for Increasing Strength and Improving Toughness of Hybrid Inorganic Biocements. Adv. Mater. 2017, 29, 1701035. [Google Scholar] [CrossRef] [PubMed]
- Barthelat, F.; Tang, H.; Zavattieri, P.; Li, C.; Espinosa, H. On the Mechanics of Mother-of-Pearl: A Key Feature in the Material Hierarchical Structure. J. Mech. Phys. Solids 2007, 55, 306–337. [Google Scholar] [CrossRef]
- Fantner, G.E.; Hassenkam, T.; Kindt, J.H.; Weaver, J.C.; Birkedal, H.; Pechenik, L.; Cutroni, J.A.; Cidade, G.A.G.; Stucky, G.D.; Morse, D.E.; et al. Sacrificial Bonds and Hidden Length Dissipate Energy as Mineralized Fibrils Separate during Bone Fracture. Nat. Mater. 2005, 4, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, Z.-H.; Wang, R. In Situ Observation of Nanograin Rotation and Deformation in Nacre. Nano Lett. 2006, 6, 2301–2304. [Google Scholar] [CrossRef]
- Gordon, L.M.; Joester, D. Nanoscale Chemical Tomography of Buried Organic-Inorganic Interfaces in the Chiton Tooth. Nature 2011, 469, 194–197. [Google Scholar] [CrossRef]
- Barthelat, F.F. Biomimetics for next Generation Materials. Philos. Trans. A Math. Phys. Eng. Sci. 2007, 365, 2907–2919. [Google Scholar] [CrossRef]
- Taylor, J.D.; Layman, M. The Mechanical Properties of Bivalve (Mollusca) Shell Structure. Paleontology 1972, 15, 73–87. [Google Scholar]
- Marin, F.; Luquet, G. Molluscan Shell Proteins. C. R. Palevol 2004, 3, 469–492. [Google Scholar] [CrossRef]
- Checa, A.G.; Cartwright, J.H.E.; Willinger, M.G. Mineral Bridges in Nacre. J. Struct. Biol. 2011, 176, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Z.; Suo, Z.; Evans, A.G.; Yao, N.; Aksay, I.A. Deformation Mechanisms in Nacre. J. Mater. Res. 2011, 16, 2485–2493. [Google Scholar] [CrossRef]
- Heinemann, F.; Launspach, M.; Gries, K.; Fritz, M. Gastropod Nacre: Structure, Properties and Growth—Biological, Chemical and Physical Basics. Biophys. Chem. 2011, 153, 126–153. [Google Scholar] [CrossRef] [PubMed]
- Barthelat, F.; Li, C.-M.; Comi, C.; Espinosa, H.D. Mechanical Properties of Nacre Constituents and Their Impact on Mechanical Performance. J. Mater. Res. 2006, 21, 1977–1986. [Google Scholar] [CrossRef]
- Weber, E.; Pokroy, B. Intracrystalline Inclusions within Single Crystalline Hosts: From Biomineralization to Bio-Inspired Crystal Growth. CrystEngComm 2015, 17, 5873–5883. [Google Scholar] [CrossRef]
- Levi-Kalisman, Y.; Falini, G.; Addadi, L.; Weinfurter, H. Structure of the Nacreous Organic Matrix of a Bivalve Mollusk Shell Examined in the Hydrated State Using Cryo-TEM. J. Struct. Biol. 2001, 135, 8–17. [Google Scholar] [CrossRef]
- Younis, S.; Kauffmann, Y.; Bloch, L.; Zolotoyabko, E. Inhomogeneity of Nacre Lamellae on the Nanometer Length Scale. Cryst. Growth Des. 2012, 12, 4574–4579. [Google Scholar] [CrossRef]
- Kim, Y.-Y.; Carloni, J.D.; Demarchi, B.; Sparks, D.; Reid, D.G.; Kunitake, M.E.; Tang, C.C.; Duer, M.J.; Freeman, C.L.; Pokroy, B.; et al. Tuning Hardness in Calcite by Incorporation of Amino Acids. Nat. Mater. 2016, 15, 903–910. [Google Scholar] [CrossRef]
- Koyama, M.; Zhang, Z.; Wang, M.; Ponge, D.; Raabe, D.; Tsuzaki, K.; Noguchi, H.; Tasan, C.C. Bone-like Crack Resistance in Hierarchical Metastable Nanolaminate Steels. Science 2017, 355, 1055–1057. [Google Scholar] [CrossRef]
- Cao, S.C.; Liu, J.; Zhu, L.; Li, L.; Dao, M.; Lu, J.; Ritchie, R.O. Nature-Inspired Hierarchical Steels. Sci. Rep. 2018, 8, 5088. [Google Scholar] [CrossRef]
- Wat, A.; Lee, J.I.; Ryu, C.W.; Gludovatz, B.; Kim, J.; Tomsia, A.P.; Ishikawa, T.; Schmitz, J.; Meyer, A.; Alfreider, M.; et al. Bioinspired Nacre-like Alumina with a Bulk-Metallic Glass-Forming Alloy as a Compliant Phase. Nat. Commun. 2019, 10, 961. [Google Scholar] [CrossRef] [PubMed]
- Kilper, S.; Facey, S.J.; Burghard, Z.; Hauer, B.; Rothenstein, D.; Bill, J. Macroscopic Properties of Biomimetic Ceramics Are Governed by the Molecular Recognition at the Bioorganic—Inorganic Interface. Adv. Funct. Mater. 2018, 28, 1705842. [Google Scholar] [CrossRef]
- Eder, M.; Amini, S.; Fratzl, P. Biological Composites—Complex Structures for Functional Diversity. Science 2018, 362, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Aizenberg, J.; Weiner, S.; Tkachenko, A.; Addadi, L.; Hendler, G. Calcitic Microlenses as Part of the Photoreceptor System in Brittlestars. Nature 2001, 412, 819–822. [Google Scholar] [CrossRef] [PubMed]
- Nys, Y.; Gautron, J. Structure and Formation of the Eggshell. In Bioactive Egg Compounds; Anton, M., Schade, R., Huopalahti, R., López-Fandino, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–4. [Google Scholar]
- Aizenberg, J.; Weaver, J.C.; Thanawala, M.S.; Sundar, V.C.; Morse, D.E.; Fratzl, P. Skeleton of Euplectella sp.: Structural Hierarchy from the Nanoscale to the Macroscale. Science 2005, 309, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Weaver, J.C.; Aizenberg, J.; Fantner, G.E.; Kisailus, D.; Woesz, A.; Allen, P.; Fields, K.; Porter, M.J.; Zok, F.W.; Hansma, P.K.; et al. Hierarchical Assembly of the Siliceous Skeletal Lattice of the Hexactinellid Sponge Euplectella Aspergillum. J. Struct. Biol. 2007, 158, 93–106. [Google Scholar] [CrossRef]
- Ehrlich, H.; Deutzmann, R.; Brunner, E.; Cappellini, E.; Koon, H.; Solazzo, C.; Yang, Y.; Ashford, D.; Thomas-Oates, J.; Lubeck, M.; et al. Mineralization of the Metre-Long Biosilica Structures of Glass Sponges Is Templated on Hydroxylated Collagen. Nat. Chem. 2010, 2, 1084–1088. [Google Scholar] [CrossRef]
- Ehrlich, H. Chitin and Collagen as Universal and Alternative Templates in Biomineralization. Int. Geol. Rev. 2010, 52, 661–699. [Google Scholar] [CrossRef]
- Kröger, N.; Deutzmann, R.; Sumper, M. Polycationic Peptides from Diatom Biosilica That Direct Silica Nanosphere Formation. Science 1999, 286, 1129–1132. [Google Scholar] [Green Version]
- Matsunaga, S.; Sakai, R.; Jimbo, M.; Kamiya, H. Long-Chain Polyamines (LCPAs) from Marine Sponge: Possible Implication in Spicule Formation. Chembiochem 2007, 8, 1729–1735. [Google Scholar] [CrossRef]
- Poulsen, N.; Kröger, N. Silica Morphogenesis by Alternative Processing of Silaffins in the Diatom Thalassiosira Pseudonana. J. Chem. Phys. 2004, 279, 42993–42999. [Google Scholar] [CrossRef]
- Shimizu, K.; Cha, J.; Stucky, G.D. Silicatein: Cathepsin L-like Protein in Sponge Biosilica. Proc. Natl. Acad. Sci. USA 1998, 95, 6234–6238. [Google Scholar] [CrossRef] [PubMed]
- Sundar, V.C.; Yablon, A.D.; Grazul, J.L.; Ilan, M.; Aizenberg, J. Fibre-Optical Features of a Glass Sponge. Nature 2003, 424, 899–900. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.G.; Wolf, S.E.; Schlossmacher, U.; Wang, X.-H.; Boreiko, A.; Brandt, D.; Tremel, W.; Schröder, H.-C. Poly(Silicate)-Metabolizing Silicatein in Siliceous Spicules and Silicasomes of Demosponges Comprises Dual Enzymatic Activities (Silica Polymerase and Silica Esterase). FEBS J. 2008, 275, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.N.; Shimizu, K.; Zhou, Y.; Christiansen, S.C.; Chmelka, B.F.; Stucky, G.D.; Morse, D.E. Silicatein Filaments and Subunits from a Marine Sponge Direct the Polymerization of Silica and Silicones in Vitro. Proc. Natl. Acad. Sci. USA 1999, 96, 361–365. [Google Scholar] [CrossRef]
- Zhou, Y.; Shimizu, K.; Cha, J.N.; Stucky, G.D.; Morse, D.E. Efficient Catalysis of Polysiloxane Synthesis by Silicatein α Requires Specific Hydroxy and Imidazole Functionalities. Angew. Chem. Int. Ed. 1999, 38, 780–782. [Google Scholar] [CrossRef]
- Shimizu, K.; Morse, D.E. Silicatein: A Unique Silica-Synthesizing Catalytic Triad Hydrolase From Marine Sponge Skeletons and Its Multiple Applications. Methods Enzymol. 2018, 605, 429–455. [Google Scholar] [CrossRef]
- Brutchey, R.L.; Morse, D.E. Silicatein and the Translation of Its Molecular Mechanism of Biosilicification into Low Temperature Nanomaterial Synthesis. Chem. Rev. 2008, 108, 4915–4934. [Google Scholar] [CrossRef]
- Kisailus, D.; Choi, J.H.; Weaver, J.C.; Yang, W.; Morse, D.E. Enzymatic Synthesis and Nanostructural Control of Gallium Oxide at Low Temperature. Adv. Mater. 2005, 17, 314–318. [Google Scholar] [CrossRef]
- Sumerel, J.L.; Yang, W.J.; Kisailus, D.; Weaver, J.C.; Choi, J.H.; Morse, D.E. Biocatalytically Templated Synthesis of Titanium Dioxide. Chem. Mater. 2003, 15, 4804–4809. [Google Scholar] [CrossRef]
- Curnow, P.; Bessette, P.H.; Kisailus, D.; Murr, M.M.; Daugherty, P.S.; Morse, D.E. Enzymatic Synthesis of Layered Titanium Phosphates at Low Temperature and Neutral PH by Cell-Surface Display of Silicatein-α. J. Am. Chem. Soc. 2005, 127, 15749–15755. [Google Scholar] [CrossRef]
- Wolf, S.E.; Schlossmacher, U.; Pietuch, A.; Mathiasch, B.; Schröder, H.-C.; Müller, W.E.G.; Tremel, W. Formation of Silicones Mediated by the Sponge Enzyme Silicatein-α. Dalton Trans. 2010, 39, 9245–9249. [Google Scholar] [CrossRef]
- O’Leary, P.; van Walree, C.A.; Mehendale, N.C.; Sumerel, J.; Morse, D.E.; Kaska, W.C.; van Koten, G.; Gebbink, R.J.M. Enzymatic Immobilization of Organometallic Species: Biosilification of NCN- and PCP-Pincer Metal Species Using Demosponge Axial Filaments. Dalton Trans. 2009, 22, 4289–4291. [Google Scholar] [CrossRef]
- Tahir, M.N.; Théato, P.; Müller, W.E.G.; Schröder, H.-C.; Janshoff, A.; Zhang, J.; Huth, J.; Tremel, W. Monitoring the Formation of Biosilica Catalysed by Histidine-Tagged Silicatein. Chem. Commun. 2004, 2848–2849. [Google Scholar] [CrossRef]
- Tahir, M.N.; Théato, P.; Müller, W.E.G.; Schröder, H.C.; Borejko, A.; Faiß, S.; Janshoff, A.; Huth, J.; Tremel, W. Formation of Layered Titania and Zirconia Catalysed by Surface-Bound Silicatein. Chem. Commun. 2005, 5533–5535. [Google Scholar] [CrossRef]
- Tahir, M.N.; Eberhardt, M.; Therese, H.A.; Kolb, U.; Theato, P.; Müller, W.E.G.; Schröder, H.-C.; Tremel, W. From Single Molecules to Nanoscopically Structured Functional Materials: Au Nanocrystal Growth on TiO2 Nanowires Controlled by Surface-Bound Silicatein. Angew. Chem. Int. Ed. 2006, 45, 4803–4809. [Google Scholar] [CrossRef]
- Shukoor, M.I.; Natalio, F.; Therese, H.A.; Tahir, M.N.; Ksenofontov, V.; Panthöfer, M.; Eberhardt, M.; Theato, P.; Schröder, H.C.; Müller, W.E.G.; et al. Fabrication of a Silica Coating on Magnetic γ-Fe2O3 Nanoparticles by an Immobilized Enzyme. Chem. Mater. 2008, 20, 3567–3573. [Google Scholar] [CrossRef]
- Natalio, F.; Link, T.; Müller, W.E.G.; Schröder, H.C.; Cui, F.-Z.; Wang, X.; Wiens, M. Bioengineering of the Silica-Polymerizing Enzyme Silicatein-α for a Targeted Application to Hydroxyapatite. Acta Biomater. 2010, 6, 3720–3728. [Google Scholar] [CrossRef]
- Natalio, F.; Corrales, T.P.; Panthofer, M.; Schollmeyer, D.; Lieberwirth, I.; Muller, W.E.G.; Kappl, M.; Butt, H.-J.; Tremel, W. Flexible Minerals: Self-Assembled Calcite Spicules with Extreme Bending Strength. Science 2013, 339, 1298–1302. [Google Scholar] [CrossRef]
- Cha, J.N.; Stucky, G.D.; Morse, D.E.; Deming, T.J. Biomimetic Synthesis of Ordered Silica Structures Mediated by Block Copolypeptides. Nature 2000, 403, 289–292. [Google Scholar] [CrossRef]
- Adamson, D.H.; Dabbs, D.M.; Pacheco, C.R.; Giotto, M.V.; Morse, D.E.; Aksay, I.A. Non-Peptide Polymeric Silicatein α Mimic for Neutral PH Catalysis in the Formation of Silica. Macromolecules 2007, 40, 5710–5717. [Google Scholar] [CrossRef]
- Robinson, D.B.; Rognlien, J.L.; Bauer, C.A.; Simmons, B.A. Dependence of Amine-Accelerated Silicate Condensation on Amine Structure. J. Mater. Chem. 2007, 17, 2113–2119. [Google Scholar] [CrossRef]
- Kisailus, D.; Najarian, M.; Weaver, J.C.; Morse, D.E. Functionalized Gold Nanoparticles Mimic Catalytic Activity of a Polysiloxane-Synthesizing Enzyme. Adv. Mater. 2005, 17, 1234–1239. [Google Scholar] [CrossRef]
- Roth, K.M.; Zhou, Y.; Yang, W.; Morse, D.E. Bifunctional Small Molecules Are Biomimetic Catalysts for Silica Synthesis at Neutral PH. J. Am. Chem. Soc. 2005, 127, 325–330. [Google Scholar] [CrossRef]
- Luckarift, H.R.; Spain, J.C.; Naik, R.R.; Stone, M.O. Enzyme Immobilization in a Biomimetic Silica Support. Nat. Biotechnol. 2004, 22, 211–213. [Google Scholar] [CrossRef]
- Corma, A.; Díaz-Cabañas, M.J.; Moliner, M.; Rodríguez, G. Synthesis of Micro- and Mesoporous Molecular Sieves at Room Temperature and Neutral PH Catalyzed by Functional Analogues of Silicatein. Chem. Commun. 2006, 3137–3139. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Böhm, C.F.; Harris, J.; Schodder, P.I.; Wolf, S.E. Bioinspired Materials: From Living Systems to New Concepts in Materials Chemistry. Materials 2019, 12, 2117. https://doi.org/10.3390/ma12132117
Böhm CF, Harris J, Schodder PI, Wolf SE. Bioinspired Materials: From Living Systems to New Concepts in Materials Chemistry. Materials. 2019; 12(13):2117. https://doi.org/10.3390/ma12132117
Chicago/Turabian StyleBöhm, Corinna F., Joe Harris, Philipp I. Schodder, and Stephan E. Wolf. 2019. "Bioinspired Materials: From Living Systems to New Concepts in Materials Chemistry" Materials 12, no. 13: 2117. https://doi.org/10.3390/ma12132117
APA StyleBöhm, C. F., Harris, J., Schodder, P. I., & Wolf, S. E. (2019). Bioinspired Materials: From Living Systems to New Concepts in Materials Chemistry. Materials, 12(13), 2117. https://doi.org/10.3390/ma12132117