A Novel Role of Ascorbic Acid in Anti-Inflammatory Pathway and ROS Generation in HEMA Treated Dental Pulp Stem Cells

 ,
, 
 ,
,  ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Ethic Statement
2.2. Cell Culture Establishment
2.3. Cell Culture Establishment
2.4. Study Design
- -
- Untreated hDPSCs, utilized as negative control (CTRL);
- -
- hDPSCs treated for 24 h with 2 mM HEMA (HEMA);
- -
- hDPSCs treated for 24 h with 50 µg mL−1 ascorbic acid (AS);
- -
- hDPSCs co-treated for 24 h with 2 mM HEMA and 50 µgmL−1 ascorbic acid (HEMA + AS);
- -
- hDPSCs co-treated for 24 h with 2 mM HEMA and 1 mM N-acetyl Cysteine (HEMA + NAC);
2.5. MTT Assay
2.6. Wound Healing Assay
2.7. Immunohistochemistry and Confocal Laser Scanning Microscope (CLSM) Analysis
2.8. Western Blot Analysis
2.9. ROS Measurements
2.10. Cytokines Evaluation
2.11. Statistical Analysis
3. Results
3.1. Characterization of hDPSCs
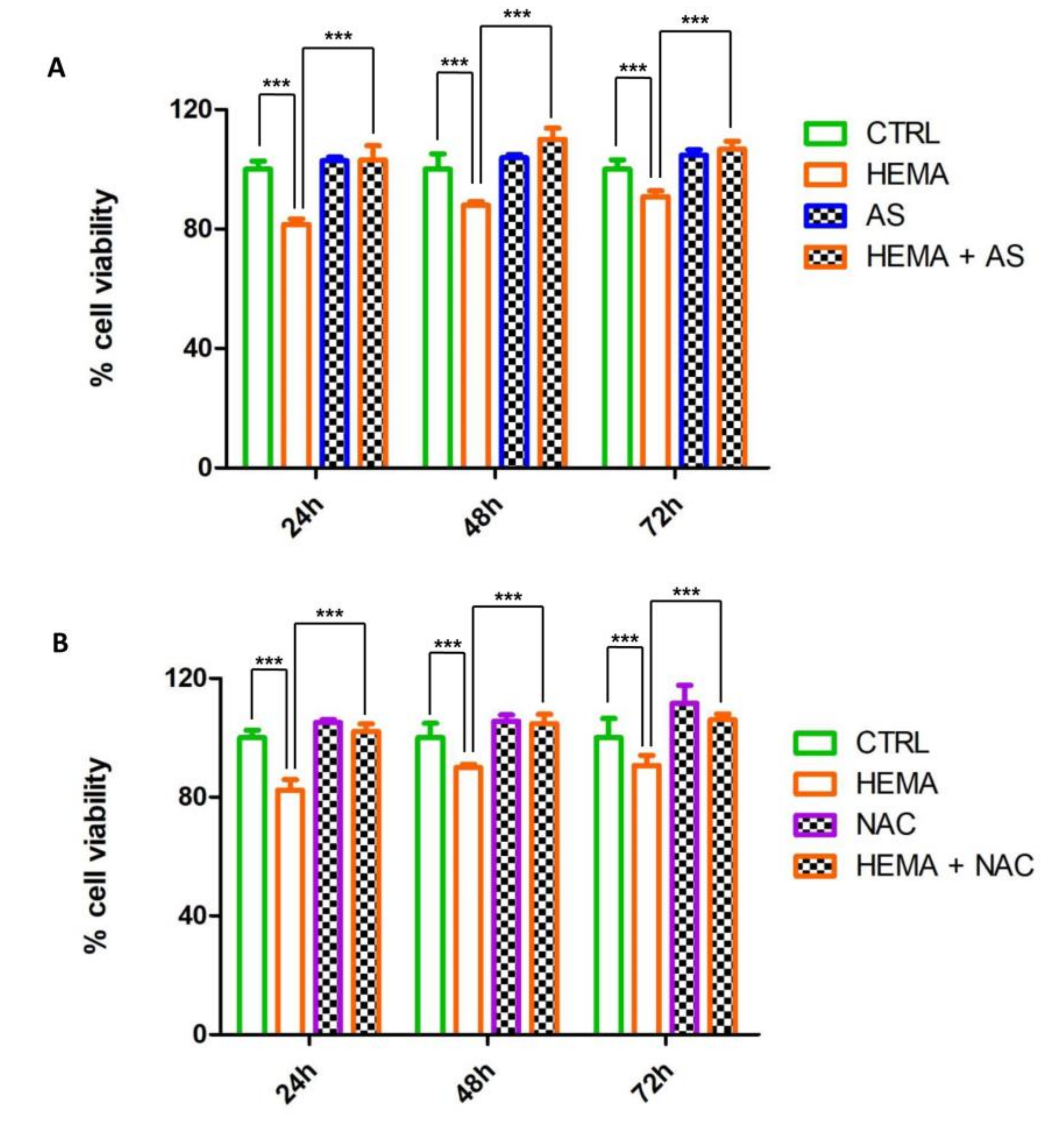
3.2. MTT Cell Viability Assay
3.3. HEMA + AS Treatment Produce Migration of hDPSCs
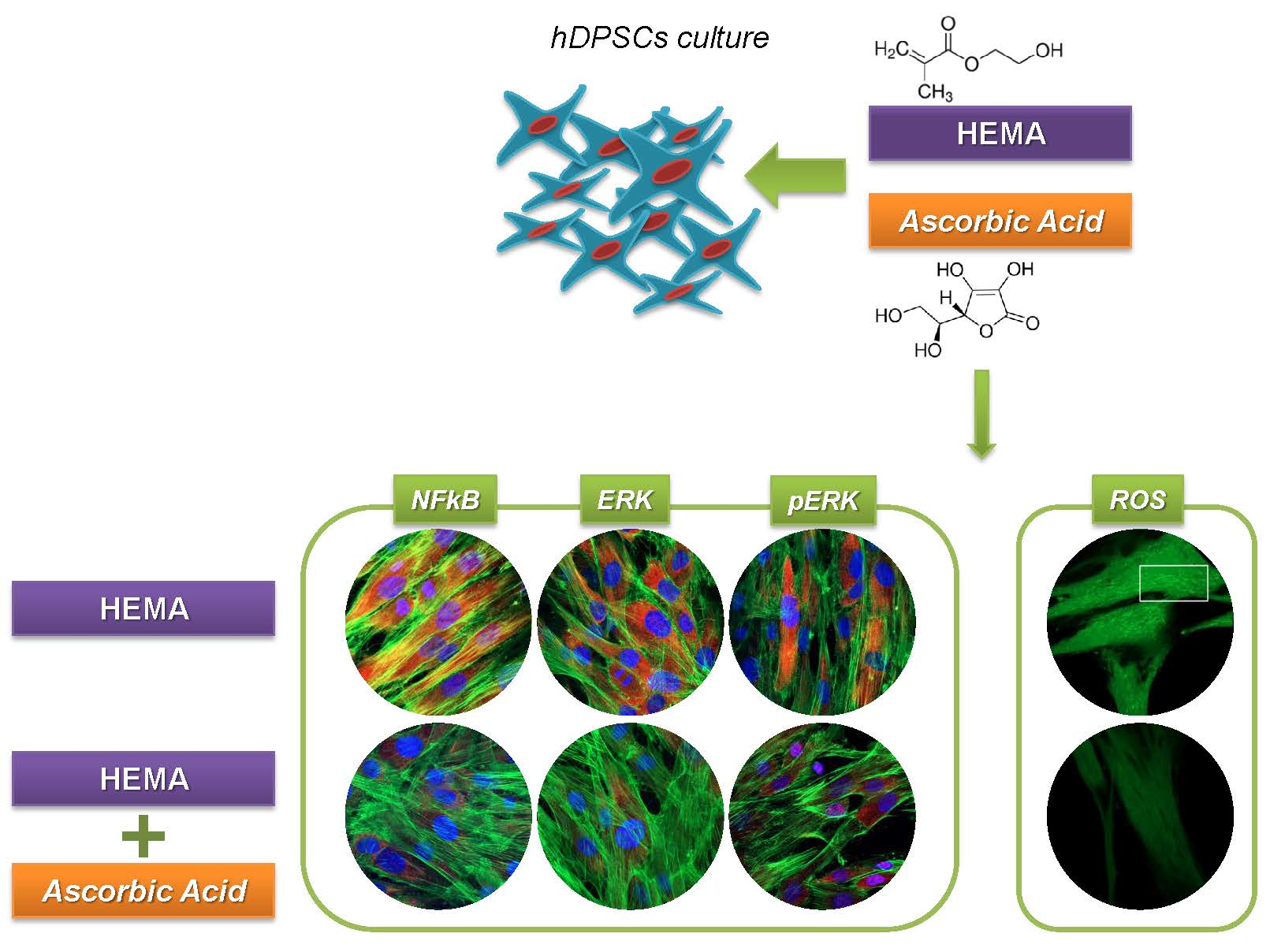
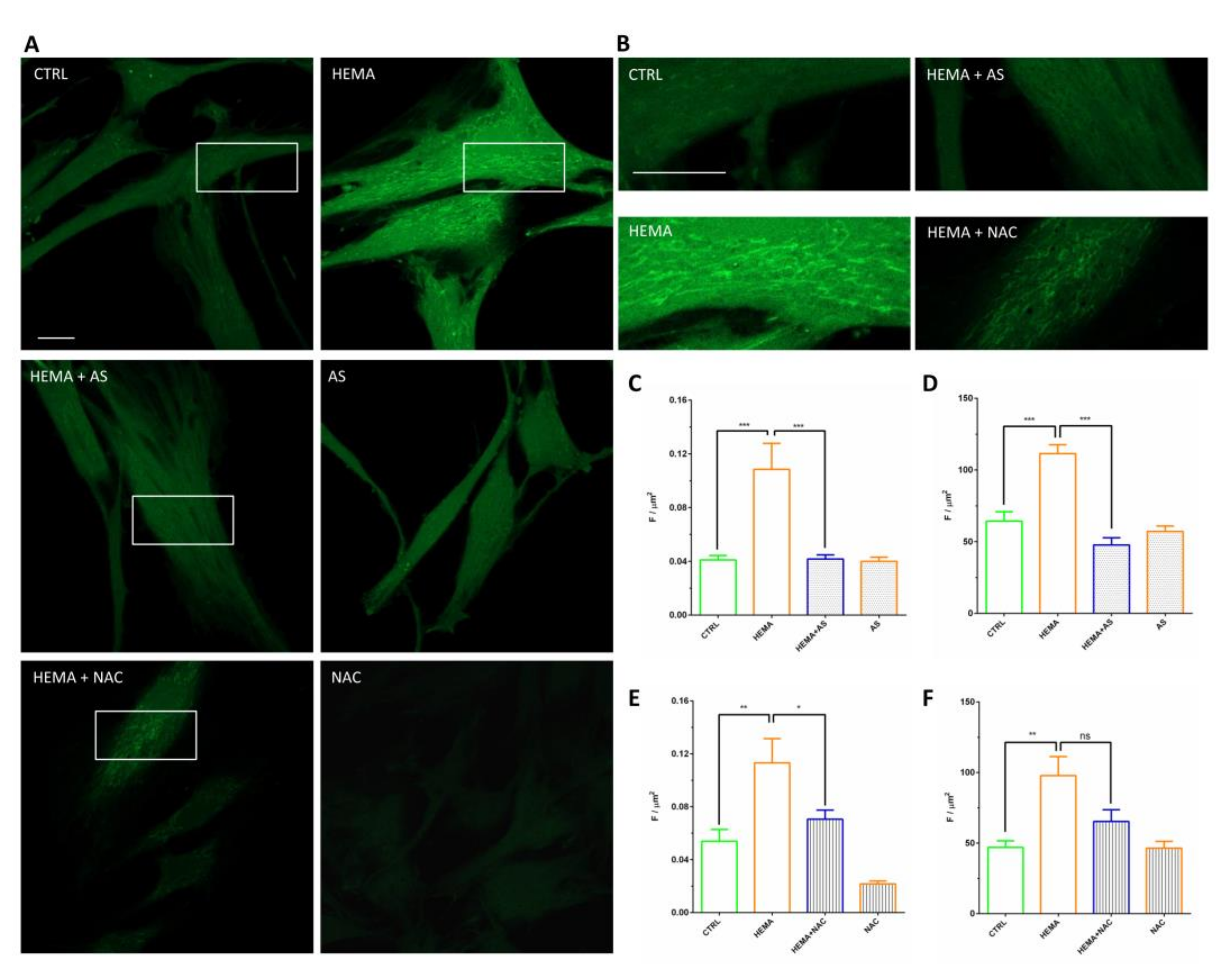
3.4. Signaling Pathway ERK, pERK and NF-kB Analyses
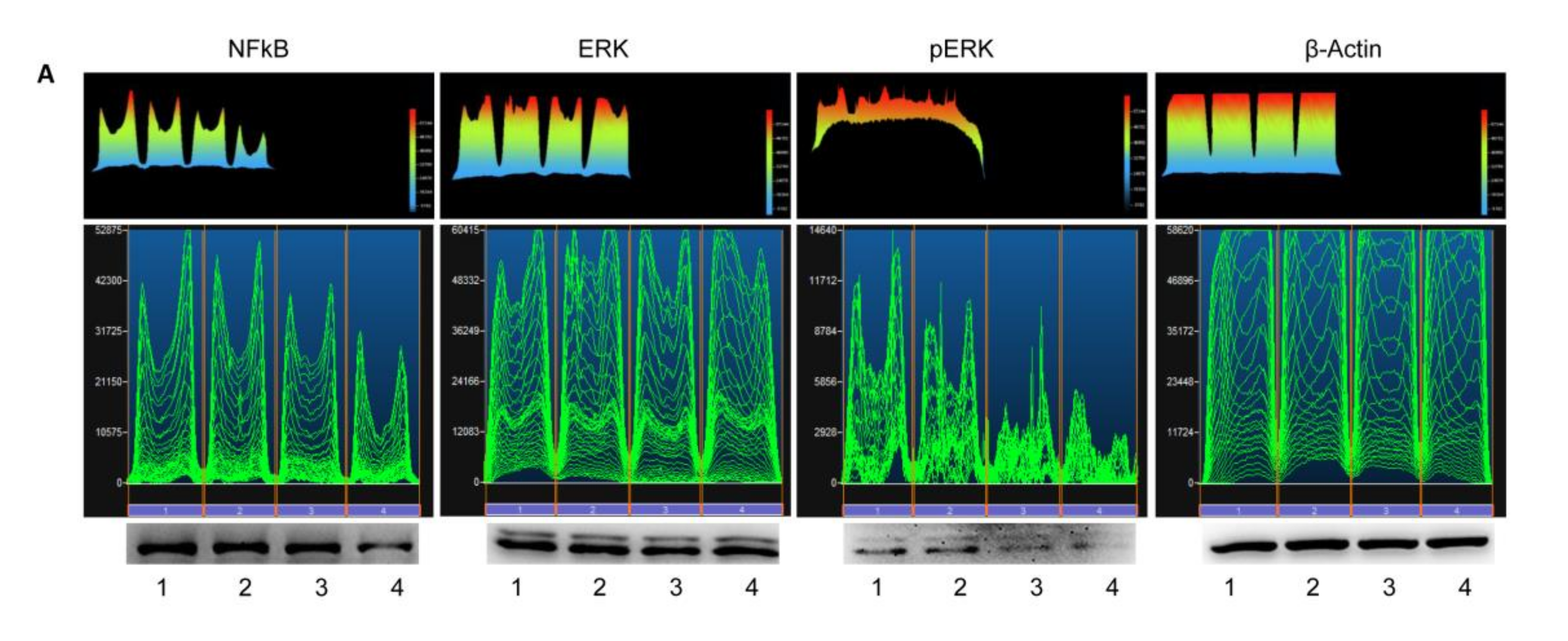
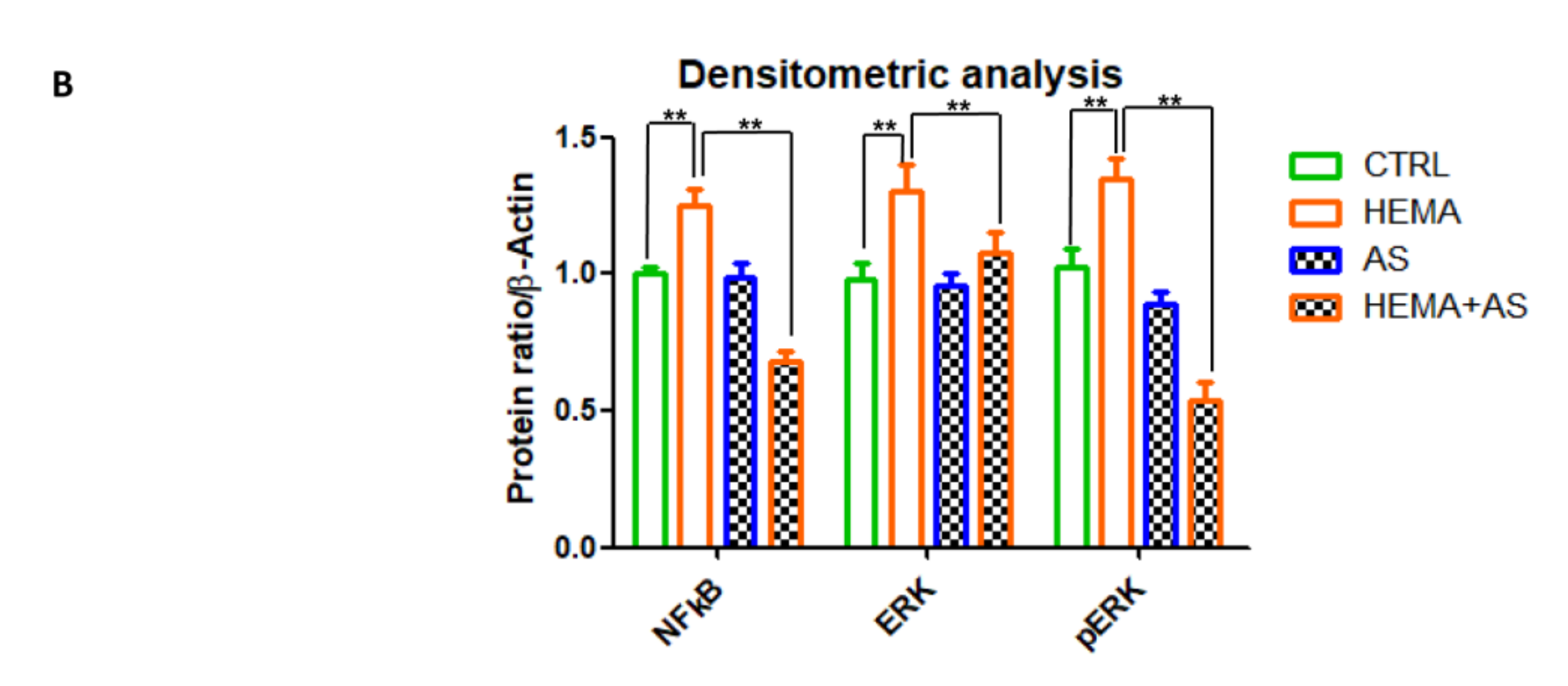
3.5. Western Blot Analyses
3.6. ROS Production
4. Discussion
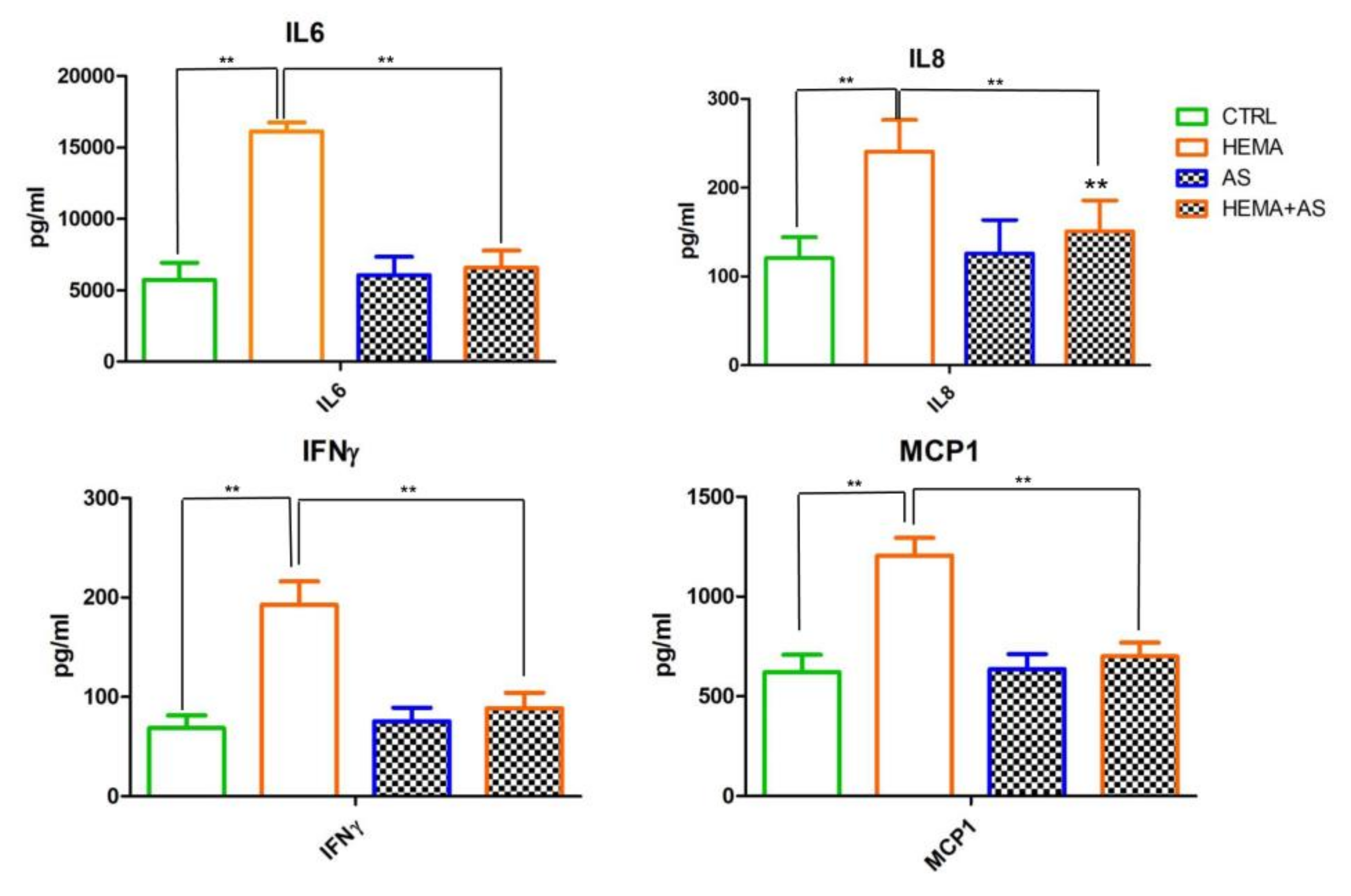
Cytokines’ Release Assessment
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Reichl, F.X.; Durner, J.; Kehe, K.; Manhart, J.; Folwaczny, M.; Kleinsasser, N.; Hume, W.R.; Hickel, R. Toxicokinetic of HEMA in guinea pigs. J. Dent. 2002, 30, 353–358. [Google Scholar] [CrossRef]
- Reichl, F.X.; Seiss, M.; Buters, J.; Behrendt, H.; Hickel, R.; Durner, J. Expression of CYP450-2E1 and formation of 2,3-epoxymethacrylic acid (2,3-EMA) in human oral cells exposed to dental materials. Dent. Mater. 2010, 26, 1151–1156. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Reichl, F.X.; Ilie, N.; Shi, J.W.; Dhein, J.; Hickel, R.; Hogg, C. Antioxidants as a novel dental resin-composite component: Effect on elution and degree of conversion. Dent. Mater. 2019, 35, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Di Nisio, C.; de Colli, M.; di Giacomo, V.; Rapino, M.; di Valerio, V.; Marconi, G.D.; Gallorini, M.; Di Giulio, M.; Cataldi, A.; Zara, S. A dual role for beta1 integrin in an in vitro Streptococcus mitis/human gingival fibroblasts co-culture model in response to TEGDMA. Int. Endod. J. 2015, 48, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Krifka, S.; Petzel, C.; Bolay, C.; Hiller, K.A.; Spagnuolo, G.; Schmalz, G.; Schweikl, H. Activation of stress-regulated transcription factors by triethylene glycol dimethacrylate monomer. Biomaterials 2011, 32, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Pagano, S.; Coniglio, M.; Valenti, C.; Negri, P.; Lombardo, G.; Costanzi, E.; Cianetti, S.; Montaseri, A.; Marinucci, L. Biological effects of resin monomers on oral cell populations: Descriptive analysis of literature. Eur. J. Paediatr. Dent. 2019, 20, 224–232. [Google Scholar] [CrossRef]
- Geurtsen, W.; Lehmann, F.; Spahl, W.; Leyhausen, G. Cytotoxicity of 35 dental resin composite monomers/additives in permanent 3T3 and three human primary fibroblast cultures. J. BioMed. Mater. Res. 1998, 41, 474–480. [Google Scholar] [CrossRef]
- Falconi, M.; Teti, G.; Zago, M.; Pelotti, S.; Breschi, L.; Mazzotti, G. Effects of HEMA on type I collagen protein in human gingival fibroblasts. Cell Biol. Toxicol. 2007, 23, 313–322. [Google Scholar] [CrossRef]
- Spagnuolo, G.; Mauro, C.; Leonardi, A.; Santillo, M.; Paterno, R.; Schweikl, H.; Avvedimento, E.V.; Rengo, S. NF-kappaB protection against apoptosis induced by HEMA. J. Dent. Res. 2004, 83, 837–842. [Google Scholar] [CrossRef]
- Nocca, G.; D’Anto, V.; Desiderio, C.; Rossetti, D.V.; Valletta, R.; Baquala, A.M.; Schweikl, H.; Lupi, A.; Rengo, S.; Spagnuolo, G. N-acetyl cysteine directed detoxification of 2-hydroxyethyl methacrylate by adduct formation. Biomaterials 2010, 31, 2508–2516. [Google Scholar] [CrossRef]
- Krifka, S.; Hiller, K.A.; Bolay, C.; Petzel, C.; Spagnuolo, G.; Reichl, F.X.; Schmalz, G.; Schweikl, H. Function of MAPK and downstream transcription factors in monomer-induced apoptosis. Biomaterials 2012, 33, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Spencer, P.; Ye, Q.; Park, J.; Topp, E.M.; Misra, A.; Marangos, O.; Wang, Y.; Bohaty, B.S.; Singh, V.; Sene, F.; et al. Adhesive/Dentin interface: The weak link in the composite restoration. Ann. BioMed. Eng. 2010, 38, 1989–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebling, J.; Giro, E.M.; Costa, C.A. Biocompatibility of an adhesive system applied to exposed human dental pulp. J. Endod. 1999, 25, 676–682. [Google Scholar] [CrossRef]
- Kiba, H.; Hayakawa, T.; Nakanuma, K.; Yamazaki, M.; Yamamoto, H. Pulpal reactions to two experimental bonding systems for pulp capping procedures. J. Oral Sci. 2000, 42, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Hanks, C.T.; Strawn, S.E.; Wataha, J.C.; Craig, R.G. Cytotoxic effects of resin components on cultured mammalian fibroblasts. J. Dent. Res. 1991, 70, 1450–1455. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Saxena, P.; Pant, V.A.; Pant, A.B. Release and toxicity of dental resin composite. Toxicol. Int. 2012, 19, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Sinjari, B.; Pizzicannella, J.; D’Aurora, M.; Zappacosta, R.; Gatta, V.; Fontana, A.; Trubiani, O.; Diomede, F. Curcumin/Liposome Nanotechnology as Delivery Platform for Anti-inflammatory Activities via NFkB/ERK/pERK Pathway in Human Dental Pulp Treated With 2-HydroxyEthyl MethAcrylate (HEMA). Front. Physiol. 2019, 10, 633. [Google Scholar] [CrossRef]
- Zara, S.; De Colli, M.; Rapino, M.; Di Valerio, V.; Marconi, G.D.; Cataldi, A.; Macchi, V.; De Caro, R.; Porzionato, A. NF-kappaB involvement in hyperoxia-induced myocardial damage in newborn rat hearts. Histochem. Cell Biol. 2013, 140, 575–583. [Google Scholar] [CrossRef]
- Diomede, F.; Tripodi, D.; Trubiani, O.; Pizzicannella, J. HEMA Effects on Autophagy Mechanism in Human Dental Pulp Stem Cells. Materials 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Starkov, A.A. The role of mitochondria in reactive oxygen species metabolism and signaling. Ann. N. Y. Acad. Sci. 2008, 1147, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Nita, M.; Grzybowski, A. The Role of the Reactive Oxygen Species and Oxidative Stress in the Pathomechanism of the Age-Related Ocular Diseases and Other Pathologies of the Anterior and Posterior Eye Segments in Adults. Oxid. Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irani, K. Oxidant signaling in vascular cell growth, death, and survival—A review of the roles of reactive oxygen species in smooth muscle and endothelial cell mitogenic and apoptotic signaling. Circ. Res. 2000, 87, 179–183. [Google Scholar] [CrossRef] [PubMed]
- De Colli, M.; Zara, S.; di Giacomo, V.; Patruno, A.; Marconi, G.D.; Gallorini, M.; Zizzari, V.L.; Tete, G.; Cataldi, A. Nitric oxide-mediated cytotoxic effect induced by zoledronic acid treatment on human gingival fibroblasts. Clin. Oral Investig. 2015, 19, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Mallick, I.H.; Yang, W.; Winslet, M.C.; Seifalian, A.M. Ischemia-reperfusion injury of the intestine and protective strategies against injury. Dig. Dis. Sci. 2004, 49, 1359–1377. [Google Scholar] [CrossRef] [PubMed]
- Deavall, D.G.; Martin, E.A.; Horner, J.M.; Roberts, R. Drug-induced oxidative stress and toxicity. J. Toxicol. 2012, 2012, 645460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roessner, A.; Kuester, D.; Malfertheiner, P.; Schneider-Stock, R. Oxidative stress in ulcerative colitis-associated carcinogenesis. Pathol. Res. Pract. 2008, 204, 511–524. [Google Scholar] [CrossRef]
- Halliwell, B.; Cross, C.E. Oxygen-Derived Species—Their Relation to Human-Disease and Environmental-Stress. Envion. Health Perspect. 1994, 102, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Schweikl, H.; Spagnuolo, G.; Schmalz, G. Genetic and cellular toxicology of dental resin monomers. J. Dent. Res. 2006, 85, 870–877. [Google Scholar] [CrossRef]
- Chang, H.H.; Guo, M.K.; Kasten, F.H.; Chang, M.C.; Huang, G.F.; Wang, Y.L.; Wang, R.S.; Jeng, J.H. Stimulation of glutathione depletion, ROS production and cell cycle arrest of dental pulp cells and gingival epithelial cells by HEMA. Biomaterials 2005, 26, 745–753. [Google Scholar] [CrossRef]
- Schweikl, H.; Hartmann, A.; Hiller, K.A.; Spagnuolo, G.; Bolay, C.; Brockhoff, G.; Schmalz, G. Inhibition of TEGDMA and HEMA-induced genotoxicity and cell cycle arrest by N-acetylcysteine. Dent. Mater. 2007, 23, 688–695. [Google Scholar] [CrossRef]
- Eckhardt, A.; Harorli, T.; Limtanyakul, J.; Hiller, K.A.; Bosl, C.; Bolay, C.; Reichl, F.X.; Schmalz, G.; Schweikl, H. Inhibition of cytokine and surface antigen expression in LPS-stimulated murine macrophages by triethylene glycol dimethacrylate. Biomaterials 2009, 30, 1665–1674. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, M. James Lind’s Treatise of the Scurvy (1753). Postgrad. Med. J. 2002, 78, 695–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarena, V.; Wang, G. The epigenetic role of vitamin C in health and disease. Cell. Mol. Life Sci. 2016, 73, 1645–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, I.B. Evolution and the biosynthesis of ascorbic acid. Science 1973, 182, 1271–1272. [Google Scholar] [CrossRef]
- Nishikimi, M.; Fukuyama, R.; Minoshima, S.; Shimizu, N.; Yagi, K. Cloning and chromosomal mapping of the human nonfunctional gene for L-gulono-gamma-lactone oxidase, the enzyme for L-ascorbic acid biosynthesis missing in man. J. Biol. Chem. 1994, 269, 13685–13688. [Google Scholar]
- Drouin, G.; Godin, J.R.; Page, B. The genetics of vitamin C loss in vertebrates. Curr. Genomics 2011, 12, 371–378. [Google Scholar] [CrossRef]
- Blanchard, J.; Tozer, T.N.; Rowland, M. Pharmacokinetic perspectives on megadoses of ascorbic acid. Am. J. Clin. Nutr. 1997, 66, 1165–1171. [Google Scholar] [CrossRef] [Green Version]
- Vitetta, L.; Sali, A.; Paspaliaris, B.; Reavley, N.J. Megadose vitamin C in treatment of the common cold: A randomised controlled trial. Med. J. Aust. 2002, 176, 298–299. [Google Scholar] [CrossRef]
- Chen, J.; Lan, J.; Liu, D.; Backman, L.J.; Zhang, W.; Zhou, Q.; Danielson, P. Ascorbic Acid Promotes the Stemness of Corneal Epithelial Stem/Progenitor Cells and Accelerates Epithelial Wound Healing in the Cornea. Stem Cells Transl. Med. 2017, 6, 1356–1365. [Google Scholar] [CrossRef]
- Sai, K.K.S.; Bashetti, N.; Chen, X.F.; Norman, S.; Hines, J.W.; Meka, O.; Kumar, J.V.S.; Devanathan, S.; Deep, G.; Furdui, C.M.; et al. Initial biological evaluations of F-18-KS1, a novel ascorbate derivative to image oxidative stress in cancer. EJNMMI Res. 2019, 9. [Google Scholar] [CrossRef]
- Le Lay, S.; Simard, G.; Martinez, M.C.; Andriantsitohaina, R. Oxidative stress and metabolic pathologies: From an adipocentric point of view. Oxidative Med. Cell. Longev. 2014, 2014, 908539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paduano, F.; Marrelli, M.; Amantea, M.; Rengo, C.; Rengo, S.; Goldberg, M.; Spagnuolo, G.; Tatullo, M. Adipose Tissue as a Strategic Source of Mesenchymal Stem Cells in Bone Regeneration: A Topical Review on the Most Promising Craniomaxillofacial Applications. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugliandolo, A.; Diomede, F.; Cardelli, P.; Bramanti, A.; Scionti, D.; Bramanti, P.; Trubiani, O.; Mazzon, E. Transcriptomic analysis of gingival mesenchymal stem cells cultured on 3D bioprinted scaffold: A promising strategy for neuroregeneration. J. Biomed. Mater. Res. Part A 2018, 106, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Zini, N.; Pizzicannella, J.; Merciaro, I.; Pizzicannella, G.; D’Orazio, M.; Piattelli, A.; Trubiani, O. 5-Aza Exposure Improves Reprogramming Process Through Embryoid Body Formation in Human Gingival Stem Cells. Front. Genet. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.F.X.B.; Mazzon, E.; Trubiani, O. Biotherapeutic Effect of Gingival Stem Cells Conditioned Medium in Bone Tissue Restoration. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Ballerini, P.; Diomede, F.; Petragnani, N.; Cicchitti, S.; Merciaro, I.; Cavalcanti, M.F.X.B.; Trubiani, O. Conditioned medium from relapsing-remitting multiple sclerosis patients reduces the expression and release of inflammatory cytokines induced by LPS-gingivalis in THP-1 and MO3.13 cell lines. Cytokine 2017, 96, 261–272. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Cavalcanti, M.; Trubiani, O.; Diomede, F. MicroRNA 210 Mediates VEGF Upregulation in Human Periodontal Ligament Stem Cells Cultured on 3DHydroxyapatite Ceramic Scaffold. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.H.; Park, H.C.; Zhu, T.; Yang, H.C. Inhibition of odontogenic differentiation of human dental pulp cells by dental resin monomers. Biomater. Res. 2015, 19, 8. [Google Scholar] [CrossRef] [Green Version]
- Debeljak Martacic, J.; Borozan, S.; Radovanovic, A.; Popadic, D.; Mojsilovic, S.; Vucic, V.; Todorovic, V.; Kovacevic Filipovic, M. N-Acetyl-L-cysteine enhances ex-vivo amplification of deciduous teeth dental pulp stem cells. Arch. Oral Biol. 2016, 70, 32–38. [Google Scholar] [CrossRef]
- Paranjpe, A.; Cacalano, N.A.; Hume, W.R.; Jewett, A. N-acetylcysteine protects dental pulp stromal cells from HEMA-induced apoptosis by inducing differentiation of the cells. Free Radic. Biol. Med. 2007, 43, 1394–1408. [Google Scholar] [CrossRef] [Green Version]
- Cavalcanti, M.F.X.B.; Maria, D.A.; de Isla, N.; Leal, E.C.P.; Joensen, J.; Bjordal, J.M.; Lopes-Martins, R.A.M.B.; Diomede, F.; Trubiani, O.; Frigo, L. Evaluation of the Proliferative Effects Induced by Low-Level Laser Therapy in Bone Marrow Stem Cell Culture. Photomed. Laser Surg. 2015, 33, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Ebihara, N.; Shima, N.; Kimoto, M.; Funaki, T.; Yokoo, S.; Murakami, A.; Yamagami, S. Adhesion, migration, and proliferation of cultured human corneal endothelial cells by laminin-5. Investig. Ophthalmol. Vis. Sci. 2011, 52, 679–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diomede, F.; D’Aurora, M.; Gugliandolo, A.; Merciaro, I.; Ettorre, V.; Bramanti, A.; Piattelli, A.; Gatta, V.; Mazzon, E.; Fontana, A.; et al. A novel role in skeletal segment regeneration of extracellular vesicles released from periodontal-ligament stem cells. Int. J. Nanomed. 2018, 13, 3805–3825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzatenta, A.; Marconi, G.D.; Macchi, V.; Porzionato, A.; Cataldi, A.; Di Giulio, C.; Pokorski, M. Coexpression of Galanin and Nestin in the Chemoreceptor Cells of the Human Carotid Body. Adv. Exp. Med. Biol. 2016, 885, 77–82. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Rabozzi, R.; Trubiani, O.; Di Giammarco, G. Histidine-Tryptophan-Ketoglutarate Solution Helps to Preserve Endothelial Integrity of Saphenous Vein: An Immunohistochemical and Ultrastructural Analysis. J. Biol. Regul. Homeost. Agents 2011, 25, 93–99. [Google Scholar]
- Di Giulio, C.; Marconi, G.D.; Zara, S.; Di Tano, A.; Porzionato, A.; Pokorski, M.; Cataldi, A.; Mazzatenta, A. Selective Expression of Galanin in Neuronal-Like Cells of the Human Carotid Body. Adv. Exp. Med. Biol. 2015, 860, 315–323. [Google Scholar] [CrossRef]
- Diomede, F.; Merciaro, I.; Martinotti, S.; Cavalcanti, M.F.X.B.; Caputi, S.; Mazzon, E.; Trubiani, O. miR-2861 Is Involved in Osteogenic Commitment of Human Periodontal Ligament Stem Cells Grown onto 3d Scaffold. J. Biol. Regul. Homeost. Agents 2016, 30, 1009–1018. [Google Scholar]
- Giacoppo, S.; Thangavelu, S.R.; Diomede, F.; Bramanti, P.; Conti, P.; Trubiani, O.; Mazzon, E. Anti-inflammatory effects of hypoxia-preconditioned human periodontal ligament cell secretome in an experimental model of multiple sclerosis: A key role of IL-37. FASEB J. 2017, 31, 5592–5608. [Google Scholar] [CrossRef] [Green Version]
- Mammana, S.; Gugliandolo, A.; Cavalli, E.; Diomede, F.; Iori, R.; Zappacosta, R.; Bramanti, P.; Conti, P.; Fontana, A.; Pizzicannella, J.; et al. Human gingival mesenchymal stem cells pretreated with vesicular moringin nanostructures as a new therapeutic approach in a mouse model of spinal cord injury. J. Tissue Eng. Regen. Med. 2019, 13, 1109–1121. [Google Scholar] [CrossRef] [Green Version]
- Mazzatenta, A.; Marconi, G.D.; Zara, S.; Cataldi, A.; Porzionato, A.; Di Giulio, C. In the carotid body, galanin is a signal for neurogenesis in young, and for neurodegeneration in the old and in drug-addicted subjects. Front. Physiol. 2014, 5, 427. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadehgharib, S.; Ostberg, A.K.; Dahlgren, U. Effects of the methacrylate/acrylate monomers HEMA, TEGDMA, DEGDA, and EMA on the immune system. Clin. Exp. Dent. Res. 2017, 3, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carita, A.C.; Fonseca-Santos, B.; Shultz, J.D.; Michniak-Kohn, B.; Chorilli, M.; Leonardi, G.R. Vitamin C: One compound, several uses. Advances for delivery, efficiency and stability. Nanomedicine 2019, 24, 102117. [Google Scholar] [CrossRef] [PubMed]
- Boyce, S.T.; Supp, A.P.; Swope, V.B.; Warden, G.D. Vitamin C regulates keratinocyte viability, epidermal barrier, and basement membrane in vitro, and reduces wound contraction after grafting of cultured skin substitutes. J. Investig. Dermatol. 2002, 118, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Maione-Silva, L.; de Castro, E.G.; Nascimento, T.L.; Cintra, E.R.; Moreira, L.C.; Cintra, B.A.S.; Valadares, M.C.; Lima, E.M. Ascorbic acid encapsulated into negatively charged liposomes exhibits increased skin permeation, retention and enhances collagen synthesis by fibroblasts. Sci. Rep. 2019, 9, 522. [Google Scholar] [CrossRef] [Green Version]
- Larouche, J.; Sheoran, S.; Maruyama, K.; Martino, M.M. Immune Regulation of Skin Wound Healing: Mechanisms and Novel Therapeutic Targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Wang, W.; Yeung, K.W.K. Bone grafts and biomaterials substitutes for bone defect repair: A review. Bioact. Mater. 2017, 2, 224–247. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, X.; Pang, N.; Xiao, L.; Li, Y.; Chen, N.; Ren, M.; Deng, X.; Wu, J. Glycation of vitronectin inhibits VEGF-induced angiogenesis by uncoupling VEGF receptor-2-alphavbeta3 integrin cross-talk. Cell Death Dis. 2015, 6, e1796. [Google Scholar] [CrossRef] [Green Version]
- Pizzicannella, J.; Gugliandolo, A.; Orsini, T.; Fontana, A.; Ventrella, A.; Mazzon, E.; Bramanti, P.; Diomede, F.; Trubiani, O. Engineered Extracellular Vesicles From Human Periodontal-Ligament Stem Cells Increase VEGF/VEGFR2 Expression During Bone Regeneration. Front. Physiol 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Krifka, S.; Spagnuolo, G.; Schmalz, G.; Schweikl, H. A review of adaptive mechanisms in cell responses towards oxidative stress caused by dental resin monomers. Biomaterials 2013, 34, 4555–4563. [Google Scholar] [CrossRef]
- Jiao, Y.; Niu, T.; Liu, H.; Tay, F.R.; Chen, J.H. Protection against HEMA-Induced Mitochondrial Injury In Vitro by Nrf2 Activation. Oxid. Med. Cell. Longev. 2019, 2019, 3501059. [Google Scholar] [CrossRef] [PubMed]
- Pizzicannella, J.; Diomede, F.; Merciaro, I.; Caputi, S.; Tartaro, A.; Guarnieri, S.; Trubiani, O. Endothelial committed oral stem cells as modelling in the relationship between periodontal and cardiovascular disease. J. Cell. Physiol. 2018, 233, 6734–6747. [Google Scholar] [CrossRef] [PubMed]
- Trubiani, O.; Ballerini, P.; Murmura, G.; Pizzicannella, J.; Giuliani, P.; Buccella, S.; Caputi, S. Toll-Like Receptor 4 Expression, Interleukin-6,-8 and Ccl-20 Release, and Nf-Kb Translocation in Human Periodontal Ligament Mesenchymal Stem Cells Stimulated with Lps-P-Gingivalis. Eur. J. Inflamm. 2012, 10, 81–89. [Google Scholar] [CrossRef]
- Heinrich, M.C.; Deloughery, T.G.; Brown, M.A.; Pham, T.Q.; Mccoy, S.L.; Wilkinson, B.M.; Rathbun, R.K.; Hefeneider, S.H.; Bagby, G.C. Il-1 Post-Transcriptionally Regulates the Expression of Il-6 in Human Vascular Endothelial-Cells. Clin. Res. 1990, 38, A150. [Google Scholar]
- De Oliveira, J.F.; Wiener, C.D.; Jansen, K.; Portela, L.V.; Lara, D.R.; Souza, L.D.M.; da Silva, R.A.; Moreira, F.P.; Oses, J.P. Serum levels of interleukins IL-6 and IL-10 in individuals with posttraumatic stress disorder in a population-based sample. Psychiatry Res. 2018, 260, 111–115. [Google Scholar] [CrossRef]
- Zizzari, V.L.; Marconi, G.D.; De Colli, M.; Zara, S.; Zavan, B.; Salini, V.; Fontana, A.; Cataldi, A.; Piattelli, A. In Vitro Behavior of Primary Human Osteoblasts onto Microrough Titanium Surface. Implant Dent. 2015, 24, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Kagan, J.C.; Medzhitov, R. Phosphoinositide-mediated adaptor recruitment controls Toll-like receptor signaling. Cell 2006, 125, 943–955. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, C.; Takeshita, K.; Ueda, J.; Nakanishi, I.; Suzuki, K.T.; Ozawa, T. Reaction of para-hydroxybenzoic acid esters with singlet oxygen in the presence of glutathione produces glutathione conjugates of hydroquinone, potent inducers of oxidative stress. Free Radic. Res. 2006, 40, 233–240. [Google Scholar] [CrossRef]
- Panzarini, S.R.; De Carvalho, A.C.P.; Poi, W.R.; Pedrini, D.; Sonoda, C.K. The use of vitamin C in delayed tooth reimplantation. J. Dent. Res. 2003, 82, 190. [Google Scholar]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diomede, F.; Marconi, G.D.; Guarnieri, S.; D’Attilio, M.; Cavalcanti, M.F.X.B.; Mariggiò, M.A.; Pizzicannella, J.; Trubiani, O. A Novel Role of Ascorbic Acid in Anti-Inflammatory Pathway and ROS Generation in HEMA Treated Dental Pulp Stem Cells. Materials 2020, 13, 130. https://doi.org/10.3390/ma13010130
Diomede F, Marconi GD, Guarnieri S, D’Attilio M, Cavalcanti MFXB, Mariggiò MA, Pizzicannella J, Trubiani O. A Novel Role of Ascorbic Acid in Anti-Inflammatory Pathway and ROS Generation in HEMA Treated Dental Pulp Stem Cells. Materials. 2020; 13(1):130. https://doi.org/10.3390/ma13010130
Chicago/Turabian StyleDiomede, Francesca, Guya Diletta Marconi, Simone Guarnieri, Michele D’Attilio, Marcos F. X. B. Cavalcanti, Maria A. Mariggiò, Jacopo Pizzicannella, and Oriana Trubiani. 2020. "A Novel Role of Ascorbic Acid in Anti-Inflammatory Pathway and ROS Generation in HEMA Treated Dental Pulp Stem Cells" Materials 13, no. 1: 130. https://doi.org/10.3390/ma13010130
APA StyleDiomede, F., Marconi, G. D., Guarnieri, S., D’Attilio, M., Cavalcanti, M. F. X. B., Mariggiò, M. A., Pizzicannella, J., & Trubiani, O. (2020). A Novel Role of Ascorbic Acid in Anti-Inflammatory Pathway and ROS Generation in HEMA Treated Dental Pulp Stem Cells. Materials, 13(1), 130. https://doi.org/10.3390/ma13010130