1. Introduction
Blood is a complex suspension of red blood cells (RBCs), white blood cells (WBCs) and platelets in an aqueous solution, the so-called plasma, containing dissolved proteins [
1]. Although many works consider blood as a Newtonian fluid [
2,
3], in fact, it has a pronounced non-Newtonian character, mostly explained in terms of the ability of RBCs to aggregate/disaggregate, deform and align to flow [
4,
5,
6,
7]. At low shear rates, blood proteins enhance the formation of a complicated network made up of column-like red cell aggregates known as rouleaux, while at almost stasis, the rouleaux forms three-dimensional networks [
8]. At higher shear rates, these structures tend to disintegrate, leading to a state where the red blood cells flow separately. The physical mechanism that drives the creation of rouleaux is not settled yet but two theories emerge as potential candidates, namely the bridging of fibrinogen [
9,
10] and the depletion theory [
11]. This aggregation is a dynamic property of blood [
12] promoting its non-Newtonian nature. The aforementioned behavior necessitates the use of sophisticated rheological models to capture adequately the rheological response of blood.
In the last decades, investigation of hemorheology has stimulated a lot of attention to the complex nature of blood mostly due to the direct relevance of blood rheology to disease detection, treatment, and prevention. A number of diseases exhibit discernable hemorheological changes and vice versa, including sickle cell anemia, which is the most frequent genetic disease in the world, diabetes, hypertension, or heart disease [
13,
14,
15] or connective tissue disease, blood hypercoagulability due to COVID-19 [
16,
17], arteriosclerosis, or diabetic blood [
18,
19,
20]. An indicative example is the coagulation process, which initiates the formation of a fibrin network along with the clumping of red blood cells giving rise to enhanced elastic aspects [
21]. The latter process becomes of significant importance when clotting of blood occurs [
22,
23], and the influence upon tissue is much more severe than when only the blood viscosity, which is another significant hemorheological parameter, is increased. It should be mentioned that during the clotting process, the elastic modulus varies with time by many orders of magnitude especially for a diabetic blood [
24]. As a result, hemorheological measurements can potentially offer an individualized and straightforward method for detecting the presence of such diseases. However, using hemorheology as a diagnostic tool requires both data and accurate constitutive models for healthy and disease states. Of equal importance for advancing the engineering design of medical devices is the ability to simulate in silico the in vivo flow of blood, which requires accurate knowledge of the transient material properties of blood.
Based on our previous discussion, it is perceivable that the rheological characterization of blood can be divided in two categories: steady state analysis and transient analysis. Initially, a significant effort focused on the steady-state shear rheometry, excluding phenomena such as the formation and dissolution of the rouleaux structures, which are placed at differing time scales, and are both flow-dependent. However, steady-state investigations should not be underestimated as they have given an insight into non-Newtonian characteristics and their association with blood flow. Blood possesses a distinct property according to which it flows as fluid if subjected to large enough stresses but behaves as a soft matter if the applied stress is below a critical value, giving rise to the experimentally measured blood yield stress [
25,
26]. The latter is an essential component of its non-Newtonian nature, stemming explicitly from the creation of rouleaux. Its role is most clearly evaluated under steady-state shear flow conditions. Experimental evidence of the exhibition of the yield stress has been provided by many investigations [
6,
25,
27,
28] leading to viscoplastic modeling of blood (VP) [
29]. Additionally, the well-known shear thinning behavior of blood has been described by a plethora of investigations [
30,
31] in various hemodynamical conditions as well as the existence of a critical hematocrit associated with blood yield stress and the transition from non-Newtonian to Newtonian flow in high shear rates [
32].
However, blood possesses also a non-negligible elastic nature, as shown experimentally [
33,
34]. Apparently, this is an inherent property of blood, [
35,
36] and it can be associated with the lipidic bilayer membrane of RBCs and hemorheological variables such as hematocrit and fibrinogen concentration [
37,
38]. Recently, Varchanis et al. [
39] showed that the elastic nature of human blood plasma should not be neglected in regimes of intense shear. Particularly, important contributions to the experimental investigation of the viscoelasticity have been provided by Thurston [
34], who performed oscillatory flow experiments in cylindrical tubes. Blood is also classified as a non-ideal yield-stress material due to its non-negligible elastic effect. Thus, the need to introduce the concept of elasto-visco-plastic (EVP) fluids is clear to describe materials that have elastic and plastic characteristics simultaneously.
Although the findings of the aforementioned investigations are important, the inherent properties of blood are strongly time-dependent as well, and thus, blood is categorized as a thixotropic material. From a rheological perspective, blood is one of the most characteristic examples of a thixotropic fluid and allows for the evaluation of generalized thixotropy models for a unique and biologically relevant case. A thixotropic fluid is a fluid whose material properties (e.g., viscosity) is a function not only of the applied stress but also of the previous history of motion within the fluid [
5,
21,
24,
40,
41]. The thixotropic nature of blood stems from the aggregation/disaggregation of rouleaux which is governed by its own time scales affected by the concentration of plasma proteins [
42] and hematocrit [
9]. In particular, the yield stress in dense, soft colloidal suspensions such as blood is typically attributed to an internal structure that develops, deforms, and decays in a way that depends critically not only on the current flow kinematics but also on its deformation history, thus giving rise to thixotropy [
9,
21,
43,
44]. Some of the first reports were those of Lacombe and Quemada [
21,
45], who focused on the description of these transient blood flow phenomena. One of the prominent experimental investigations is that of Bureau et al. [
46] who systematically obtained hysteresis and step-up curves of pathological and physiological human blood. Here, we invoke the experimental work of McMillan et al. [
47], who performed steady-state shear and rectangular shear-step tests which are capable to probe the elasto-thixotropic response of blood making our model robust and reliable. More recently, Armstrong et al. [
48] and Horner et al. [
49] conducted experiments in steady and transient rheometric flows. Except for the triangular step (hysteresis) experiments, the application of the oscillatory flow is commonly employed to probe the complex properties of blood.
The rheological modeling of blood should consider all the above phenomena to predict a realistic behavior. Therefore, the focus has been shifting towards constitutive equations that incorporate plasticity, elasticity and thixotropy. The model of Owens and coworkers [
50], which has been recently revised [
51,
52,
53], was derived using ideas drawn from polymer network theory accounting for the agglomeration and deagglomeration of erythrocytes in healthy human blood at different shear rates. Although this model was subsequently applied to simple shear flows as well as to steady, oscillatory, and pulsatile flow in rigid vessels [
54], it lacks explicit accounting for yield stress, the most important manifestation of the viscoplastic nature of blood. Additionally, Anand and Rajagopal [
55] used a generalized Oldroyd-B model, which was developed in the context of the general thermodynamic framework of Rajagopal and Srinivasa [
56]. They made use of a tensorial viscoelastic model and therefore do not explicitly take into account the viscoplastic nature of blood, although their work was found to agree with steady-state and transient experiments. Early efforts to investigate the thixotropy of blood were made by Apostolidis et al. [
57], who based on a phenomenological model to incorporate thixotropy and viscoplasticity of blood, employed the Casson model, which is suitable for steady-state flows. They offered a systematic study of the rheology of blood in transient shear flows, based on a single scalar internal structural thixotropic parameter. We note that these models have primarily originated from the need for macroscopic modelling of materials composed by particles where phenomena such as the formation of agglomerates are pronounced [
58,
59]. Later it was suitably modified to exploit recent advances based on the kinematic hardening model of the elastic strain in plasticity theory of Dimitriou et al. [
60]. Although this model was evaluated in transient and steady experiments, it lacks a tensorial form preventing its use in simulating multi-dimensional flows. Another remarkable theoretical investigation was made by Kaliviotis & Yianneskis [
61] who presented a theoretical examination of the hypothesis that the network characteristics of red blood cell influence the mechanical properties of the blood. They developed an energy-rate based model, which incorporated network dynamics used to predict the transient behavior of blood suggesting that network characteristics play a significant role in the configuration of the blood viscosity.
The coupled appearance of several elastic, plastic and thixotropic phenomena and the underlying mechanisms and internal variables in thixotropic elasto-visco-plastic (TEVP) materials necessitates the introduction of a large number of adjustable parameters to describe their rheology. The model proposed by Varchanis et al. [
62] was suitable for highly concentrated suspensions, which exhibit shear thinning, thixotropy, plasticity and viscoelasticity. It is adequately modified in order to have a stress-controlled dynamic configuration of the structure parameter, accompanied by a reduction in fitting parameters. Our model consists of a set of eleven adjustable parameters, contrary to the original model, which in addition incorporates the kinematic hardening mechanism and consequently has extra parameters. We offer a consistent TEVP constitutive model that encompasses the most crucial characteristics of the aforementioned blood properties in order to validate experimental results satisfactorily and provide accurate predictions for steady-state flow regimes in microchannels. Our model is shown to produce results that agree well both with steady state and transient blood flow data [
47]. Apart from the integrated modeling of blood rheological complexity, our implementation is adequate for multi-dimensional simulations due to its tensorial formalism, contrary to recent investigations of blood flow [
57], which do not offer a tensorial form of their model. Another superior advantage is the single mode approach, meaning that the thixotropic effects follow a single time scale. A multimode thixotropic model is directly related to additional computational cost, which in the case of -two or -three-dimensional flows would be prohibitive. The single thixotropic time scale used in the present study, can yield satisfactory results and capture the behavior of blood, given that a proper rheological characterization has been performed. Our model provides an extensive predictive capability in steady and transient testing flow regimes and demonstrates rheological behavior, which is consistent with limited existing experimental data.
2. Constitutive Modelling
In this section, we outline the components of the constitutive model by invoking the work of Varchanis et al. [
62], which has been based on Saramito’s model (SRM) [
63] for EVP materials. Generally speaking, the starting point is the definition of the extra stress tensor
which is composed of a purely Newtonian contribution,
, and a viscoelastic one,
as:
Our model accounts not only for the viscoelastic contribution of the RBCs but also for the viscoelasticity of blood plasma since it has been proved to present a significant non-Newtonian behavior because of the presence of plasma proteins [
39,
64]. Thus, there is no explicit Newtonian contribution
as long as the solvent (plasma) is included in the viscoelastic term (
).
Following the studies of Dimitriou and McKinley [
60], Fraggedakis et al. [
65], Stickel et al. [
66], Dimitriou et al. [
67], Clarion et al. [
68] and Horner et al. [
49], we assume that the total rate of deformation tensor:
is composed of an elastic contribution
and a viscoplastic one denoted by
so that:
The elastic term of the total deformation rate tensor
which can be written as:
Considering the memory effects by introducing the upper-convected time derivative as in the majority of VE models, while
denotes the elastic modulus of the blood. For relatively low shear rates, the elastic aspects are pronounced, and the suspension is acting like a neo-Hookean solid. This can be justified by the appearance of the upper convected derivative into the elastic term. At the same time, for a Hookean solid, the linear relationship between strain and stress would be denoted by the introduction of the material derivative. For any tensor z, the upper convective time derivative is defined as:
The effect of plasticity is introduced via multiplication of distinct functions and contributions given by the expression:
Following the original TEVP model proposed by Varchanis et al. [
62],
incorporates the back stresses which arise due to kinematic hardening (KH). The latter along with isotropic hardening (IH) are mechanisms related to variations in the material structure (e.g., dynamics of microparticles of a suspension, RBC aggregates). Also, they are very well known in the plasticity literature of solid mechanics. Primarily, the KH function is highlighted by flow reversal experiments, and it has been observed to be an inherent property of solids. Consequently, although KH could be a crucial component for a VP constitutive modelling, here we assume that it has negligible contribution, in the absence of flow reversal experiments and due to the belief that blood does not exhibit intense kinematic hardening, so as
.
The von Mises criterion is essential for controlling the transition between the structured (e.g., rouleaux) and the destructured (e.g., individual RBCs) states which is introduced via a continuous max() function for any value of the magnitude of the effective stress
. This criterion is an ellipsoidal representation in stress space when the equal sign holds in the following,
. The effective stress is defined through the deviatoric part of the stress tensor as
, where the deviatoric part of any tensor
is defined as
. The plasticity term is defined in accordance with the SRM model [
63] multiplied with a nonlinear term which is a function of the trace of the viscoelastic stress tensor
, as imposed by the linear Phan-Thien Tanner (PTT) function
. In contrast to the original model, here the PTT model bounds the extensional viscosity by an extra fitting parameter at high strain rates. Using the linear form of PTT function, which has been found to be the most suitable for this VE modeling, we exclude the contribution of shear thinning because this is suitable for exclusively VE consideration as argued by Varchanis et al. [
69]. However, in our model shear thinning is mainly introduced via thixotropic viscosity
. Accordingly, the stress PTT function is defined as:
where
is a fitted parameter. For closure reasons of the model, we need to define equations that govern the evolution of the internal variables. Most of the investigations on thixotropic modelling incorporate a component of the structural statement, which is represented by a non-dimensional structure variable,
λ. This parameter, in our case, reflects the current level of rouleaux formation within the blood sample, which changes over time. For simplicity, it varies in a predetermined range with limits corresponding to the two extreme cases regarding the level of the material structure. More specifically, when
, the blood is assumed to be in a fully structured state (jammed rouleaux), while when
, the blood is supposed to be completely unstructured (individual RBCs deform and flow with blood) and rheologically softer. Firstly, we present the equation governing the thixotropic structural changes within the blood. The time depended evolution of the structure (or rouleaux) parameter is then given by Wei et al. [
59,
70]:
The term
in Equation (8) represents the rebuilding of the structure given by:
which is caused either by Brownian collisions of individual RBCs, scaled with
, or by deformation scaled with
, while both contributions are proportional to the fraction of unstructured blood (e.g., individual RBCs)
. On the contrary,
refers to the breakdown which is assumed to grow proportionally to the shearing and the fraction of structured blood,
λ as:
where the
parameter scales the impact of the viscoplastic rate of deformation on the breaking of rouleaux. Several different representative kinetic equations can be found in the literature and are summarized in the review of [
70], while the «flow parameter»
can be either deformation rate or stress controlled, or a combination of both. Contrary to Varchanis et al. [
62], we have chosen to base the dependency of the level of structure explicitly on the stresses as also adopted by Wei et al. [
59]. Consequently, our model incorporates a stress-controlled form given by:
Note that the rate-controlled (RC) and stress-controlled (SC) forms correspond to two similar, but different models. They make different constitutive assumptions about how flow conditions influence the thixotropic kinetics.
In the absence of kinematic hardening (KH), the material possesses a yield stress that is denoted by
. Although in the original TEVP model [
62], the last parameter depends on the magnitude of the deformation rate through
, here for simplicity, we argue that the blood yield stress is not related to history effects, and thus, the impact of isotropic hardening (IH) on the yield stress is not taken into consideration. This contribution to the yield stress is most easily appreciated, representing the strength of the structural network due to bonds between its components. On the contrary, the plastic viscosity of the material
is a thixotropy-dependent variable. Because of its dependence on
, the current model is classified as nonideal thixotropic material. Finally, the variation of the plastic viscosity
must be specified. Contrary to the simplest linear relationship proposed by Dimitriou et al. [
60], we generalize the dependence of the rheological parameters on the structure variable and propose a simple relationship to include a nonlinearity, as suggested by Geri et al. [
71] for these materials. From a mathematical perspective, we propose an expression able to control this nonlinearity based on the response of the TEVP material to flow. Thus, the dependence of the viscosity on
is defined as:
where
,
are fitting parameters. Because of the bounded value of the structure parameter in the range of 0–1, plastic viscosity will attain values in a predetermined range (
,
] depending on the value of
. The limit
denotes the value of the parameter
when the blood is in a fully structured state (
), while in almost fully unstructured case (
) the plastic viscosity should tend to the high-shear rate viscosity of blood which is
times greater than that of plasma. In this way, the latter rheological property is allowed to be affected by this impact in a different manner. We can define the relaxation time of blood,
, as the ratio of the plastic viscosity to the shear modulus:
where
.
3. Rheological Data Fitting and Calculation of the Model Parameters
This section is dedicated to the fitting of our model to experimental data from rheometric flows. The coupled appearance of several elastic and plastic phenomena and the underlying mechanisms and internal variables in TEVP materials, such as blood, necessitates the introduction of a large number of adjustable parameters to describe their rheology. The available experimental data in the literature are limited to those reported in the works of McMillan et al. [
47], Armstrong et al. [
48] and Bureau et al. [
46], which were restricted to a small range of hemodynamical properties eliminating the opportunity of making a parametric correlation between the model’s features and fundamental macroscopic properties such as fibrinogen or hematocrit. Primarily, we use the experiments conducted by McMillan et al. [
47] for a healthy subject of systemic hematocrit equal to
. The steady-state experiment refers to simple shear flow data providing the shear stress response as a function of the imposed shear rate. Transient experiments are flow curves exported from transient repetitive shearing steps, also known as intermittent steps or rectangular shear steps, with an imposed shear rate equal to
with shearing periods lasting
, while the intermittent non-shearing intervals have a duration of about
. Additionally, we provide the fitting of our model to another subject, and especially we use the steady simple shear, the triangular shear-step and the oscillatory shear data of Donor 1 of the experiments conducted by Armstrong et al. [
48]. To extract realistic parameters for the constitutive model, we adopt a non-linear regression [
69] on experimental data for steady-state and transient experiments.
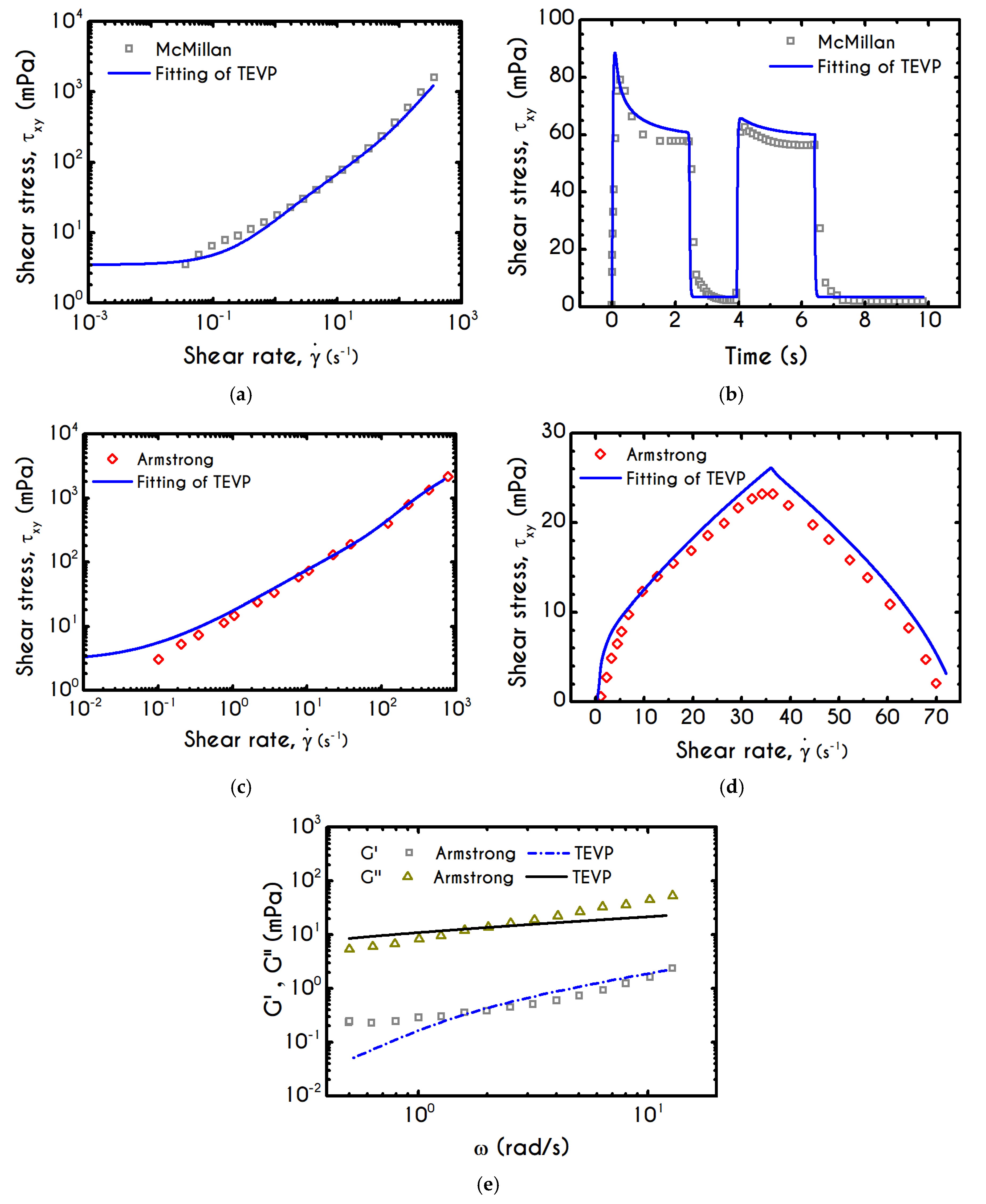
The fitted curves illustrated in
Figure 1, depict an excellent capturing of the major characteristics of both steady shear and transient experiments for both blood samples reported in [
47,
48], while the values of the adjustable parameters are given in
Table 1 for each subject separately.
The extracted parameters of our model for both subjects are in agreement with the theoretical predictions and the experimental measurements given in [
21,
72]. Also, both sets of parameters have the same order of magnitude even though the corresponding subjects have different hematological and aging features. We have to point out that shearing in Equation (8) has a dual role, which simultaneously contributes to the dissolution and reconnection of the structural clusters (linear rouleaux and three-dimensional networks). However, its prominent role is to collapse the microstructure necessitating that
is significantly larger than
. The same pattern is valid for the corresponding exponents where
is expected to be lower than
as well as they must be positively defined including
. Another significant issue regarding the fitting of a model is the impact a small change of the input (adjustable parameters) to the output (stresses or microstructure parameter). To make appropriate estimations of the uncertainty of the model’s parameters, the exact values of the experimental errors should also be provided along with the experimental data. None of the known experimental works [
46,
47,
48] gives these errors, and hence we are not able to provide the admissible ranges of our model’s parameters accurately.
Steady flow properties in simple shear are characterized by the stationary relationship
which can be obtained in both stress-controlled and shear-rate-controlled experiments. McMillan et al. [
47] adopted a rate-controlled technique, which enforces an immediate breakdown of the microstructure and consequently blood begins to fluidize when
. The imposition of a shear-rate that enforces the system to reach a steady state, indicates that stress always exceeds yield stress and the blood is yielded. On the contrary, a flow curve from a stress-controlled rheometer gives a definite interpretation of both yield stress and unyielded region. This means that when applying a stress value, the blood is initially below the yielding point, and thus it is in its solid regime where its behavior is basically elastic, dictating a linear relationship between stress and strain (
). The value above which a clear non-linear behavior between stress and shear rate is observed, indicates the yield stress,
. The convexity in the experimental data of McMillan et al. [
47] is originated from the experimental protocol, and our fit is in accordance with other similar fittings in [
73]. The correct way to obtain the real shear stress – shear rate relationship is to subject the material at a shearing where the shear rate increases initially up to a specific value, and then it decreases to zero. The ‘ramp-down’ section of the flow curve should be used to determine the yield stress of the fluid. Regarding the steady-state shear experiment, the fitting of our model depicts an excellent agreement across the whole range of the shear rates while the predicted yield stress is equal to
.
The blood sample is characterized by a hematocrit equal to 45%, but the fibrinogen value is not provided by McMillan et al. [
47]. As we have already mentioned, blood rheology is significantly affected by the fibrinogen concentration [
25] and consequently it is directly influencing the yield stress magnitude. Morris et al. [
74] have proposed a parametric estimation of blood yield stress based on their experimental data. The experiments covered a wide range of fibrinogen concentrations
and hematocrits
). The empirical expression extracted by a fitting procedure is then given by:
where
is the yield stress in
and
is the fibrinogen concentration in
. Since, the blood sample data used from the current work indicates a yield-stress equal to
, it corresponds to a fibrinogen concentration equal to
. This is an absolutely physiological value consistent with hemorheological characteristics of the blood samples examined by McMillan et al. [
47].
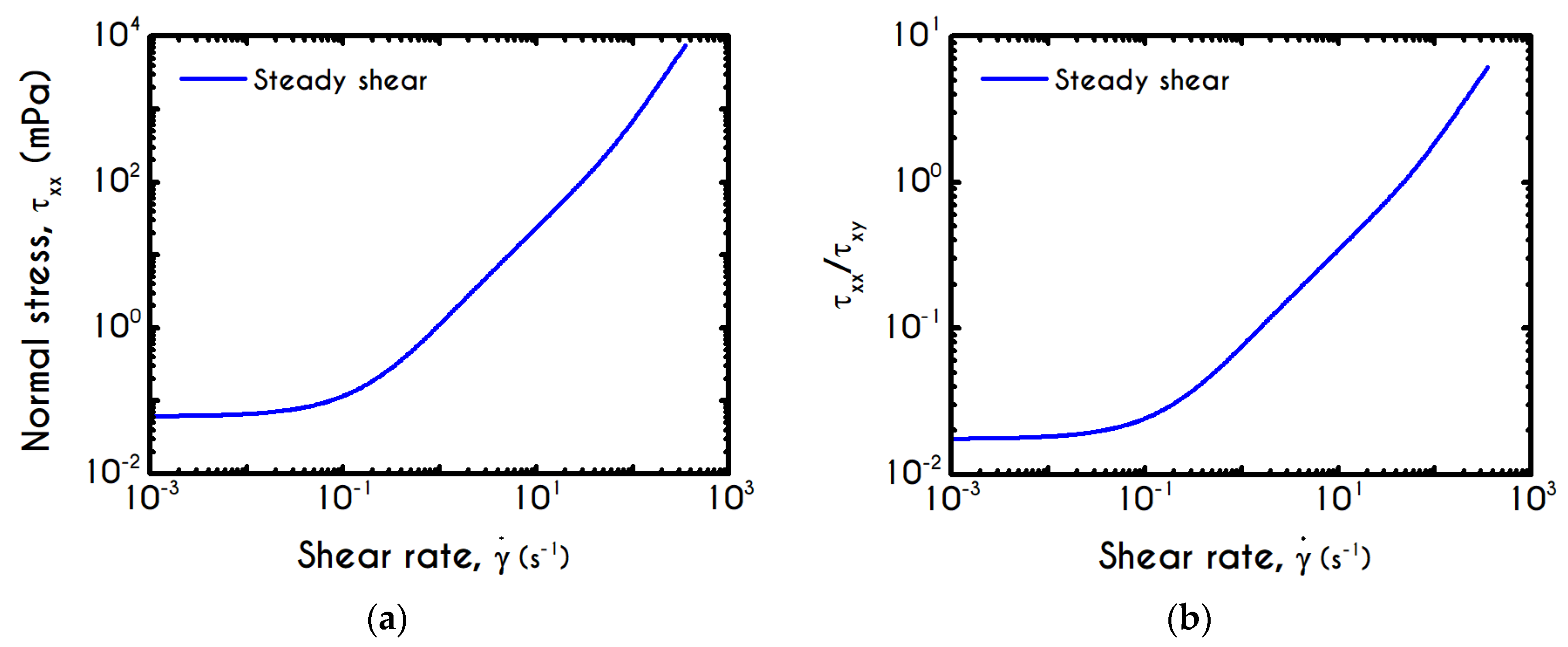
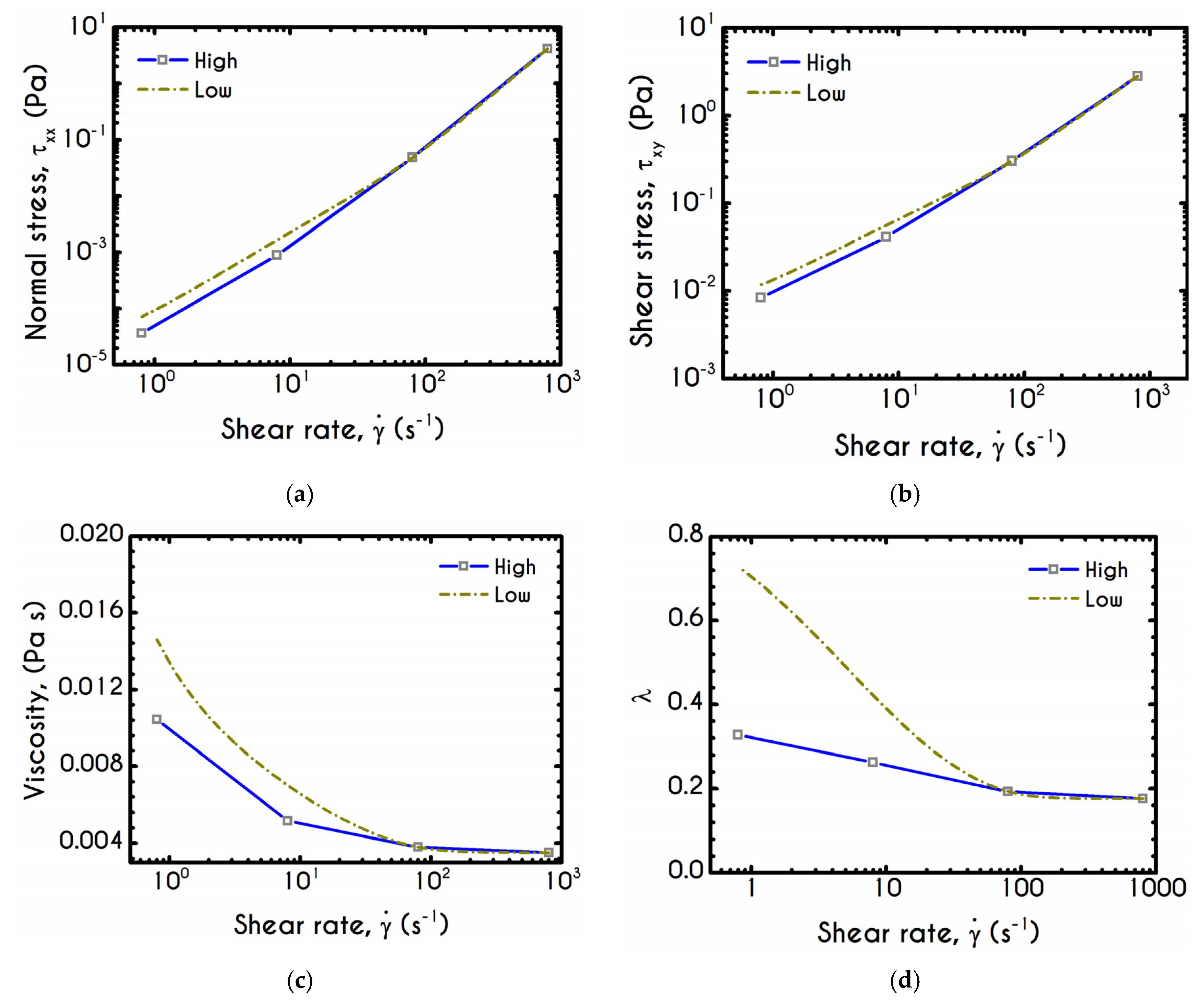
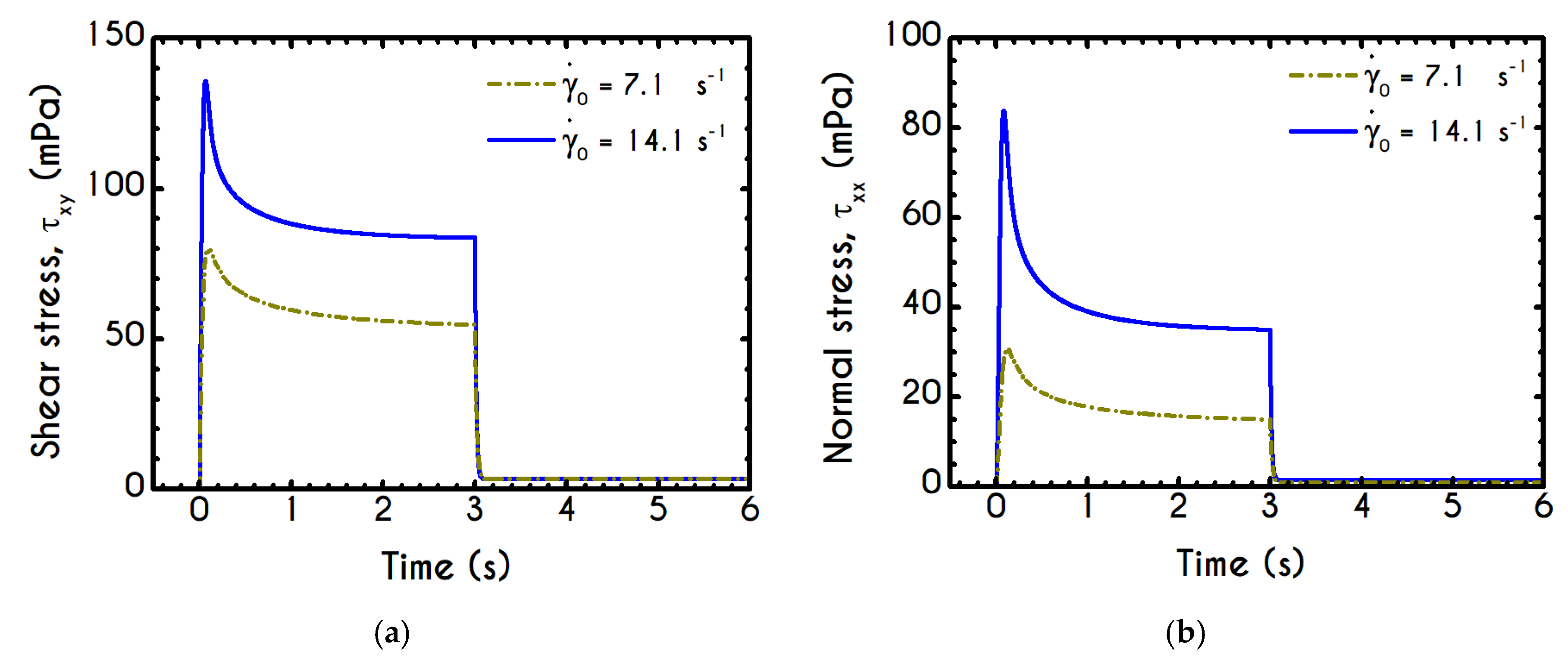
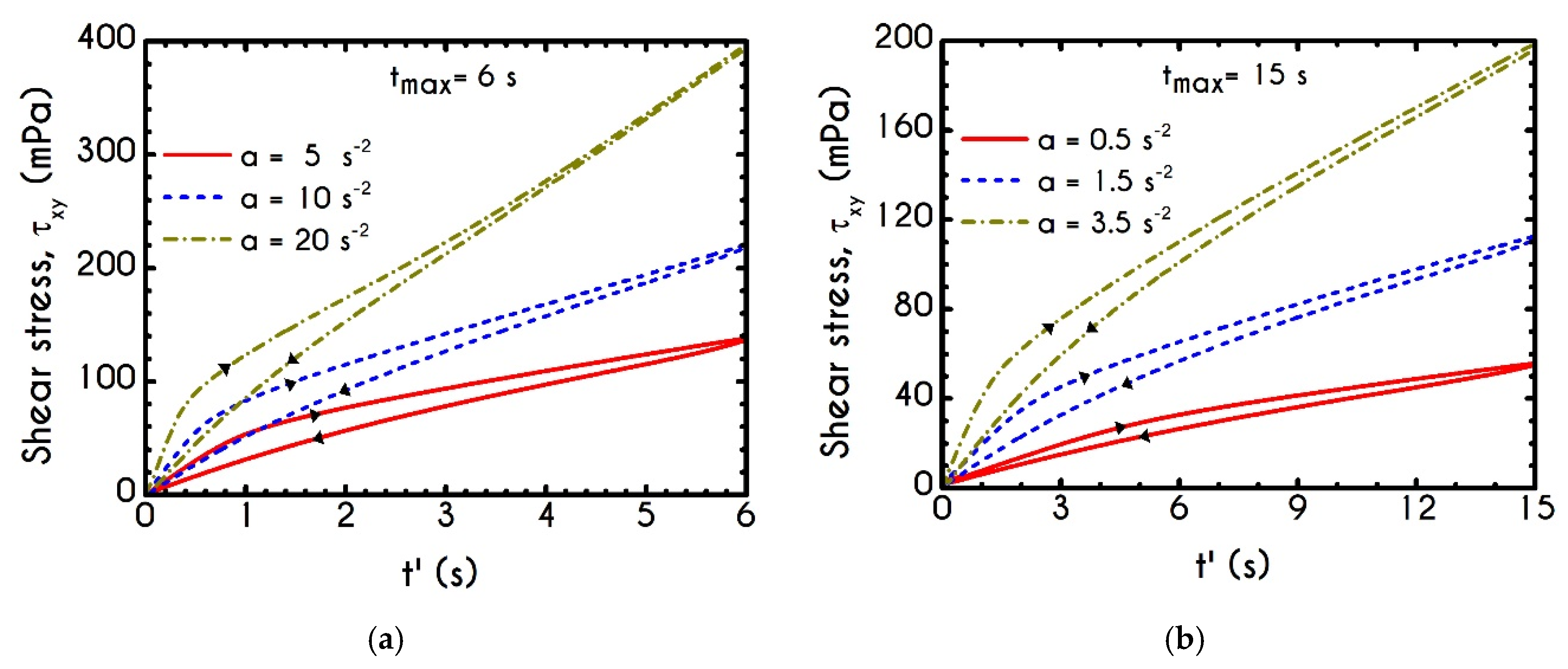
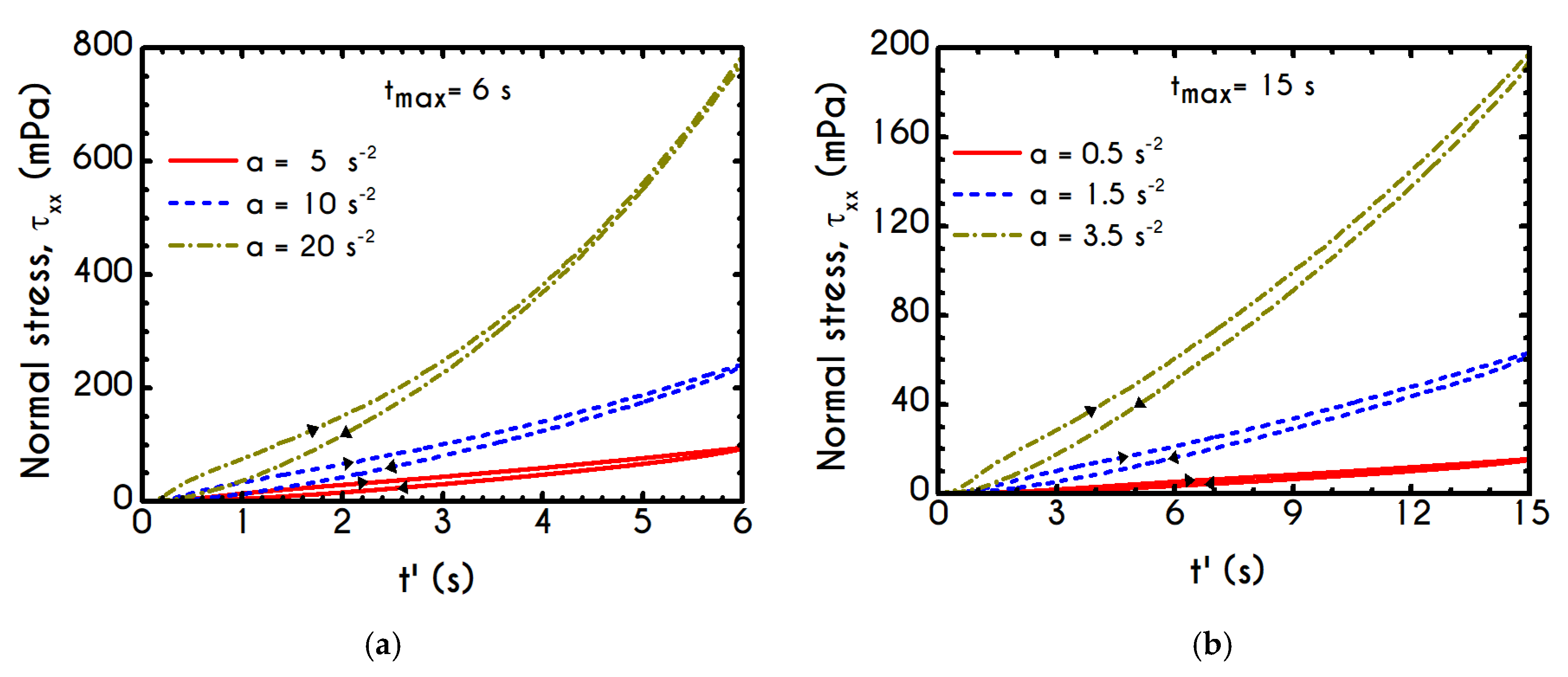
Having determined the model parameters from available experiments, we proceed with predictions for other variables under the same two experimental procedures. In
Figure 2a,b, we present the predictions of our model for the normal stress component as well as the ratio
using the rheological parameters of
Table 1 exported by the previous fitting procedure. Regarding
Figure 2, both normal stress
and the ratio
depict a similar dependence on shear rate. Except for the shear stress, viscoelastic materials or biological systems develop normal stresses even in pure shear flows, which sometimes are found to be equal or to exceed
[
75]. At low values of the imposed shear-rate, our model predicts an almost zero normal stress, while at moderate to high shear rates, the flow curve demonstrates an abrupt increase, especially when
exceeds the value of
. From
Figure 2b we can observe that normal stress has a significant contribution, and it is comparable to the shear component. For shear rates higher than
the ratio
is greater than unity and hence
surpasses the value of
. Apparently, a condition where
exceeds the
is not expected for the blood unless the imposed shear-rates are high enough to produce significant velocity gradients in the xx- direction. The xx- stress component is originated at the extension of the plasmatic proteins that bridge the rouleaux components in the x-direction as blood is sheared in the same direction, as well as the extension of the membrane of each RBC [
76]. Most of the blood constitutive modelling investigations do not present the normal stress prediction, and hence we are not able to make a comparison with other findings. Varchanis et al. [
62] in their work reported a significant contribution of normal stress in simple shear tests and compared their findings with those predicted by the ML-IKH model, which was found to have similar behavior. The presence of normal stresses is suspensions is attributed the intense interaction between the particles. Moreover, the viscoelastic contribution of the plasma due to protein stretching. Similar arguments are presented by Mall-Gleissle et al. [
77] for suspensions with viscoelastic matrix fluids.
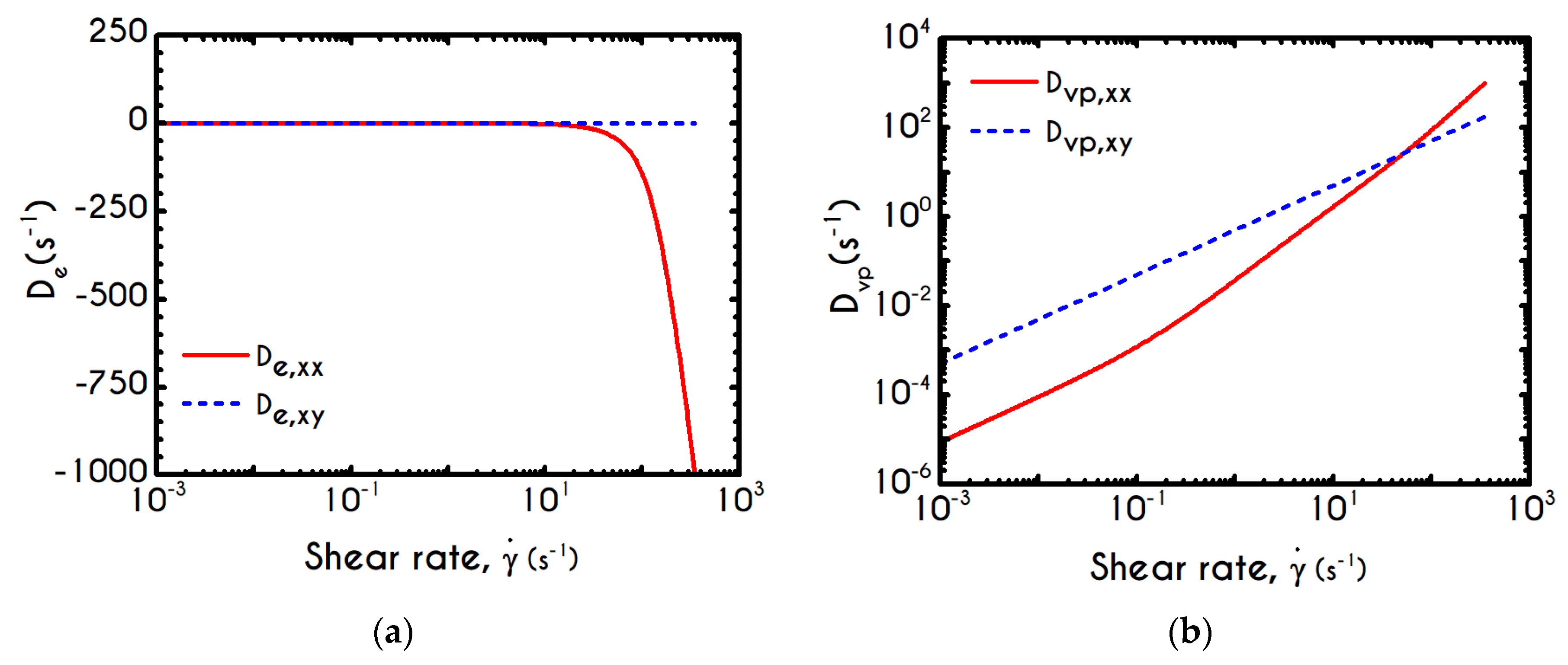
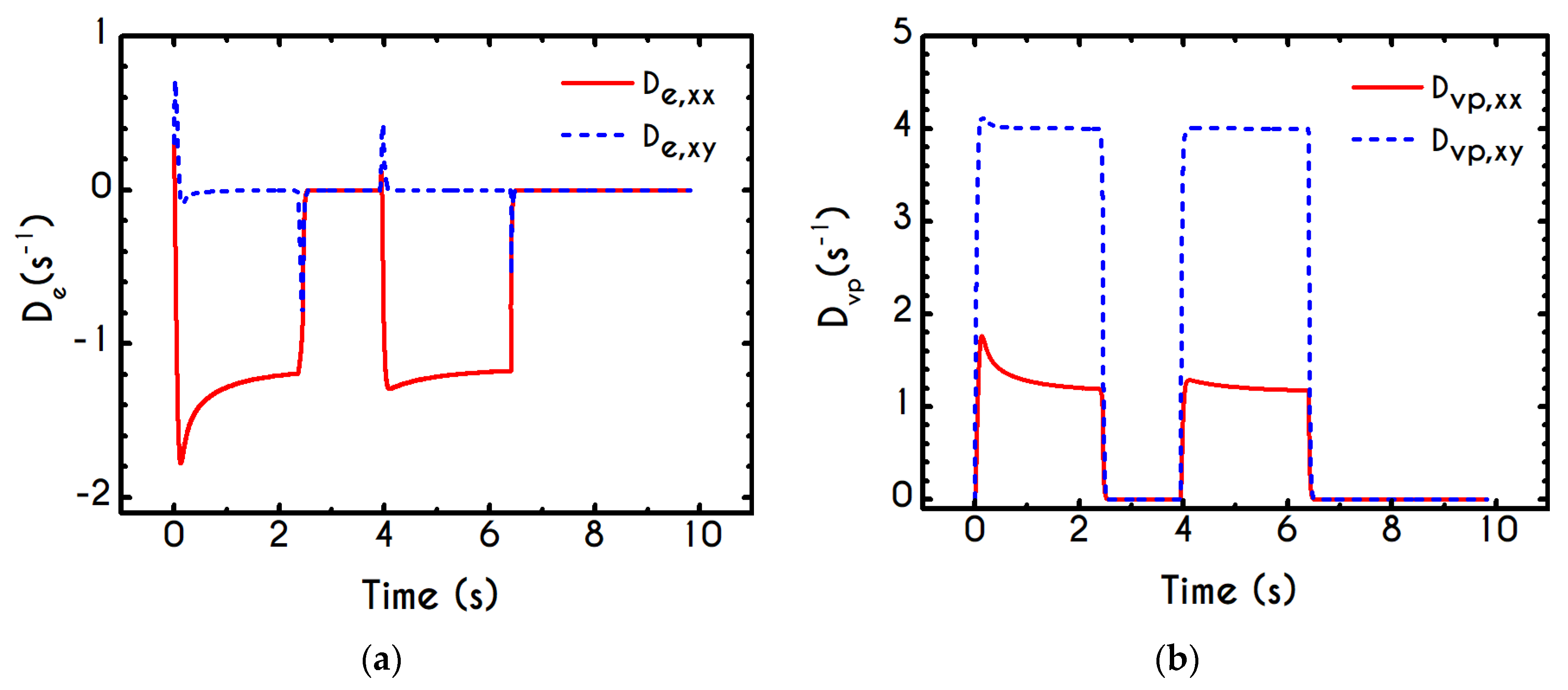
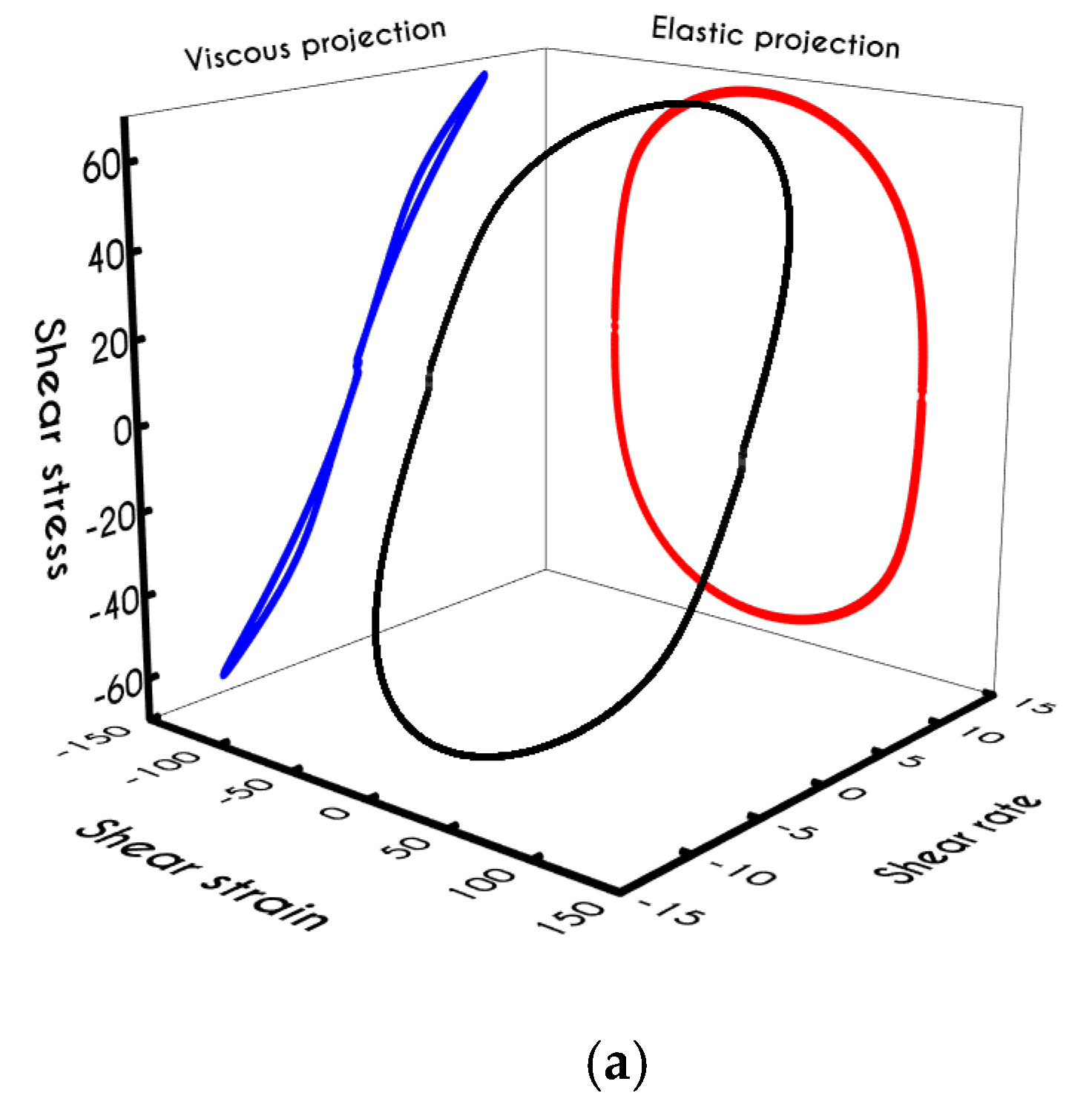
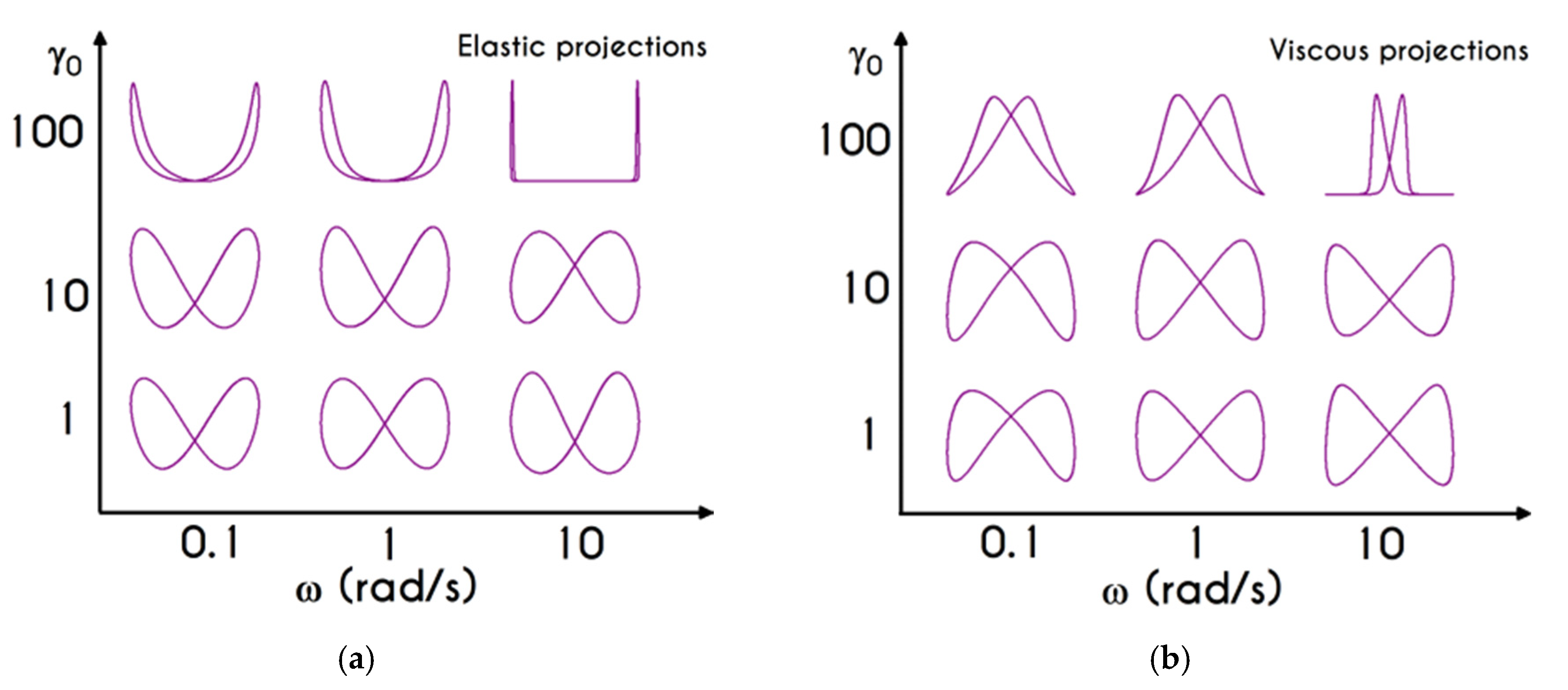
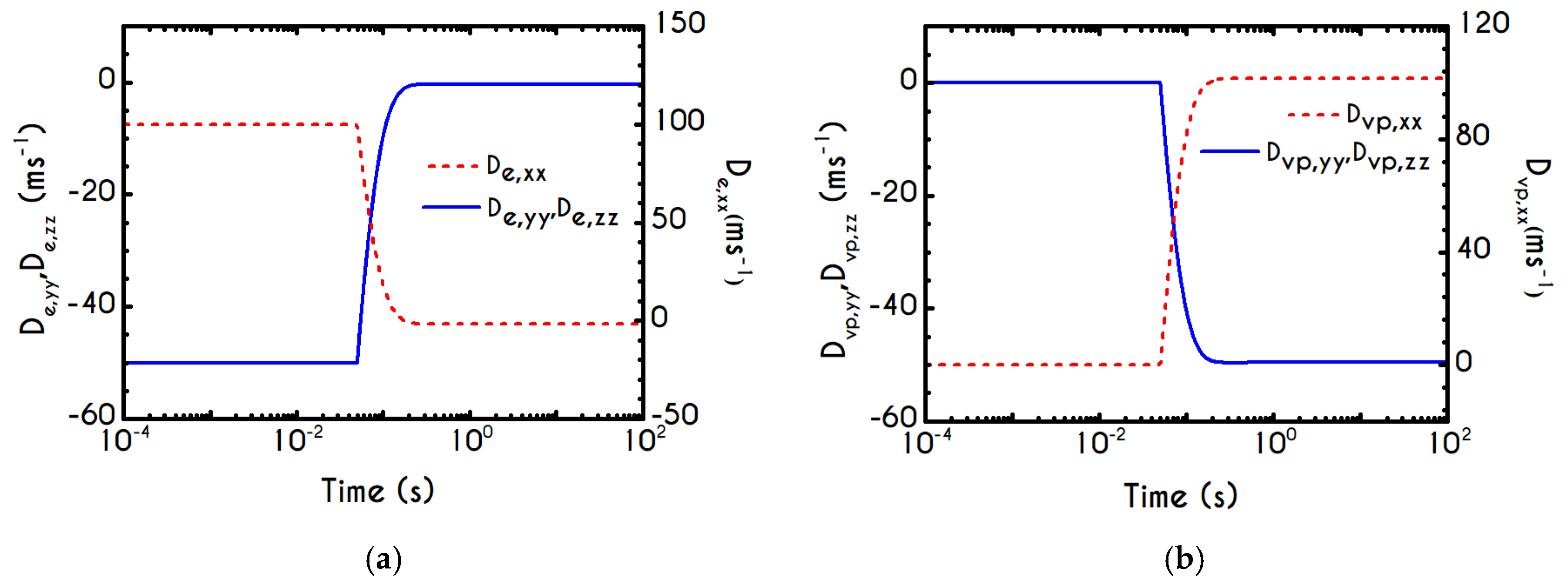
To further analyze the predicted response, and thus improve the understanding of the underlying mechanisms in terms of macroscopic modeling, we present the separate contributions in the deformation-rate tensor (Equation (3)) using the set of parameters of
Table 1 for the experimental data reported in [
47]. In particular, the use of the current thixotropic elasto-viscoplastic model, allows us to probe further the shear-rate development in terms of its elastic and viscoplastic contributions. Particularly,
Figure 3 illustrates the xx- and xy- components of elastic (
Figure 3a) and viscous (
Figure 3b) projections as a function of the imposed shear-rate, referring to the steady-state values of the startup test. As we can see in
Figure 3, in a steady shear experiment, the developed rate of strain is contributed both by elastic and viscoplastic mechanisms. However, the elastic contribution originates from the xx- component, while the gradients in xy- direction are exactly zero across the whole range of the imposed conditions; this happens only upon steady state (
is non-zero during the development of flow), is attributed to the fact that the model predicts
, and is a standard feature of most constitutive models that induce elastic effects via the upper convected derivative). Above the low shear-rate plateau (
), our model predicts an abrupt increase in the absolute value of
, which indicates that blood rouleaux undergoes high strains in the xx- direction. All these agree with our conclusions for the dependence of the stress components on the shear-rate.
Figure 3a demonstrates that viscoplasticity is equally significant in both directions for shear-rates up to ~
. When
is higher than
the velocity gradients in the xx- direction are high enough to make the contribution of the xx- component of viscoplastic deformation tensor is more significant than the xy- one. Indicatively, for an imposed shear rate
equal to
the
is equal to
while the
is only
.
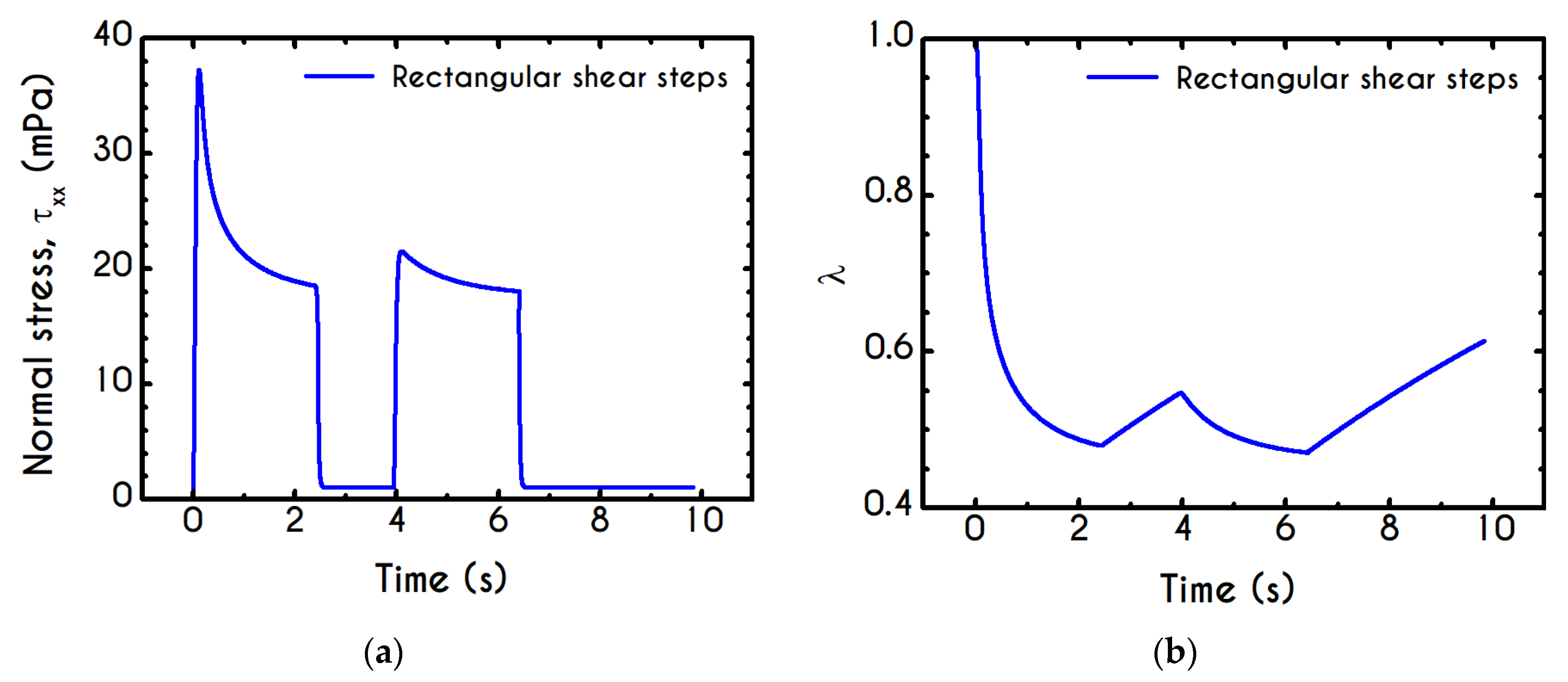
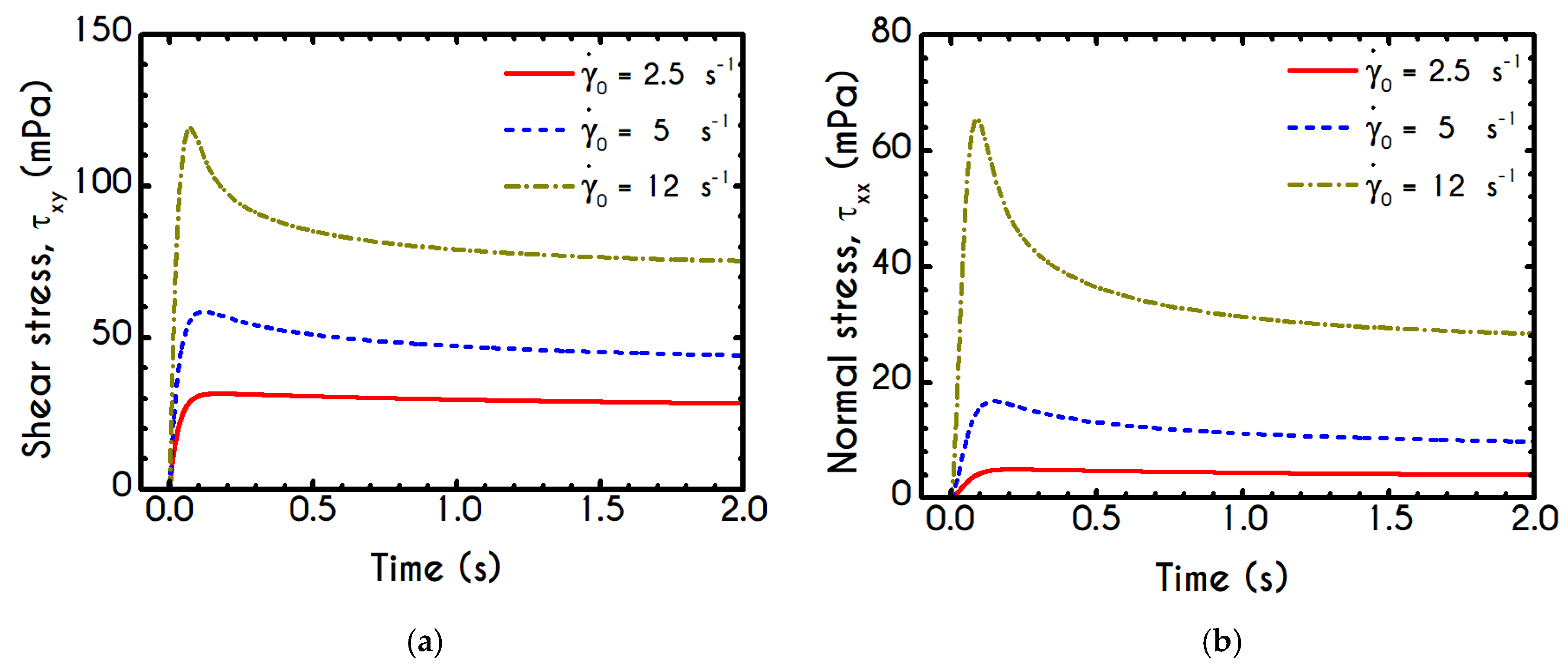
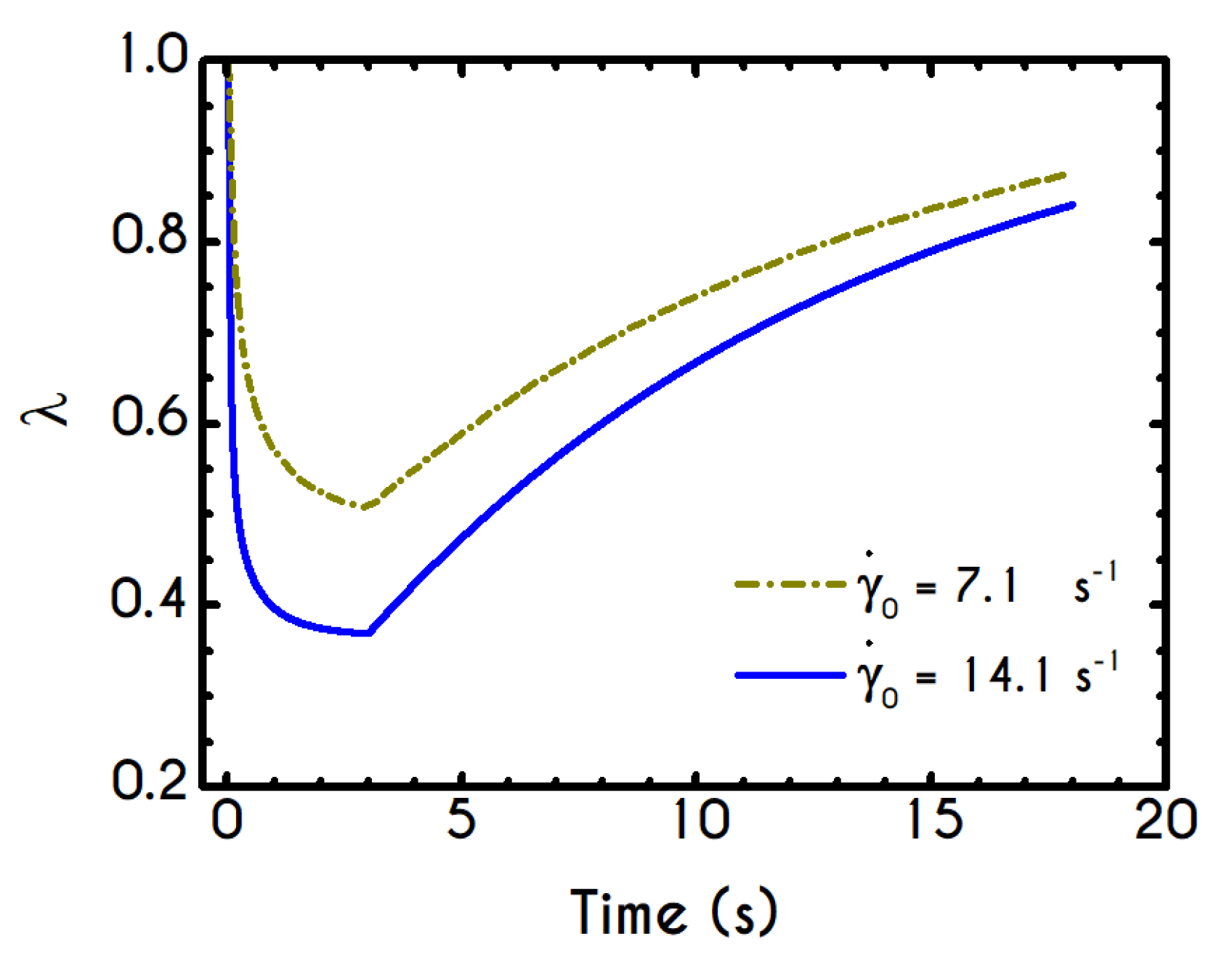
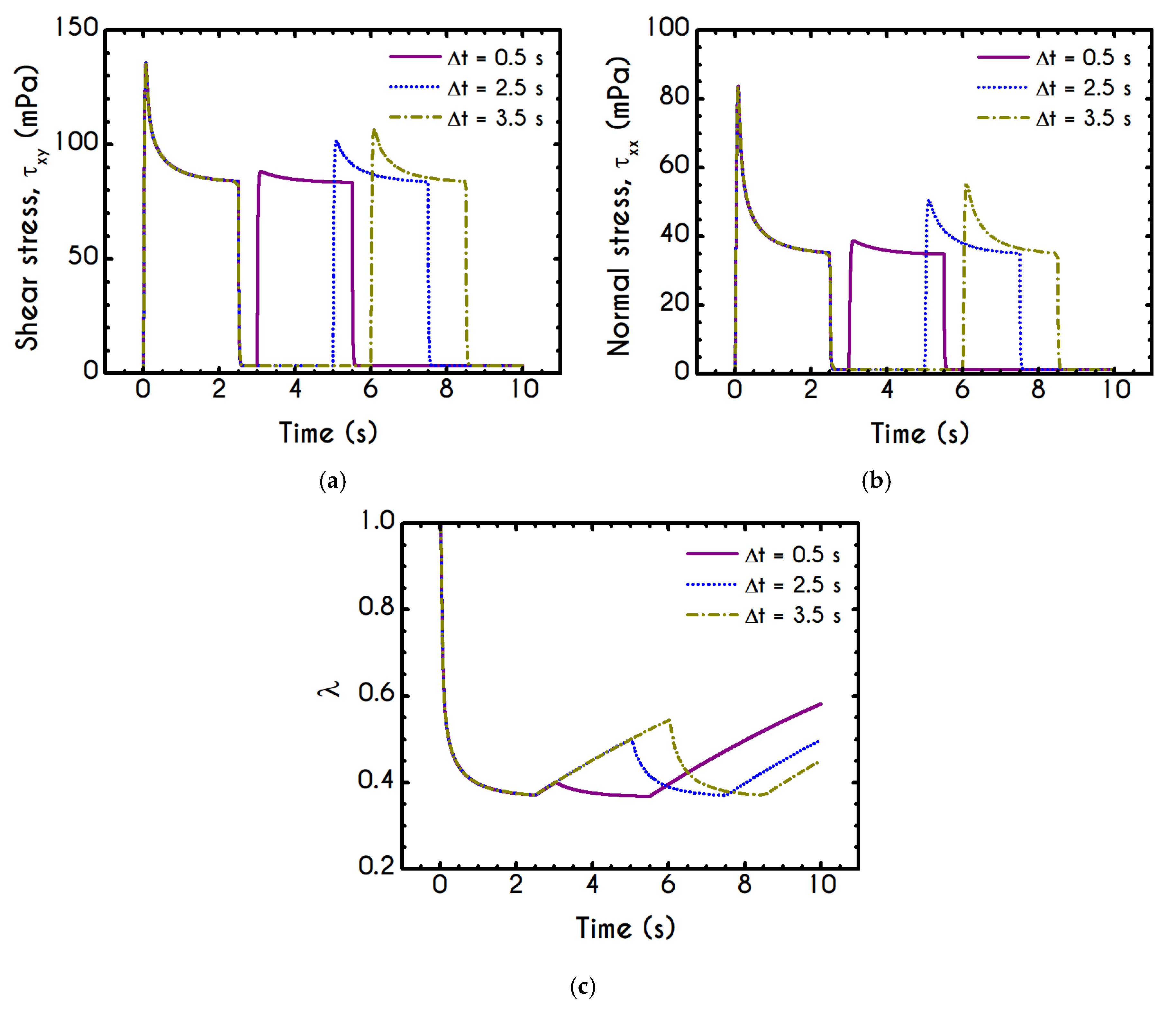
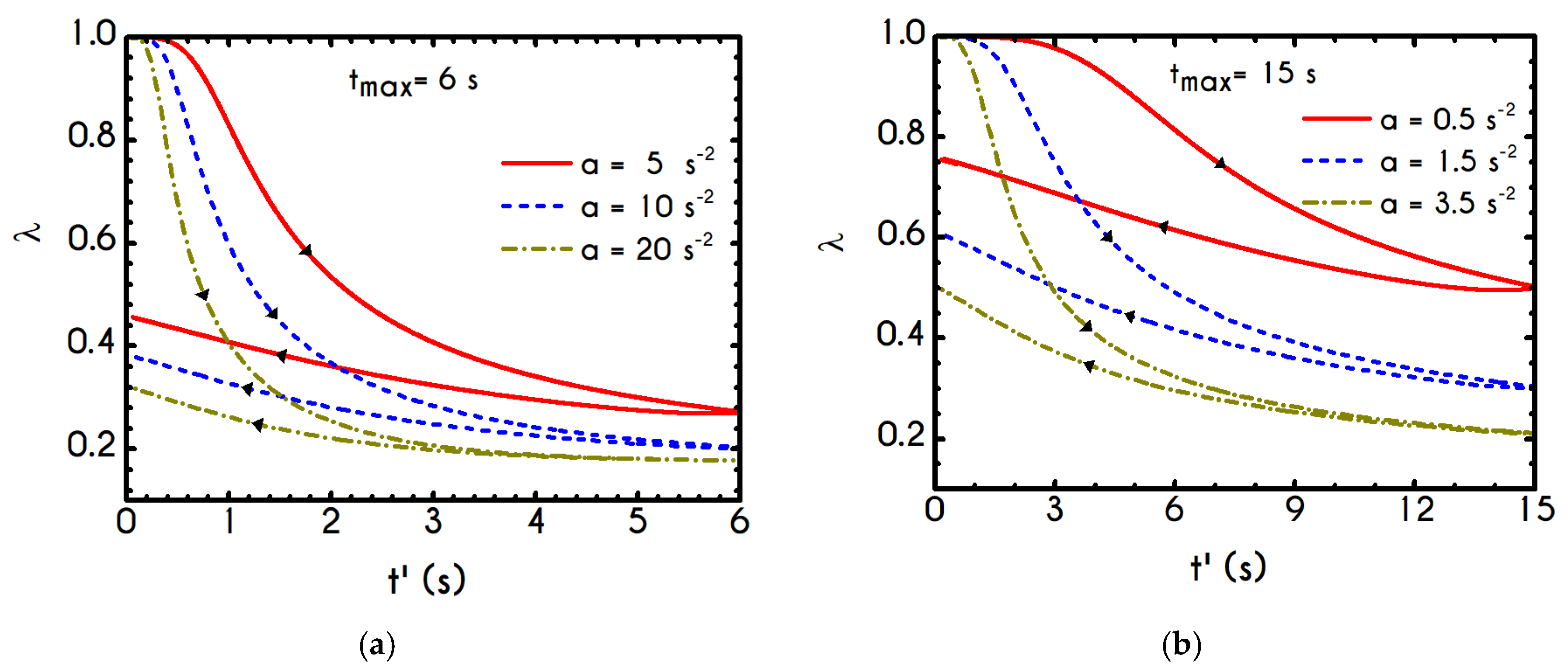
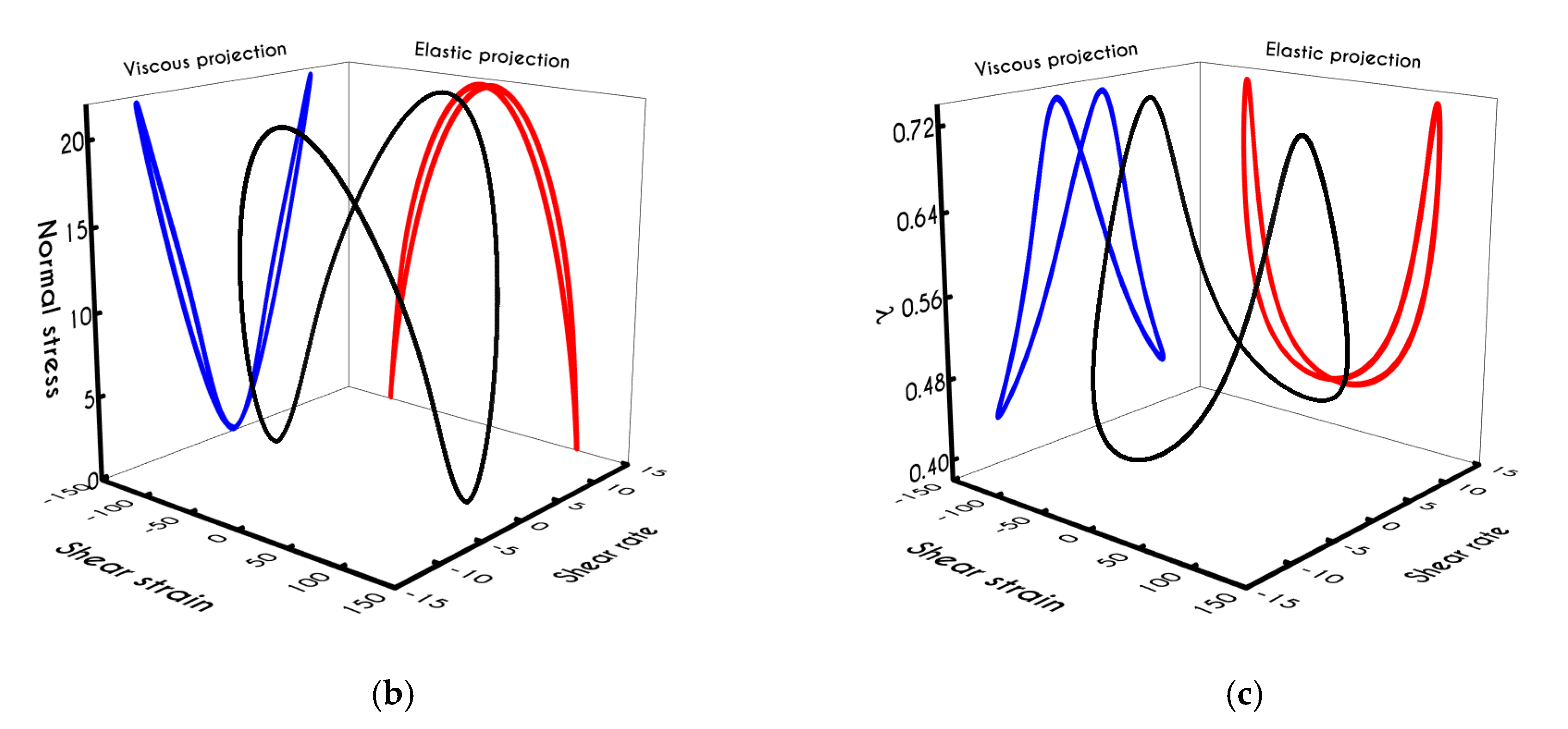
Using the set of rheological parameters of
Table 1 for the experimental data reported in [
47], we can predict the normal stress (
Figure 4a) and the structure parameter (
Figure 4b) for the intermittent shear-step experiment [
47]. Quantitively, the evolution of
resembles that of the shear-stress (
Figure 1b). However, similar to the steady experiments, the xx- component takes smaller values than the xy- component (almost half) due to the relatively small value of shear-rate pulses (
). Carefully studying
Figure 4a, we observe an initial overshoot that is a direct consequence of the combination of elastic effects and the thixotropic evolution of the viscosity [
1], which is a typical behavior for VE materials or biological systems. We are going to discuss this in detail in the following sections. This can be justified by the implication of the shearing on the stretch of the RBC membrane and the rest of the components of blood, which depicts that the elastic response of blood is significant, especially in the xx- direction, as it is illustrated in
Figure 5a. The contribution of the xy- component is negligible, enhancing the fact that stretching of blood is mainly in the flow direction, which may explain the tendency of the RBCs to align with the flow [
78]. After the interruption of the shearing, the stresses are drastically reduced until the imposition of the new shearing, producing a second but much milder overshoot. Regarding the structure parameter, it follows a decrease with increasing the shear-rate, indicating a continuous deconstruction of the rouleaux (
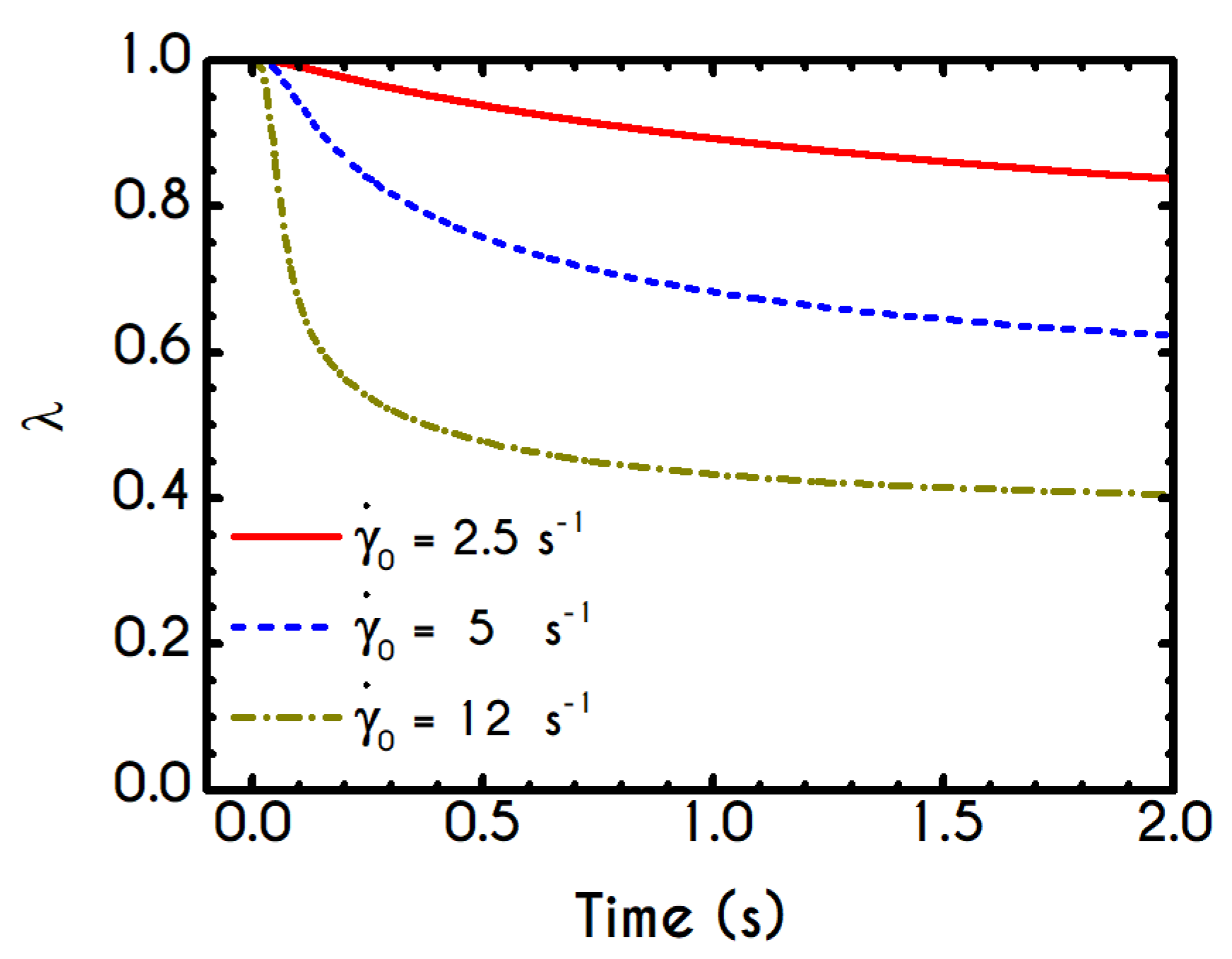
Figure 4b). This behavior is also observed during the second shear imposition, but the deconstruction of blood follows a much smaller reduction. During the period of flow cessation, the stresses present in the system are small enough to promote the rebuilding of the aggregates, providing a linear increase of
which lasts as long as the intermittent steps. Particularly, during the intermittent steps test, the first overshoot is observed at
while the second at
while the corresponding values for the shear stress are equal to
and
respectively. The impact of this test on the structure parameter is high enough to induce a
at the end of the first shearing while blood manages to recover its microstructure by 15%. After imposing the second shearing, the microstructure is disintegrated again to a value equal to
while after the cessation of the shearing this value is increased to
which corresponds to the last observable value in our test. Longer duration of cessation would lead
λ to increase further.
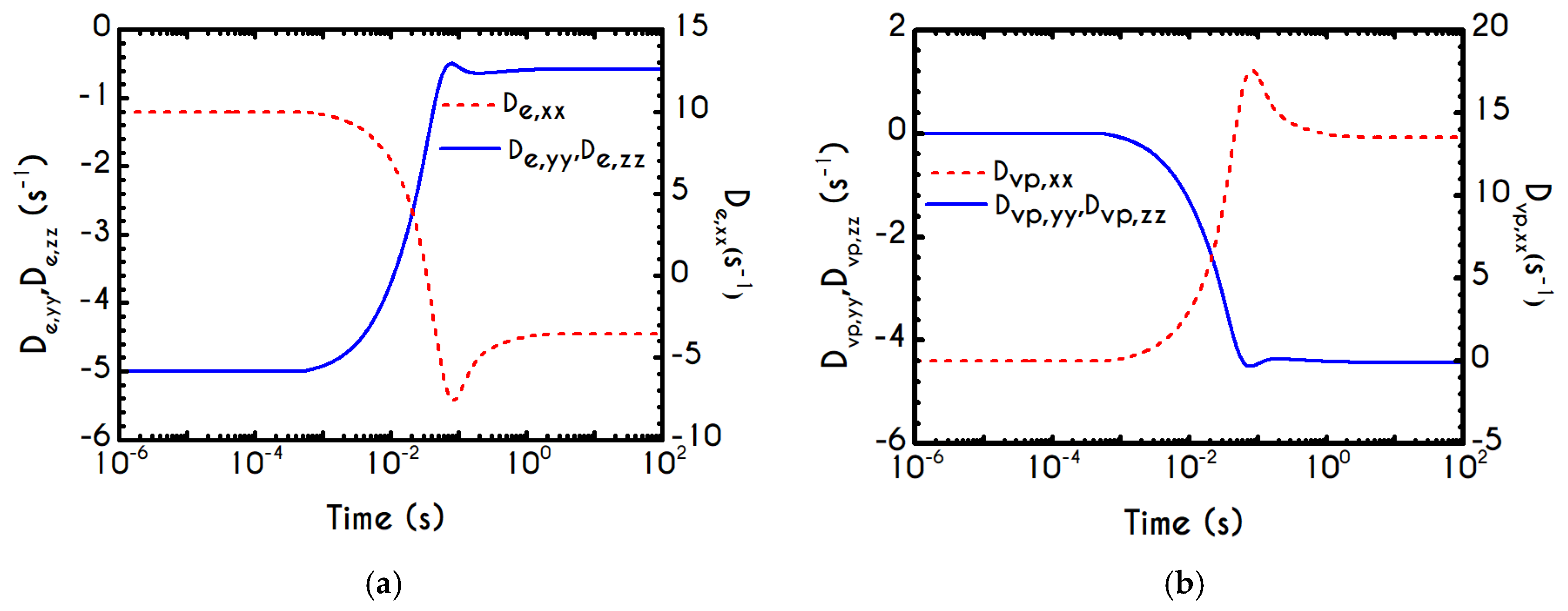
If we examine the normal elastic component of the deformation-rate (
Figure 5a), we will realize that it follows a similar evolution with that predicted for the normal stress (
Figure 4a), exhibiting an overshoot at the beginning of each shearing pulse. On the other hand, the shear elastic contribution is zero during the shearing, which means that viscoplasticity dominates the shear forces. Regarding the viscoplastic part of the deformation-rate tensor (
Figure 5b), both its components exhibit smoother variations in comparison with the elastic components. In particular, the shear viscoplastic deformation-rate increases smoothly up to
at the end of the first pulse, and up to
at the end of the second pulse, while the normal component takes an order of magnitude smaller values.
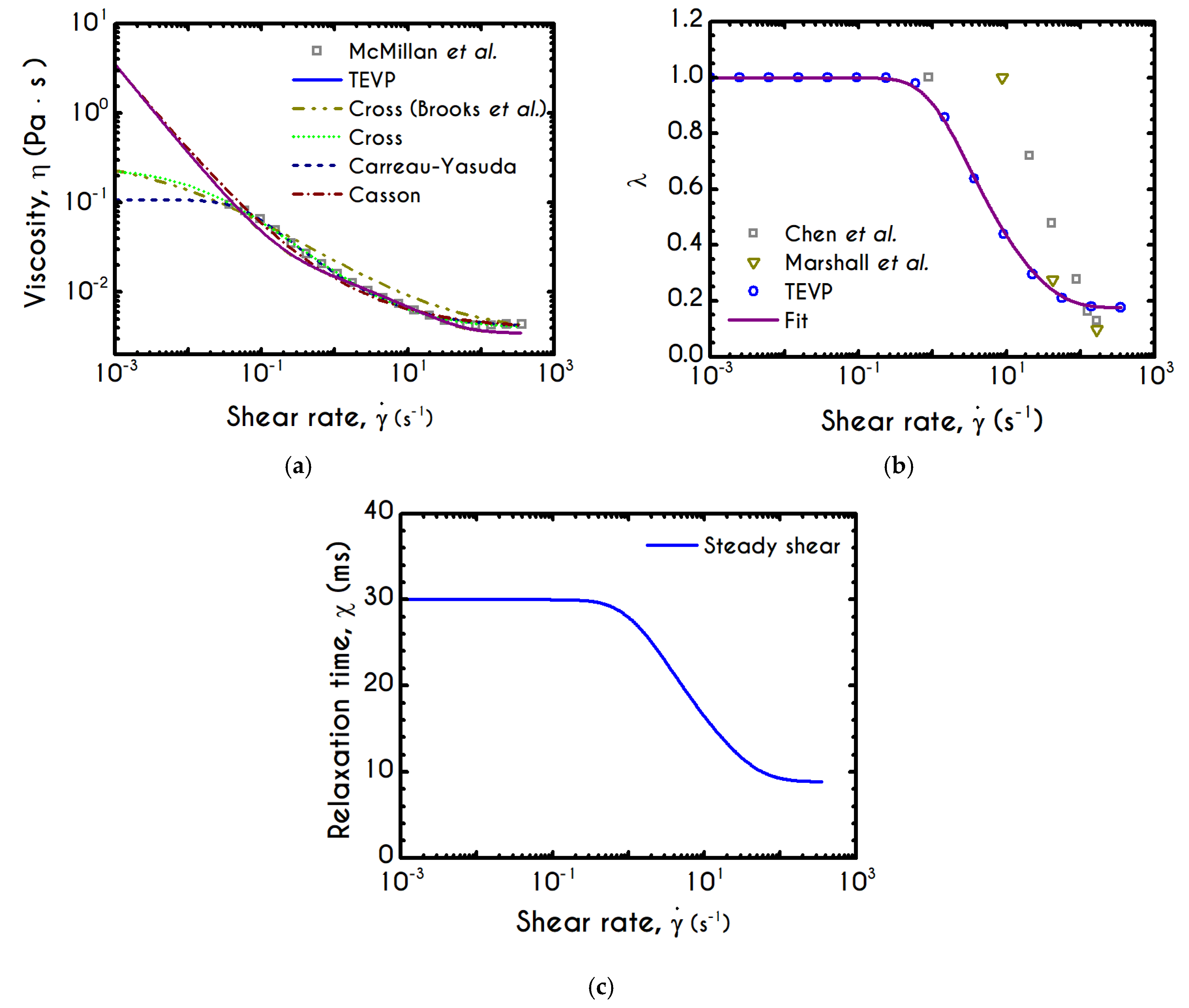
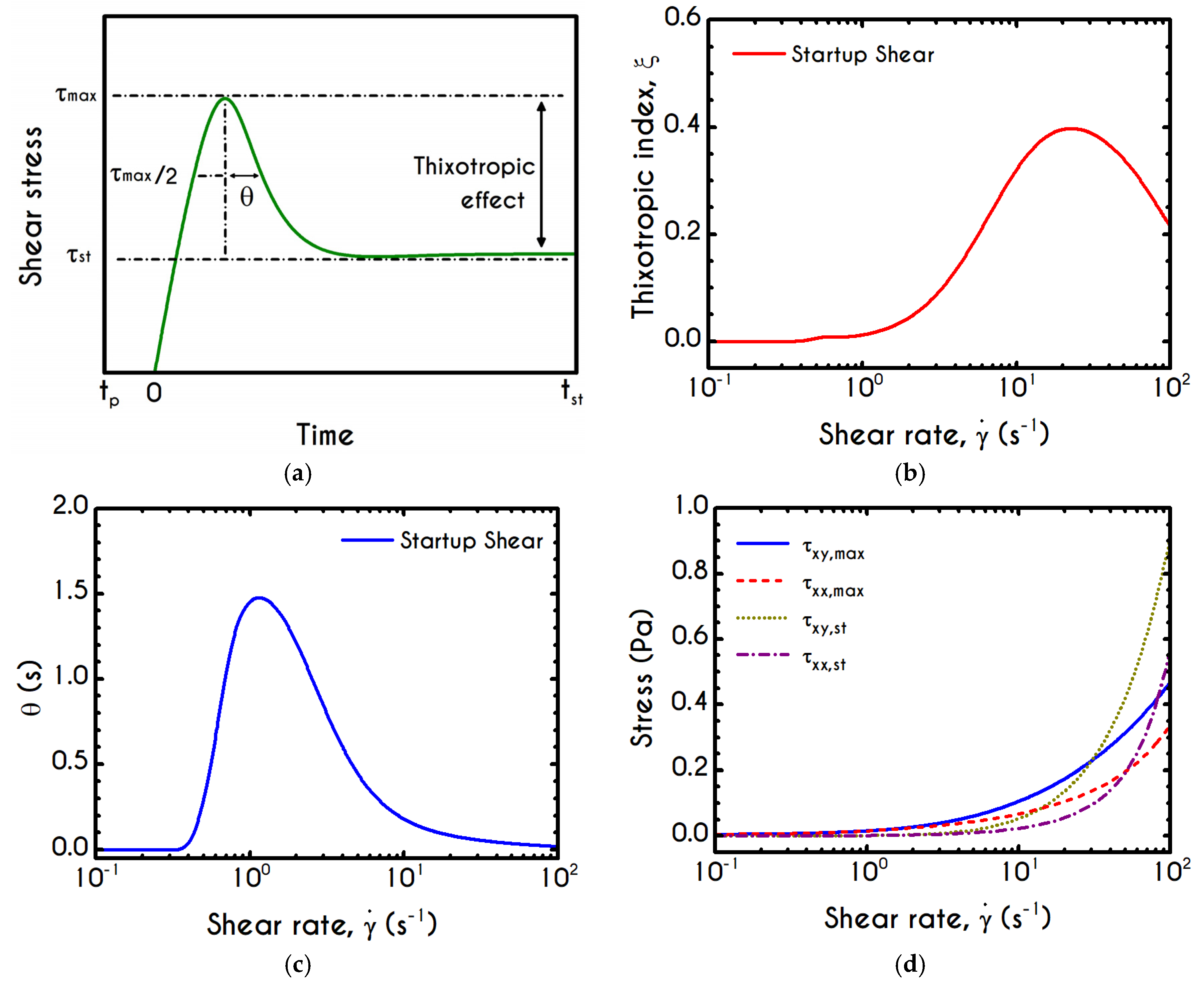
Figure 6a presents the experimental measurements of the apparent viscosity with respect to the imposed shear-rate for a hematocrit equal to 45% from the data reported in [
79]. Along with our model predictions for the aforementioned hematocrit, we also reproduce those of the Casson, the Cross, and the Carreau-Yasuda models for improved clarity. The parameters of these models have been determined in
Appendix A.
We observe that our model follows closely the predictions of the Casson model, and also captures the experimental data with an excellent agreement for shear-rates above
, but slightly deviates by 18% from the experimental viscosity below this value. The deviation is strongly associated with the aggregability of blood or in terms of modeling, with the kinetic constants of Equation (8). Regarding the inelastic models, they present an excellent agreement with experiments across their range of validity. However, they provide different predictions for the zero-shear-rate viscosity, which sometimes are unrealistic (e.g., finite zero-shear-rate viscosity). The generalized Newtonian models are algebraic, phenomenological, and able to capture only the steady-state viscous or viscoplastic behavior of blood. In contrast, they are inadequate to reproduce transient as well as elastic effects. Regarding our model’s prediction, the curve fitting in
Figure 1 is excellent, even at high shear rates. When the fitting curve of stress is translated into an apparent viscosity curve, at high shear rates, the deviation from the experimental data is more pronounced. When the fitting curve of stress is translated into an apparent viscosity curve, at high shear rates, the deviation from the experimental data is more pronounced. The high-shear experimental viscosity is about
, while the theoretically predicted value is about
, which is about 3.5–4 times greater than that of plasma (
). If we bear in mind that: (a) our model is fitted in steady and transient experimental data simultaneously, (b) there are significant experimental errors because at high shear-rates the flow in a cone-and-plate rheometer is inhomogeneous. Our model does not only account for steady-state phenomena but has also been configured to reproduce transient experiments, revealing the complex rheological behavior of blood such as elasto-visco-plasticity along with potential thixotropic effects.
As noted in the introduction, RBC aggregability is an important determinant of hemodynamics [
82], particularly due to its association with microcirculatory disorders, mainly cardiovascular diseases [
83,
84], because the growth of the size of agglomerates may lead to reduced tissue perfusion for example. Although our model does not explicitly account for the RBCs aggregation size, it describes in detail the level of structure of the RBCs, which depends dynamically on the imposed rheological conditions. In
Figure 6b, we can see the structure parameter predicted by our model as a function of the shear-rate. For low values of
, plasticity dominates the flow, the applied stresses are slightly above the yield stress and hence deconstruction term is close to zero. Consequently, blood continues to be in a fully structured state until the shear rate reaches a critical value above which the clustering is starting to collapse. The abrupt reduction of
is then followed by a stabilized condition where the internal microstructure does not exhibit further deconstruction. Irrespective of the intensity of shear rate,
never reaches
, but asymptotes to a finite value indicating that the material never becomes fully unstructured. In order to make a sufficient comparison of our model predictions with those exported by experimental observations of the rouleau size, we invoke the work of Chen et al. [
80]. They provided quantitative measures of RBC aggregability from direct visual monitoring of the aggregation and disaggregation processes. If we consider that at the lowest shear rate, the level of structure is the highest and equal to unity, we are able to correlate any other state of the blood as a percentage of the fully structured state. Before comparison, we transform the median aggregate size of RBCs given by Chen et al. by dividing these values with the average size at the zero-shear rate. After that,
Figure 6b illustrates the predictions of TEVP along with the transformed data of RBC median aggregate size at equilibrium as a function of shear rate. Qualitatively the model predictions are in quite good agreement with the experimental data if one considers that our simulation accounts for a hematocrit value equal to 45%. In comparison, the experiment has been conducted for a significantly lower value of about 10%. As a result, in our case, the interaction of the dissolved phase is more intense and “unjamming” and “disentanglment” phenomena appear at lower values of the applied shear rate, something that is consistent with the data that we present. Despite the large deviation on the hematocrit level, our model captures the continuous deconstruction of blood’s microstructure as shear rate increases but with a steeper way than the experimental observation. Although the quantitative deviation is great enough below
, for higher values of the shear rate, we observe an overall discrepancy of about 12%. It should be mentioned that the average aggregation size can be optically measured in quite transparent systems when the suspension or the blood is relatively dilute (e.g., systemic hematocrit below 15%). For the sake of convenience, we also illustrate the fit of a sigmoid like function (
) on our theoretical predictions of the TEVP model in order to provide a more natural way to reproduce the predicted microstructural response. The proposed function of the thixotropic structure parameter
distribution with respect to shear rate is given by the following expression, while the corresponding parameters are illustrated in
Table 2:
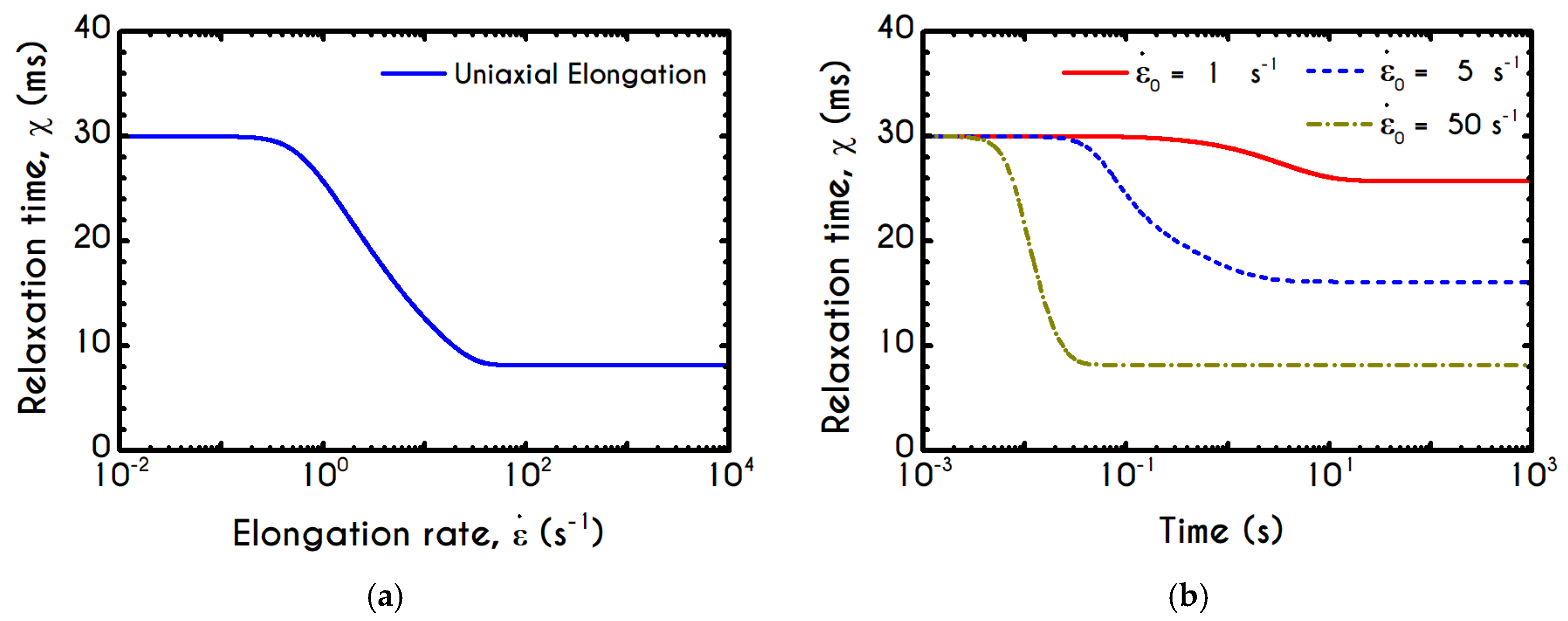
The relaxation time of the blood sample,
, is given by Equations (13) and (15). It exhibits a similar sigmoidal variation as a function of the shear-rate
(
Figure 6c). It smoothly decays from
to
, which is the asymptotic value in the limit of high shear-rates. The variation occurs in the range of
to
, where rouleaux rearrangements and disintegration take place [
85]. Although we have a qualitative agreement with the experiments by Thurston & Henderson [
85] for the range of variation of the relaxation time, there is a deviation between the model predictions and their estimations. This is strongly related to the experimental procedure and the theoretical approximation that Thurston & Henderson [
85] have adopted. Specifically, they provide estimates of the relaxation-time based on the definitions of the theory of linear viscoelasticity (Maxwell model, which is valid only for infinitesimal deformations), while the experiments where performed in the nonlinear regime (Bird et al. [
86]). They made this poor assumption in the whole range of shear-rate. Thus, from this work, we can only be sure for the shear-thinning dependence of the relaxation time, which match pretty good with the prediction of TEVP model. However, there is no indication that experiments were done in the limit of infinitesimal strains. On the contrary, the imposition of any finite rate of strain (or velocity field) causes the yielding of the blood sample and its flow. Thus, the experiments were executed in the nonlinear regime, where the Maxwell theory is not valid. However, the systematic misassumption that they introduced in their calculation, does not affect the dependence of the relaxation time on the shear rate, but only affects the magnitude of the blood relaxation time.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






























