
Potential for and Distribution of Enzymatic Biodegradation of Polystyrene by Environmental Microorganisms



Abstract
:1. Introduction
2. Materials and Methods
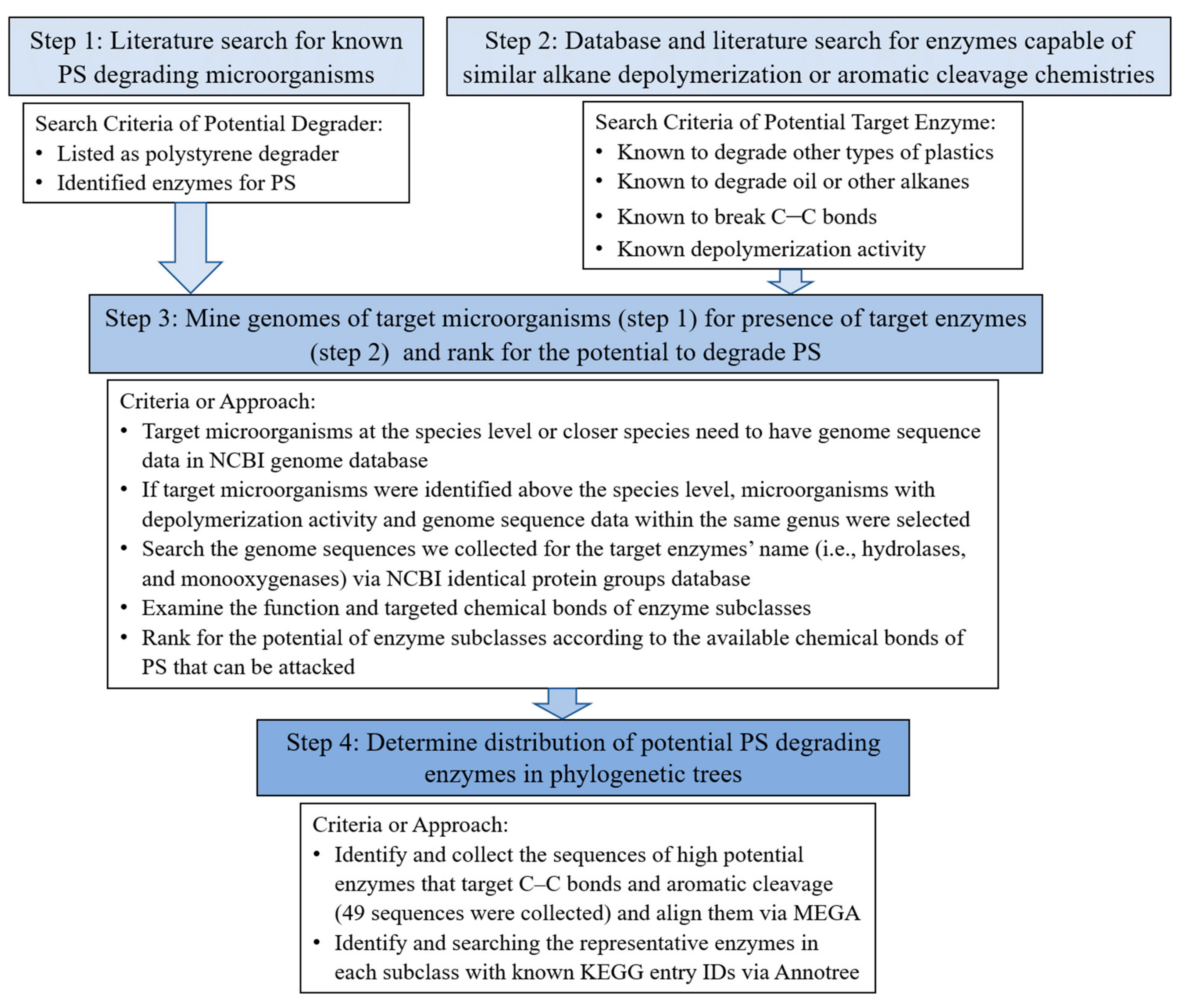
2.1. Literature Search for Microbes and Enzymes with Potential PS Degrading Ability
2.2. Mining Genomes of Target Microorganisms for the Presence of Target Enzymes
2.3. Visualization of Enzyme Sequence Relationships and Distributions in Genomes and Phylogentic Trees
3. Results and Discussion
3.1. Potential PS Degradting Enzymes
3.2. Distribution of Potential PS Degradting Enzymes across Domian Bacteria
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Plastics Europe-Association of Plastics Manufacturers. Plastics—The Facts 2017. Available online: https://www.issuelab.org/resource/plastics-the-facts-2017-an-analysis-of-european-plastics-production-demand-and-waste-data.html (accessed on 1 January 2018).
- Yuan, J.; Ma, J.; Sun, Y.; Zhou, T.; Zhao, Y.; Yu, F. Microbial degradation and other environmental aspects of microplastics/plastics. Sci. Total Environ. 2020, 715, 136968. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Kumar, D.; Yoo, C.G.; Gitsov, I.; Majumder, E.L.W. Conversion and removal strategies for microplastics in wastewater treatment plants and landfills. Chem. Eng. J. 2021, 406, 126715. [Google Scholar] [CrossRef]
- Pathak, V.M. Review on the current status of polymer degradation: A microbial approach. Bioresour. Bioprocess. 2017, 4, 15. [Google Scholar] [CrossRef]
- Syranidou, E.; Karkanorachaki, K.; Amorotti, F.; Franchini, M.; Repouskou, E.; Kaliva, M.; Vamvakaki, M.; Kolvenbach, B.; Fava, F.; Corvini, P.F.; et al. Biodegradation of weathered polystyrene films in seawater microcosms. Sci. Rep. 2017, 7, 17991. [Google Scholar] [CrossRef] [Green Version]
- Muenmee, S.; Chiemchaisri, W.; Chiemchaisri, C. Microbial consortium involving biological methane oxidation in relation to the biodegradation of waste plastics in a solid waste disposal open dump site. Int. Biodeterior. Biodegrad. 2015, 102, 172–181. [Google Scholar] [CrossRef]
- Pattanasuttichonlakul, W.; Sombatsompop, N.; Prapagdee, B. Accelerating biodegradation of PLA using microbial consortium from dairy wastewater sludge combined with PLA-degrading bacterium. Int. Biodeterior. Biodegrad. 2018, 132, 74–83. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, W.; Xing, R.; Xie, S.; Yang, X.; Cui, P.; Lü, J.; Liao, H.; Yu, Z.; Wang, S.; et al. Enhanced in situ biodegradation of microplastics in sewage sludge using hyperthermophilic composting technology. J. Hazard. Mater. 2020, 384, 121271. [Google Scholar] [CrossRef]
- Lou, Y.; Ekaterina, P.; Yang, S.-S.; Lu, B.; Liu, B.; Ren, N.; Corvini, P.F.X.; Xing, D. Biodegradation of polyethylene and polystyrene by greater wax moth larvae (Galleria mellonella L.) and the effect of co-diet supplementation on the core gut microbiome. Environ. Sci. Technol. 2020, 54, 2821–2831. [Google Scholar] [CrossRef]
- Auta, H.S.; Emenike, C.U.; Fauziah, S.H. Screening of Bacillus strains isolated from mangrove ecosystems in Peninsular Malaysia for microplastic degradation. Environ. Pollut 2017, 231, 1552–1559. [Google Scholar] [CrossRef]
- Yan, F.; Wei, R.; Cui, Q.; Bornscheuer, U.T.; Liu, Y.-J. Thermophilic whole-cell degradation of polyethylene terephthalate using engineered Clostridium thermocellum. Microb. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Austin, H.P.; Allen, M.D.; Donohoe, B.S.; Rorrer, N.A.; Kearns, F.L.; Silveira, R.L.; Pollard, B.C.; Dominick, G.; Duman, R.; El Omari, K.; et al. Characterization and engineering of a plastic-degrading aromatic polyesterase. Proc. Natl. Acad. Sci. USA 2018, 115, E4350–E4357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, H.F.; Cho, I.J.; Joo, S.; Seo, H.; Sagong, H.-Y.; Choi, S.Y.; Lee, S.Y.; Kim, K.-J. Rational protein engineering of thermo-stable PETase from Ideonella sakaiensis for highly efficient PET degradation. ACS Catal. 2019, 9, 3519–3526. [Google Scholar] [CrossRef]
- Vroman, I.; Tighzert, L. Biodegradable Polymers. Materials 2009, 2, 307–344. [Google Scholar] [CrossRef]
- Dharmalingam, S.; Hayes, D.G.; Wadsworth, L.C.; Dunlap, R.N.; DeBruyn, J.M.; Lee, J.; Wszelaki, A.L. Soil degradation of polylactic acid/polyhydroxyalkanoate-based nonwoven mulches. J. Polym. Environ. 2015, 23, 302–315. [Google Scholar] [CrossRef]
- Danso, D.; Chow, J.; Streit, W.R. Plastics: Environmental and biotechnological perspectives on microbial degradation. Appl Environ. Microb 2019, 85, e01095-19. [Google Scholar] [CrossRef] [Green Version]
- Palm, G.J.; Reisky, L.; Böttcher, D.; Müller, H.; Michels, E.A.P.; Walczak, M.C.; Berndt, L.; Weiss, M.S.; Bornscheuer, U.T.; Weber, G. Structure of the plastic-degrading Ideonella sakaiensis MHETase bound to a substrate. Nat. Commun. 2019, 10, 1717. [Google Scholar] [CrossRef]
- Ho, B.T.; Roberts, T.K.; Lucas, S. An overview on biodegradation of polystyrene and modified polystyrene: The microbial approach. Crit. Rev. Biotechnol. 2018, 38, 308–320. [Google Scholar] [CrossRef]
- Krueger, M.C.; Seiwert, B.; Prager, A.; Zhang, S.; Abel, B.; Harms, H.; Schlosser, D. Degradation of polystyrene and selected analogues by biological Fenton chemistry approaches: Opportunities and limitations. Chemosphere 2017, 173, 520–528. [Google Scholar] [CrossRef]
- Johnston, B.; Radecka, I.; Hill, D.; Chiellini, E.; Ilieva, V.I.; Sikorska, W.; Musiol, M.; Zieba, M.; Marek, A.A.; Keddie, D.; et al. The Microbial Production of Polyhydroxyalkanoates from Waste Polystyrene Fragments Attained Using Oxidative Degradation. Polymers 2018, 10, 957. [Google Scholar] [CrossRef] [Green Version]
- Mohan, A.J.; Sekhar, V.C.; Bhaskar, T.; Nampoothiri, K.M. Microbial assisted High Impact Polystyrene (HIPS) degradation. Bioresour. Technol. 2016, 213, 204–207. [Google Scholar] [CrossRef]
- Przemieniecki, S.W.; Kosewska, A.; Ciesielski, S.; Kosewska, O. Changes in the gut microbiome and enzymatic profile of Tenebrio molitor larvae biodegrading cellulose, polyethylene and polystyrene waste. Environ. Pollut. 2020, 256, 113265. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Zimmermann, W. Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: How far are we? Microb. Biotechnol. 2017, 10, 1308–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Cui, Z.; Nie, K.; Cao, H.; Jiang, M.; Xu, H.; Tan, T.; Liu, L. A quantum mechanism study of the C-C bond cleavage to predict the bio-catalytic polyethylene degradation. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yang, J.; Wu, W.M.; Zhao, J.; Song, Y.L.; Gao, L.C.; Yang, R.F.; Jiang, L. Biodegradation and mineralization of polystyrene by plastic-eating mealworms: Part 2. Role of gut microorganisms. Environ. Sci. Technol. 2015, 49, 12087–12093. [Google Scholar] [CrossRef]
- O’Leary, N.D.; O’Connor, K.E.; Dobson, A.D.W. Biochemistry, genetics and physiology of microbial styrene degradation. Fems Microbiol. Rev. 2002, 26, 403–417. [Google Scholar] [CrossRef] [Green Version]
- Grbicgalic, D.; Churchmaneisel, N.; Mrakovic, I. Microbial transformation of styrene by anaerobic consortia. J. Appl. Bacteriol. 1990, 69, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.-L.; Kuo, T.-A.; Liu, H.-H. The study of the microbes degraded polystyrene. Adv. Technol. Innov. 2017, 2, 4. [Google Scholar]
- Raddadi, N.; Fava, F. Biodegradation of oil-based plastics in the environment: Existing knowledge and needs of research and innovation. Sci. Total Environ. 2019, 679, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Sekhar, V.C.; Nampoothiri, K.M.; Mohan, A.J.; Nair, N.R.; Bhaskar, T.; Pandey, A. Microbial degradation of high impact polystyrene (HIPS), an e-plastic with decabromodiphenyl oxide and antimony trioxide. J. Hazard. Mater. 2016, 318, 347–354. [Google Scholar] [CrossRef]
- Urbanek, A.K.; Rybak, J.; Wrobel, M.; Leluk, K.; Mironczuk, A.M. A comprehensive assessment of microbiome diversity in Tenebrio molitor fed with polystyrene waste. Environ. Pollut. 2020, 262, 114281. [Google Scholar] [CrossRef]
- Kim, H.R.; Lee, H.M.; Yu, H.C.; Jeon, E.; Lee, S.; Li, J.; Kim, D.-H. Biodegradation of Polystyrene by Pseudomonas sp. Isolated from the Gut of Superworms (Larvae of Zophobas atratus). Environ. Sci. Technol. 2020, 54, 6987–6996. [Google Scholar] [CrossRef] [PubMed]
- Brandon, A.M.; Gao, S.H.; Tian, R.; Ning, D.; Yang, S.S.; Zhou, J.; Wu, W.M.; Criddle, C.S. Biodegradation of polyethylene and plastic mixtures in mealworms (larvae of Tenebrio molitor) and effects on the gut microbiome. Environ. Sci. Technol. 2018, 52, 6526–6533. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.Y.; Su, Y.; Chen, Z.; Chen, J.; Zhou, X.; Benbow, M.E.; Criddle, C.S.; Wu, W.M.; Zhang, Y. Biodegradation of polystyrene by dark (Tenebrio obscurus) and yellow (Tenebrio molitor) mealworms (Coleoptera: Tenebrionidae). Environ. Sci. Technol. 2019, 53, 5256–5265. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, J.; Xia, M. Biodegradation and mineralization of polystyrene by plastic-eating superworms Zophobas atratus. Sci. Total Environ. 2020, 708, 135233. [Google Scholar] [CrossRef]
- Ogonowski, M.; Motiei, A.; Ininbergs, K.; Hell, E.; Gerdes, Z.; Udekwu, K.I.; Bacsik, Z.; Gorokhova, E. Evidence for selective bacterial community structuring on microplastics. Environ. Microbiol. 2018, 20, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Delacuvellerie, A.; Cyriaque, V.; Gobert, S.; Benali, S.; Wattiez, R. The plastisphere in marine ecosystem hosts potential specific microbial degraders including Alcanivorax borkumensis as a key player for the low-density polyethylene degradation. J. Hazard. Mater. 2019, 380, 120899. [Google Scholar] [CrossRef] [PubMed]
- Mor, R.; Sivan, A. Biofilm formation and partial biodegradation of polystyrene by the actinomycete Rhodococcus ruber: Biodegradation of polystyrene. Biodegradation 2008, 19, 851–858. [Google Scholar] [CrossRef]
- Chauhan, D.; Agrawal, G.; Priyadarshini, R.; Deshmukh, S.; Roy, S.S. Biofifilm formation by Exiguobacterium sp. DR11 and DR14 alter polystyrene surface properties and initiate biodegradation. RSC Adv. 2018, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oelschlägel, M.; Zimmerling, J.; Tischler, D. A review: The styrene metabolizing cascade of side-chain oxygenation as biotechnological basis to gain various valuable compounds. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Knott, B.C.; Erickson, E.; Allen, M.D.; Gado, J.E.; Graham, R.; Kearns, F.L.; Pardo, I.; Topuzlu, E.; Anderson, J.J.; Austin, H.P.; et al. Characterization and engineering of a two-enzyme system for plastics depolymerization. Proc. Natl. Acad. Sci. USA 2020, 117, 25476. [Google Scholar] [CrossRef]
- Lenfant, N.; Hotelier, T.; Bourne, Y.; Marchot, P.; Chatonnet, A. Proteins with an alpha/beta hydrolase fold: Relationships between subfamilies in an ever-growing superfamily. Chem. Biol. Interact. 2013, 203, 266–268. [Google Scholar] [CrossRef]
- Shoji, O.; Fujishiro, T.; Nakajima, H.; Kim, M.; Nagano, S.; Shiro, Y.; Watanabe, Y. Hydrogen peroxide dependent monooxygenations by tricking the substrate recognition of cytochrome P450BSβ. Angew. Chem. Int. Ed. 2007, 46, 3656–3659. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, T.; Shoji, O.; Kawakami, N.; Watanabe, T.; Sugimoto, H.; Shiro, Y.; Watanabe, Y. Chiral-substrate-assisted stereoselective epoxidation catalyzed by H2O2-dependent cytochrome P450SPα. Chem. Asian J. 2012, 7, 2286–2293. [Google Scholar] [CrossRef]
- Sabirova, J.S.; Ferrer, M.; Regenhardt, D.; Timmis, K.N.; Golyshin, P.N. Proteomic insights into metabolic adaptations in Alcanivorax borkumensis induced by alkane utilization. J. Bacteriol. 2006, 188, 3763–3773. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, I.; Sumimoto, T.; Ueda, A.; Kusunose, E.; Ichihara, K. Fatty acid-specific, regiospecific, and stereospecific hydroxylation by cytochrome P450 (CYP152B1) from Sphingomonas paucimobilis: Substrate structure required for alpha-hydroxylation. Lipids 2000, 35, 365–371. [Google Scholar] [CrossRef]
- Minerdi, D.; Sadeghi, S.J.; Di Nardo, G.; Rua, F.; Castrignanò, S.; Allegra, P.; Gilardi, G. CYP116B5: A new class VII catalytically self-sufficient cytochrome P450 from Acinetobacter radioresistens that enables growth on alkanes. Mol. Microbiol. 2015, 95, 539–554. [Google Scholar] [CrossRef]
- Moreno, R.; Rojo, F. Enzymes for Aerobic Degradation of Alkanes in Bacteria. In Aerobic Utilization of Hydrocarbons, Oils and Lipids; Rojo, F., Ed.; Springer International Publishing: Cantoblanco, Spain, 2017. [Google Scholar] [CrossRef]
- Van Bogaert, I.N.A.; Groeneboer, S.; Saerens, K.; Soetaert, W. The role of cytochrome P450 monooxygenases in microbial fatty acid metabolism. FEBS J. 2011, 278, 206–221. [Google Scholar] [CrossRef]
- Tsai, Y.-F.; Luo, W.-I.; Chang, J.-L.; Chang, C.-W.; Chuang, H.-C.; Ramu, R.; Wei, G.-T.; Zen, J.-M.; Yu, S.S.F. Electrochemical hydroxylation of C3–C12 n-alkanes by recombinant alkane hydroxylase (AlkB) and rubredoxin-2 (AlkG) from Pseudomonas putida GPo1. Sci. Rep. 2017, 7, 8369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tani, A.; Ishige, T.; Sakai, Y.; Kato, N. Gene structures and regulation of the alkane hydroxylase complex in Acinetobacter sp. Strain M-1. J. Bacteriol. 2001, 183, 1819–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Liu, X.; Yang, W.; Xu, F.; Wang, W.; Feng, L.; Bartlam, M.; Wang, L.; Rao, Z. Crystal structure of long-chain alkane monooxygenase (LadA) in complex with coenzyme FMN: Unveiling the long-chain alkane hydroxylase. J. Mol. Biol. 2008, 376, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wang, L.; Feng, L. Characterization of a novel Rieske-type alkane monooxygenase system in Pusillimonas sp. strain T7-7. J. Bacteriol. 2013, 195, 1892–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Shao, Z. Diversity of flavin-binding monooxygenase genes (almA) in marine bacteria capable of degradation long-chain alkanes. Fems Microbiol. Ecol. 2012, 80, 523–533. [Google Scholar] [CrossRef] [Green Version]
- Lyu, Y.; Zheng, W.; Zheng, T.; Tian, Y. Biodegradation of polycyclic aromatic hydrocarbons by Novosphingobium pentaromativorans US6-1. PLoS ONE 2014, 9, e101438. [Google Scholar] [CrossRef]
- Chen, X.; Tang, H.; Liu, Y.; Xu, P.; Xue, Y.; Lin, K.; Cui, C. Purification and initial characterization of 3-hydroxybenzoate 6-hydroxylase from a halophilic Martelella Strain AD-3. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.-R. Comprehensive Handbook of Chemical Bond Energies; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Peng, T.; Luo, A.; Kan, J.; Liang, L.; Huang, T.; Hu, Z. Identification of a ring-hydroxylating dioxygenases capable of anthracene and benz[a]anthracene oxidization from Rhodococcus sp. P14. J. Mol. Microbiol. Biotechnol. 2018, 28, 183–189. [Google Scholar] [CrossRef]
- Fu, B.; Xu, T.; Cui, Z.; Ng, H.L.; Wang, K.; Li, J.; Li, Q.X. Mutation of phenylalanine-223 to leucine enhances transformation of benzo[a]pyrene by ring-hydroxylating dioxygenase of Sphingobium sp. FB3 by increasing accessibility of the catalytic site. J. Agric. Food Chem. 2018, 66, 1206–1213. [Google Scholar] [CrossRef]
- Patrauchan, M.A.; Florizone, C.; Eapen, S.; Gómez-Gil, L.; Sethuraman, B.; Fukuda, M.; Davies, J.; Mohn, W.W.; Eltis, L.D. Roles of ring-hydroxylating dioxygenases in styrene and benzene catabolism in Rhodococcus jostii RHA1. J. Bacteriol. 2008, 190, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Zhu, Q.; Wu, Y.; Chen, H.; Lin, X. Characterization of a polycyclic aromatic ring-hydroxylation dioxygenase from Mycobacterium sp. NJS-P. Chemosphere 2017, 185, 67–74. [Google Scholar] [CrossRef]
- Lu, Q.; Chen, K.; Long, Y.; Liang, X.; He, B.; Yu, L.; Ye, J. Benzo(a)pyrene degradation by cytochrome P450 hydroxylase and the functional metabolism network of Bacillus thuringiensis. J. Hazard. Mater. 2019, 366, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Mao, G.; Wang, Y.; Bartlam, M. Structural insights into diversity and n-alkane biodegradation mechanisms of alkane hydroxylases. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, A.; Chatterjee, K.; Madras, G. Enzymatic degradation of polymers: A brief review. Mater. Sci. Technol. 2014, 30, 567–573. [Google Scholar] [CrossRef]
- Noble, M.A.; Miles, C.S.; Chapman, S.K.; Lysek, D.A.; Mackay, A.C.; Reid, G.A.; Hanzlik, R.P.; Munro, A.W. Roles of key active-site residues in flavocytochrome P450 BM3. Biochem. J. 1999, 339, 371–379. [Google Scholar] [CrossRef]
- Schrewe, M.; Magnusson, A.O.; Willrodt, C.; Bühler, B.; Schmid, A. Kinetic analysis of terminal and unactivated C-H bond oxyfunctionalization in fatty acid methyl esters by monooxygenase-based whole-cell biocatalysis. Adv. Synth. Catal. 2011, 353, 3485–3495. [Google Scholar] [CrossRef]
- Schäfer, L.; Karande, R.; Bühler, B. Maximizing biocatalytic cyclohexane hydroxylation by modulating cytochrome P450 monooxygenase expression in P. taiwanensis VLB120. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Liu, J. Evaluating bacterial community structures in oil collected from the sea surface and sediment in the northern Gulf of Mexico after the Deepwater Horizon oil spill. MicrobiologyOpen 2013, 2, 492–504. [Google Scholar] [CrossRef]
- Lu, C.; Hong, Y.; Liu, J.; Gao, Y.; Ma, Z.; Yang, B.; Ling, W.; Waigi, M.G. A PAH-degrading bacterial community enriched with contaminated agricultural soil and its utility for microbial bioremediation. Environ. Pollut. 2019, 251, 773–782. [Google Scholar] [CrossRef]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum hydrocarbon-degrading bacteria for the remediation of oil pollution under aerobic conditions: A perspective analysis. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pure or Mixture Species | Microorganisms | Potential Enzymes | Reference |
|---|---|---|---|
| Pure species (two) | Bacillus cereus and Bacillus gottheilii | N/A 1 | [10] |
| Pure species | Cupriavidus necator H16 | N/A | [20] |
| Microbial consortium from superworms’ guts | Alcaligenes sp., Pseudomonas sp., or Acinetobacter sp., and Klebsiella pneumoniae | N/A | [28] |
| Microbial consortium from an open waste dump | Shingobacterium sp., Flavobacterium spp., Pseudoxanthomonas sp., Burkholderia sp., Xanthobacter sp., Methylobacter sp., Methylococcus sp., Methylocella sp., Methylocystis sp., Nitrobacter hamburgensis, Nitrobacter wingogradskyi and Nitrosomonas sp. | N/A | [6] |
| Pure species | Penicillium variabile CCF3219 | N/A | [29] |
| Pure species (two) | Pseudomonas spp. and Bacillus spp. | Lipase and esterase | [21] |
| Pure species (four) | Enterobacter sp., Citrobacter sedlakii, Alcaligenes sp. and Brevundimonas diminuta | Extracellular deploymerase enzymes | [30] |
| Pure species(three) | Serratia marcescens PCM3034, Klebsiella oxytoca PCM3036 and Pseudomonas aeruginosa PCM3035. | N/A | [31] |
| Pure species | Gloeophyllum trabeum DSM 1398 | Oxidative (exo)enzymes | [19] |
| Pure species | Exiguobacterium sp. YT2 | N/A | [25] |
| Pure species from superworms’ guts | Pseudomonas aeruginosa strain DSM 50071 | N/A | [32] |
| Microbial consortium from moths’ guts | Enterococcus sp., Geobacillus sp. Serratia marcescens, Pseudomonas sp. and Bacillus cereus. | N/A | [9] |
| Microbial consortium from seawater | N/A | Alkane 1-monooxygenase | [5] |
| Microbial consortium from mealworms’ guts | Citrobacter spp., Kosakonia spp., Listeria spp. and Nitrospira defluvii | β-Galactosidase, acid phosphatase, β-glucuronidase, naphthol-AS-BI-phosphohydrolase, leucine arylamidase, and alkaline phosphatase | [22] |
| Microbial consortium from mealworms’ guts | Listeria sp., Nitrospira defluvii, Pedomicrobium sp., Aquihabitans sp., unclassified Xanthomonadaceae, unclassified Saprospiraceae, and unclassified Burkholeriales | N/A | [33] |
| Microbial consortium from mealworms’ guts | Spiroplasmataceae, Enterococcaceae, and Enterobacteriaceae | N/A | [34] |
| Microbial consortium from superworms’ guts | N/A | N/A | [35] |
| N/A | N/A | P450 monooxygenases | [24] |
| Microbial consortium from brackish water | Burkholderiales | N/A | [36] |
| Pure species | Azotobacter beijerinckii HM121 | Hydroquinone peroxidase | [23] |
| Enrichment culture | Winogradskyella, Salinimicrobium, Vibrio, Photobacterium and Pseudomonas | N/A | [37] |
| Pure species | Rhodococcus ruber C208 | N/A | [38] |
| Pure species (two) | Exiguobacterium sibiricum strain DR11 and Exiguobacterium undae strain DR14 | Hydrolyzing enzymes | [39] |
| Kingdom | Phylum | Family | Genus | Species | Number of Hydrolases in Search Result | Number of Monooxygenases in Search Result |
|---|---|---|---|---|---|---|
| Bacteria | Firmicutes | Bacillaceae | Bacillus | Bacillus cereus ATCC 14579 | 290 | 24 |
| Firmicutes | Bacillaceae | Cytobacillus | Cytobacillus gottheilii ASM163634v1 | 156 | 11 | |
| Firmicutes | Enterococcaceae | Enterococcus | Enterococcus faecium DO | 177 | 7 | |
| Firmicutes | Listeriaceae | Listeria | Listeria innocua Clip11262 | 90 | 9 | |
| Firmicutes | Bacillales Family XII. Incertae Sedis | Exiguobacterium | Exiguobacterium sibiricum 255-15 | 74 | 11 | |
| Proteobacteria | Burkholderiaceae | Cupriavidus | Cupriavidus necator N-1 | 254 | 59 | |
| Proteobacteria | Burkholderiaceae | Burkholderia | Burkholderia cepacia ATCC 25416 | 292 | 53 | |
| Proteobacteria | Burkholderiaceae | Burkholderia | Burkholderia pseudomallei K96243 | 270 | 40 | |
| Proteobacteria | Enterobacteriaceae | Klebsiella | Klebsiella pneumoniae subsp. pneumoniae HS11286 | 193 | 34 | |
| Proteobacteria | Enterobacteriaceae | Kosakonia | Kosakonia radicincitans DSM 16656 | 132 | 19 | |
| Proteobacteria | Enterobacteriaceae | Cronobacter | Cronobacter sakazakii ASM98282v1 | 2662 * | 254 * | |
| Proteobacteria | Enterobacteriaceae | Klebsiella | Klebsiella oxytoca ASM102219v1 | 6807 * | 801 * | |
| Proteobacteria | Enterobacteriaceae | Citrobacter | Citrobacter sedlakii NBRC 105722 | 64 | 4 | |
| Proteobacteria | Alcaligenaceae | Alcaligenes | Alcaligenes sp. Strain HPC1271 | N/A | N/A | |
| Proteobacteria | Aeromonadales | Aeromonas | Aeromonas taiwanensis LMG 24683 | 129 | 14 | |
| Proteobacteria | Caulobacteraceae | Brevundimonas | Brevundimonas diminuta 48290_B02 | 633 | 59 | |
| Proteobacteria | Enterobacterales | Serratia | Serratia marcescens subsp. marcescens Db11 | 145 | 26 | |
| Proteobacteria | Pseudomonadaceae | Pseudomonas | Pseudomonas aeruginosa PAO1 | 471 | 90 | |
| Actinobacteria | Nocardiaceae | Rhodococcus | Rhodococcus ruber ASM274172v1 | 938 * | 270 * | |
| Eukaryota | Fungi | Gloeophllaceae | Gloeophyllum | Gloeophyllum trabeum | 234 | 21 |
| Fungi | Trichocomaceae | Talaromyces | Talaromyces islandicus | 49 | 29 |
| Selected Enzymes | Enzyme Class | Reaction Likely Catalyzed | Potential Carbon Targeted by Enzyme | Enzyme Ranking * | |
|---|---|---|---|---|---|
| Enzyme Family | Representative Subclass | ||||
| Cytochrome P450 | Oxidoreductases | Catalyzing the introduction of one atom of molecular oxygen into nonactivated C-H bonds. | β-carbon | High | |
| Monooxygenase | Alkane monooxygenase, 4-hydroxybenzoate 3-monooxygenase, 3-hydroxybenzoate 6-monooxygenase | Oxidoreductases | Incorporating one atom of the oxygen molecule into substrates. | β-carbon | High |
| Aromatic ring hydroxylase | 2-octaprenyl-6-methoxyphenol hydroxylase, 2-octaprenyl-3-methyl-6-methoxy-1,4-benzoquinol hydroxylase, aromatic ring hydroxylating dioxygenase | Oxidoreductases | Incorporating two atoms of dioxygen into the aromatic ring with the dihydroxylation reaction. | U-ring-carbon | Moderate |
| Esterase | Hydroxylase | Splitting esters into an acid and an alcohol. | Esters | Low | |
| Alpha/beta hydrolase | Hydroxylase | Diverse catalytic functions including hydrolysis, proteolysis, removal of a halogen atom, etc. | Ester and peptide bonds. | Low | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, L.; Majumder, E.L.-W. Potential for and Distribution of Enzymatic Biodegradation of Polystyrene by Environmental Microorganisms. Materials 2021, 14, 503. https://doi.org/10.3390/ma14030503
Hou L, Majumder EL-W. Potential for and Distribution of Enzymatic Biodegradation of Polystyrene by Environmental Microorganisms. Materials. 2021; 14(3):503. https://doi.org/10.3390/ma14030503
Chicago/Turabian StyleHou, Liyuan, and Erica L.-W. Majumder. 2021. "Potential for and Distribution of Enzymatic Biodegradation of Polystyrene by Environmental Microorganisms" Materials 14, no. 3: 503. https://doi.org/10.3390/ma14030503
APA StyleHou, L., & Majumder, E. L. -W. (2021). Potential for and Distribution of Enzymatic Biodegradation of Polystyrene by Environmental Microorganisms. Materials, 14(3), 503. https://doi.org/10.3390/ma14030503