
Decellularized Avian Cartilage, a Promising Alternative for Human Cartilage Tissue Regeneration

Abstract
:1. Introduction
2. Results
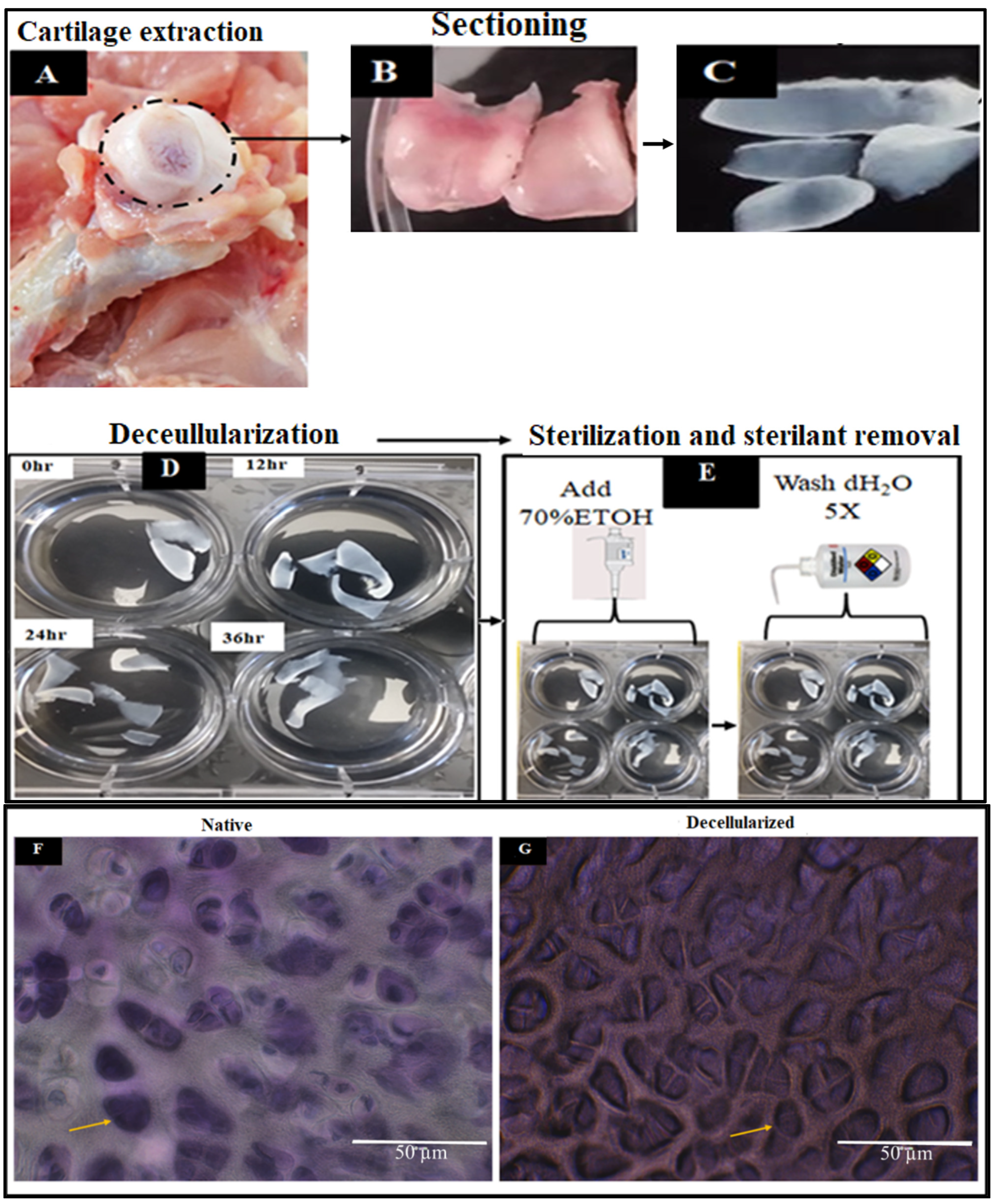
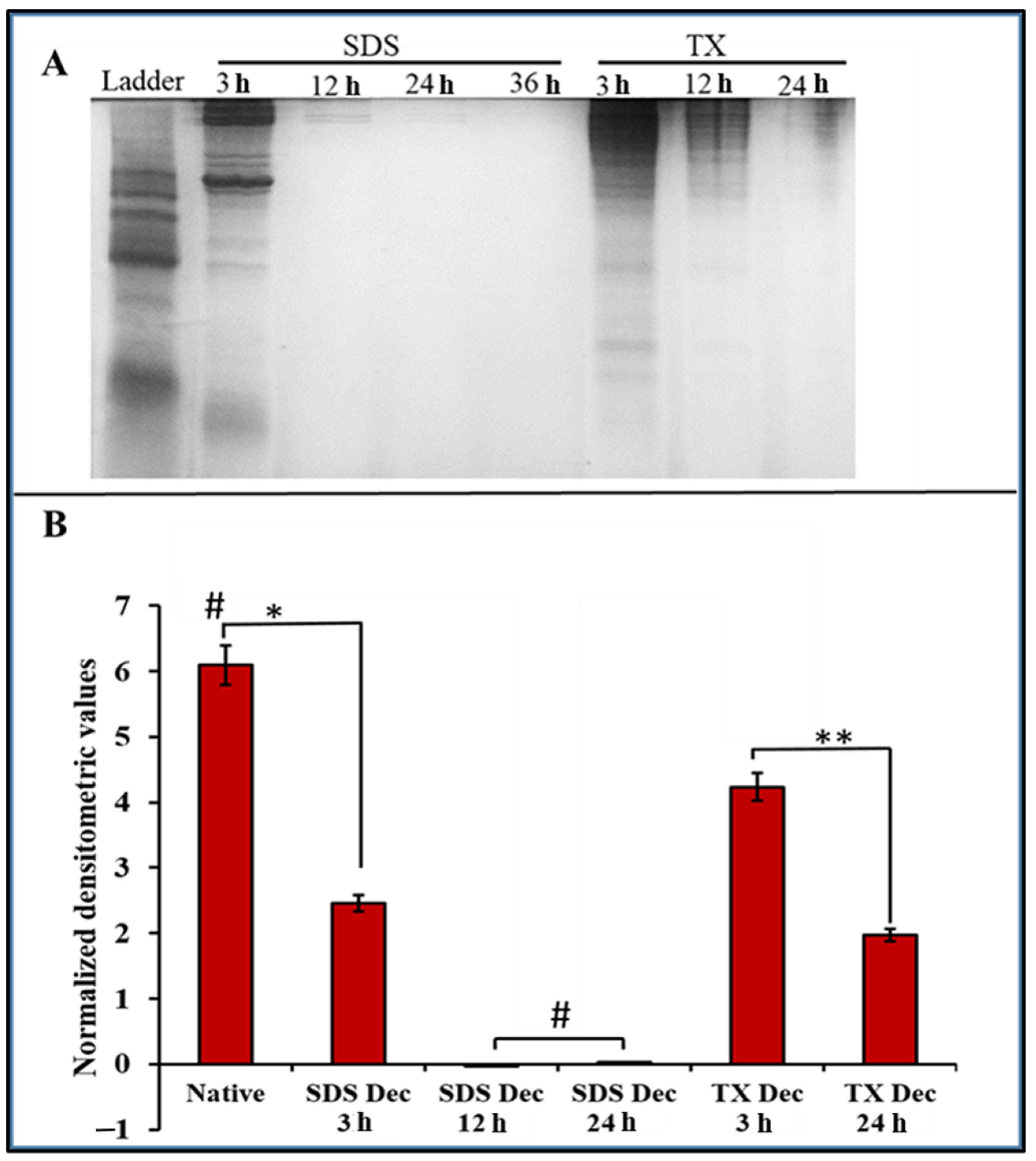
2.1. Extraction and Decellularization of Avian Articular Cartilage
2.1.1. Scanning Electron Microscopy
2.1.2. Physical Characteristics of Decellularized Cartilages
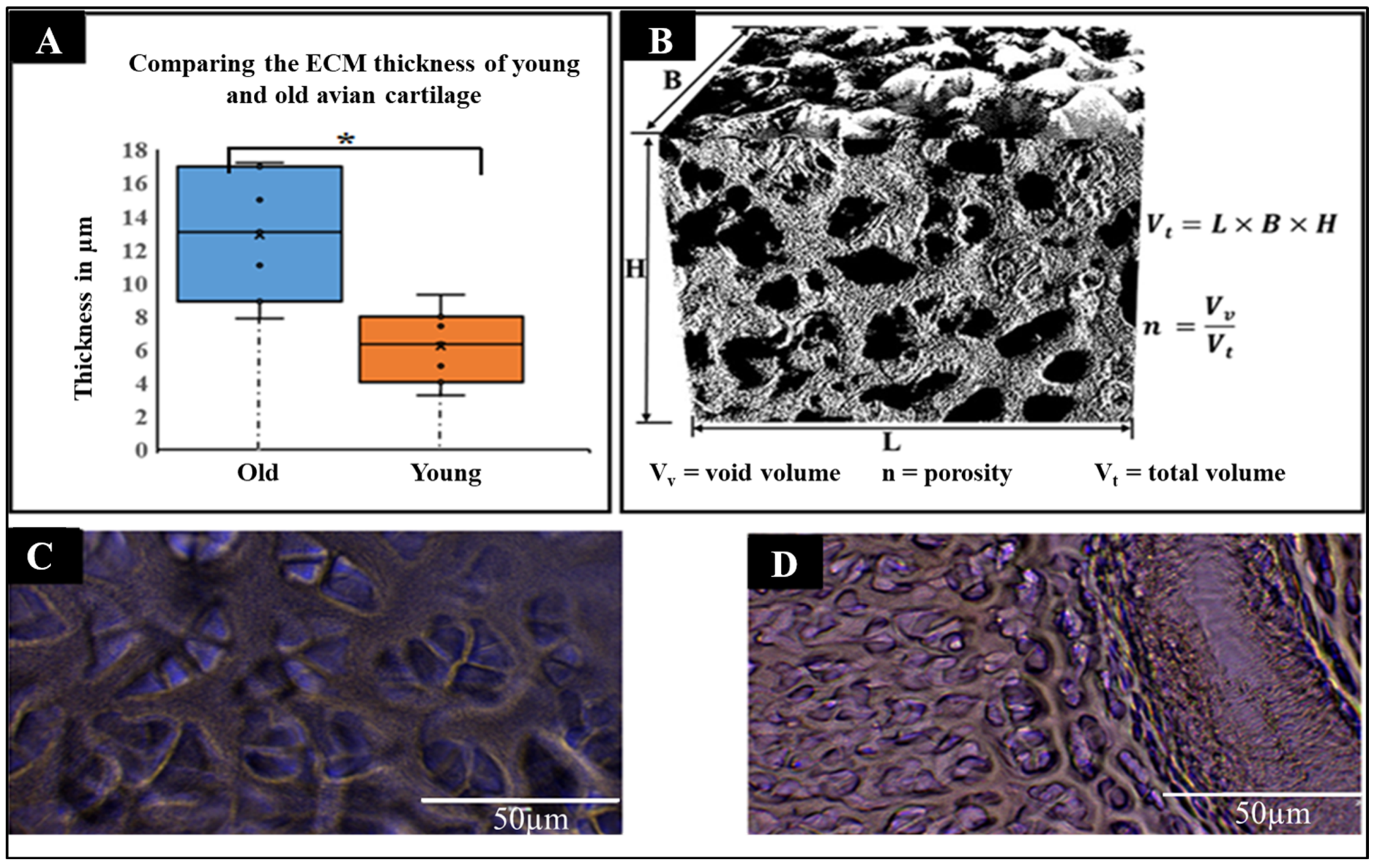
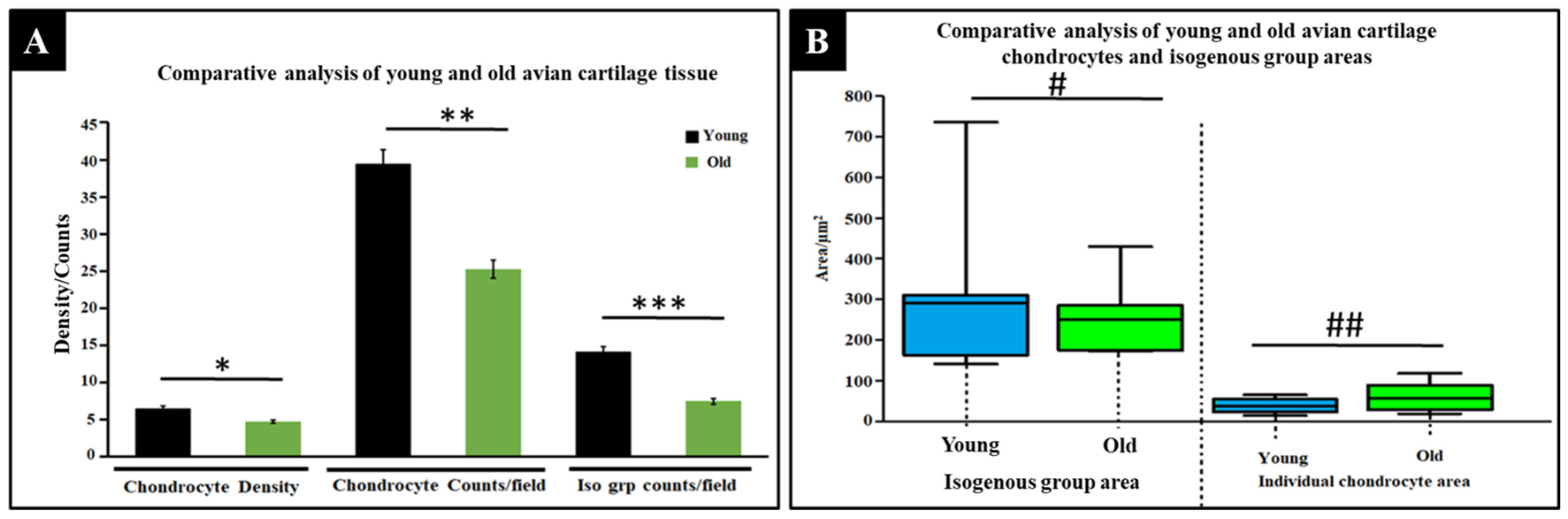
2.1.3. Comparative Physical Analysis of Young and Old Avian Cartilage Tissue
2.2. Protein Assessment of DACS
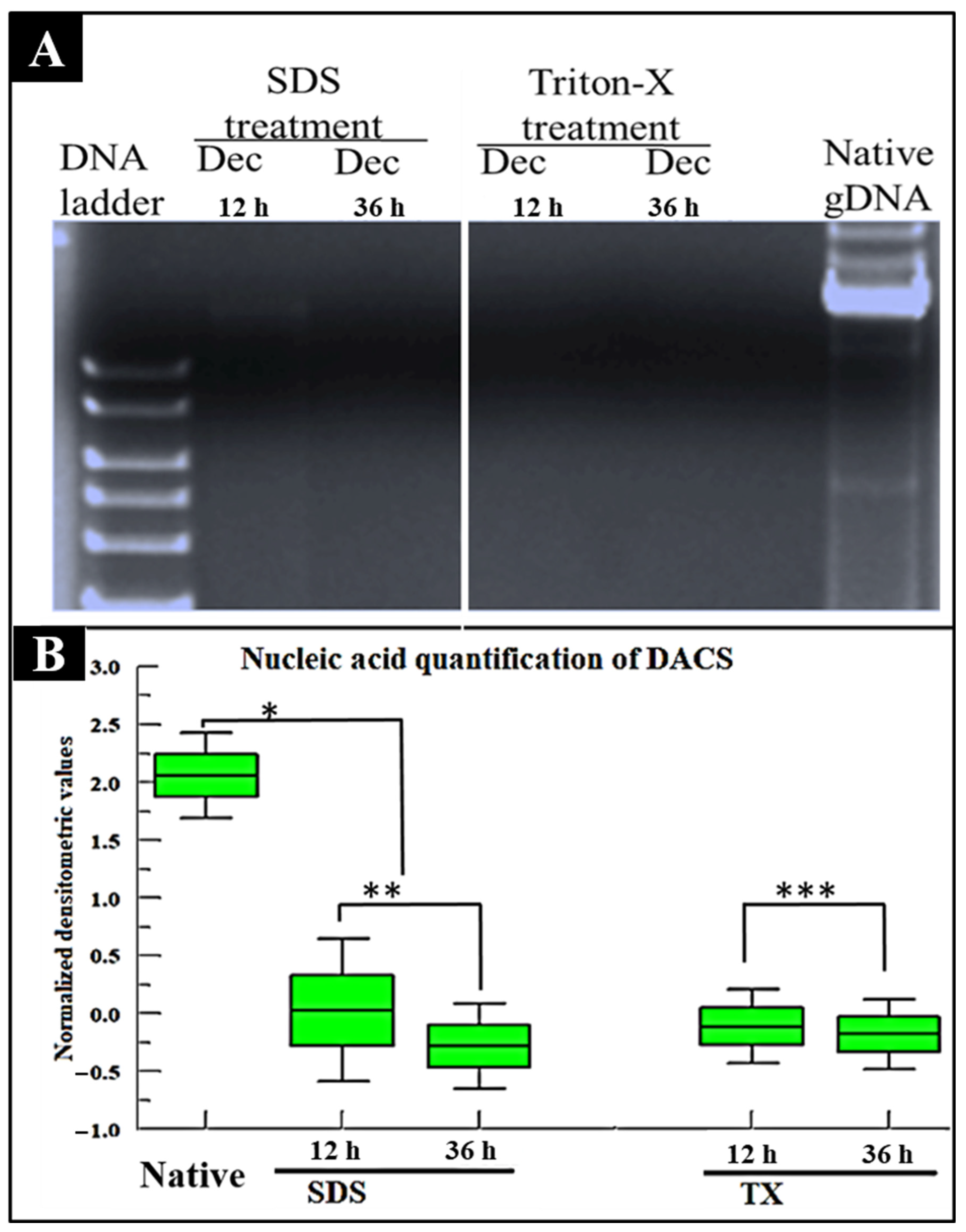
2.3. Nucleic Acid Assessment of DACS
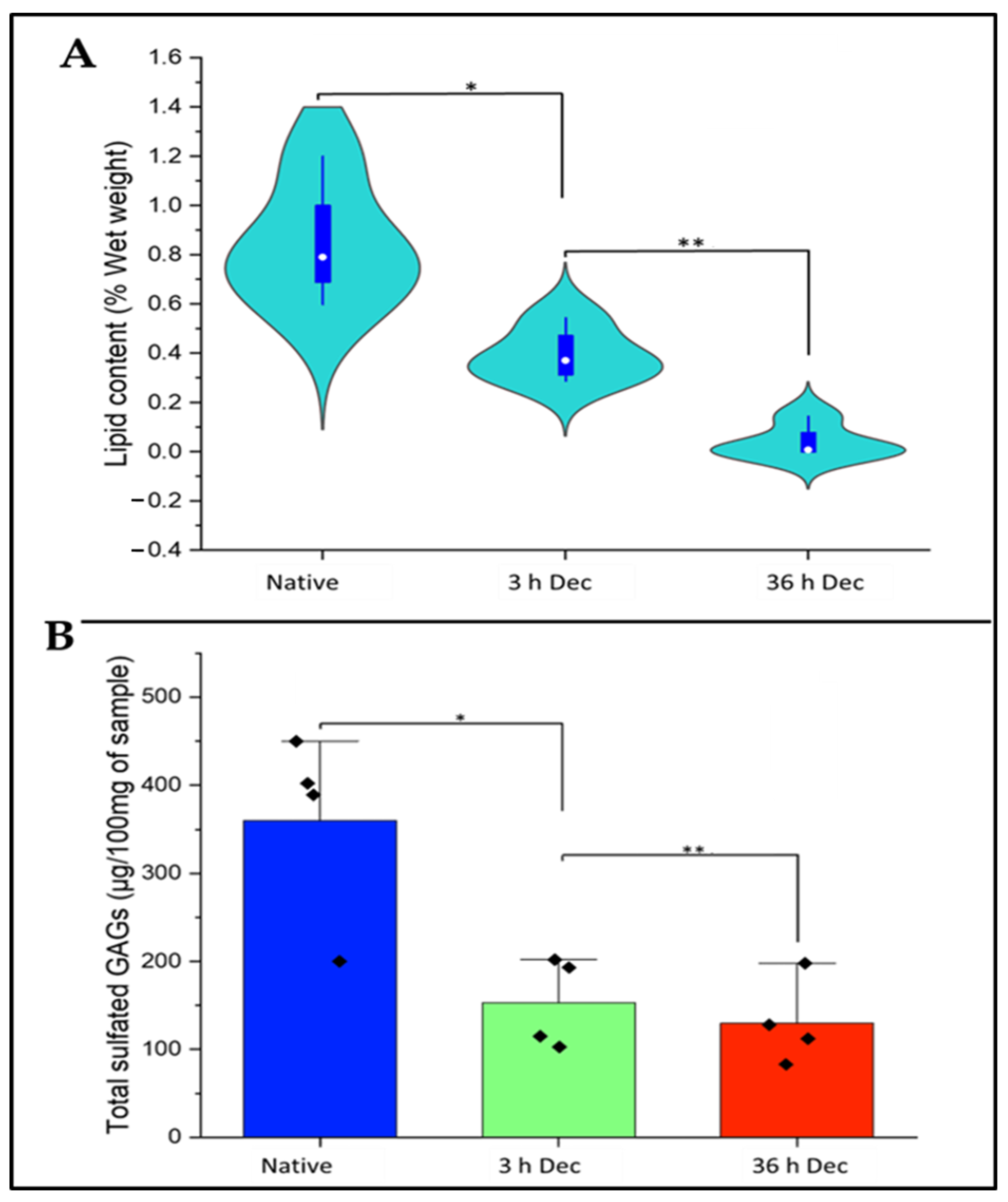
2.4. Analysis of Lipid Content of DACS
2.5. Analysis of Sulfated Sgags Content of DACS
2.6. Histochemical Analysis
2.6.1. DACS Lipid Content
2.6.2. DACS Sulfated Gags (Sgags) Content
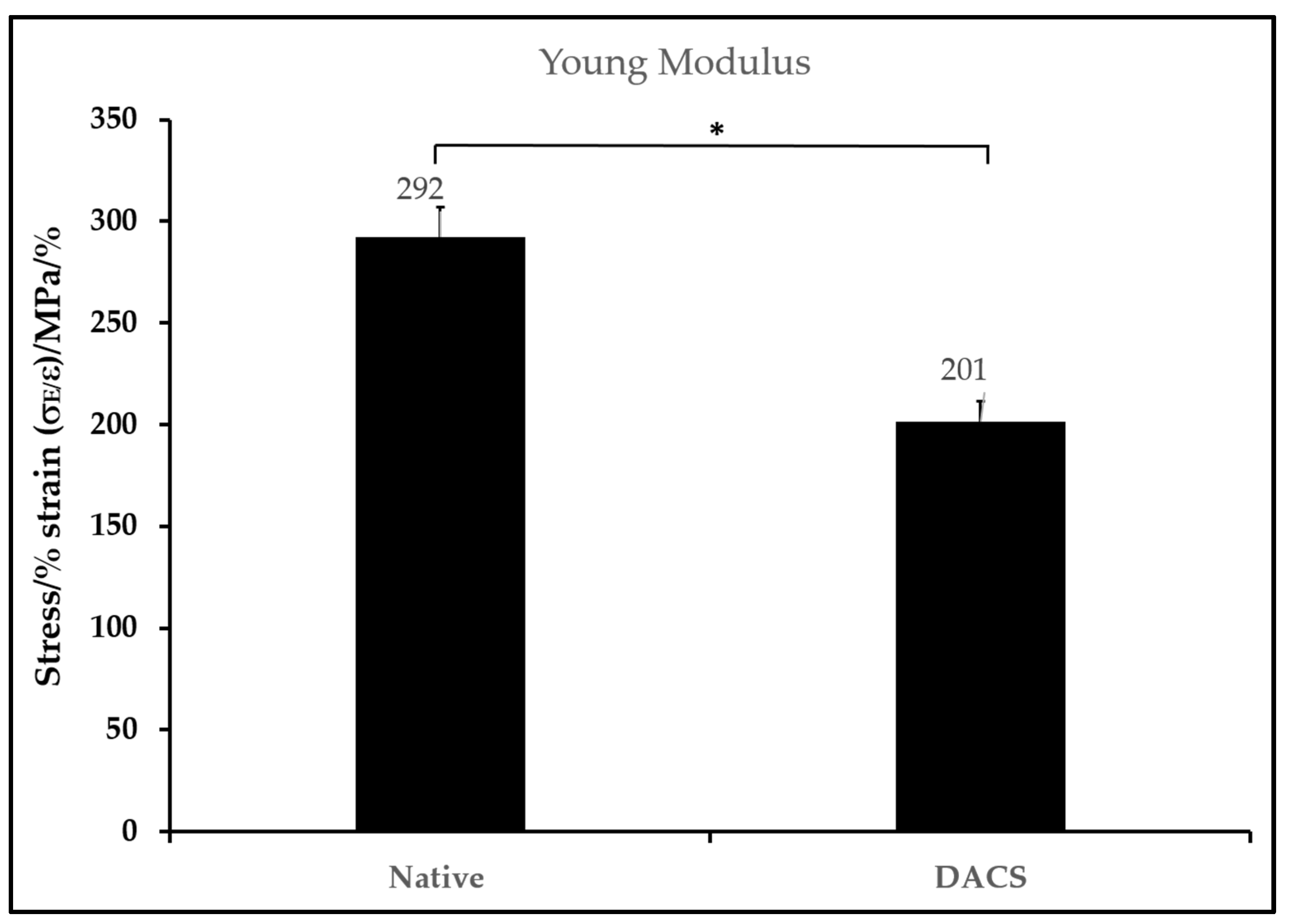
2.7. Biomechanical Characteristics of DACS
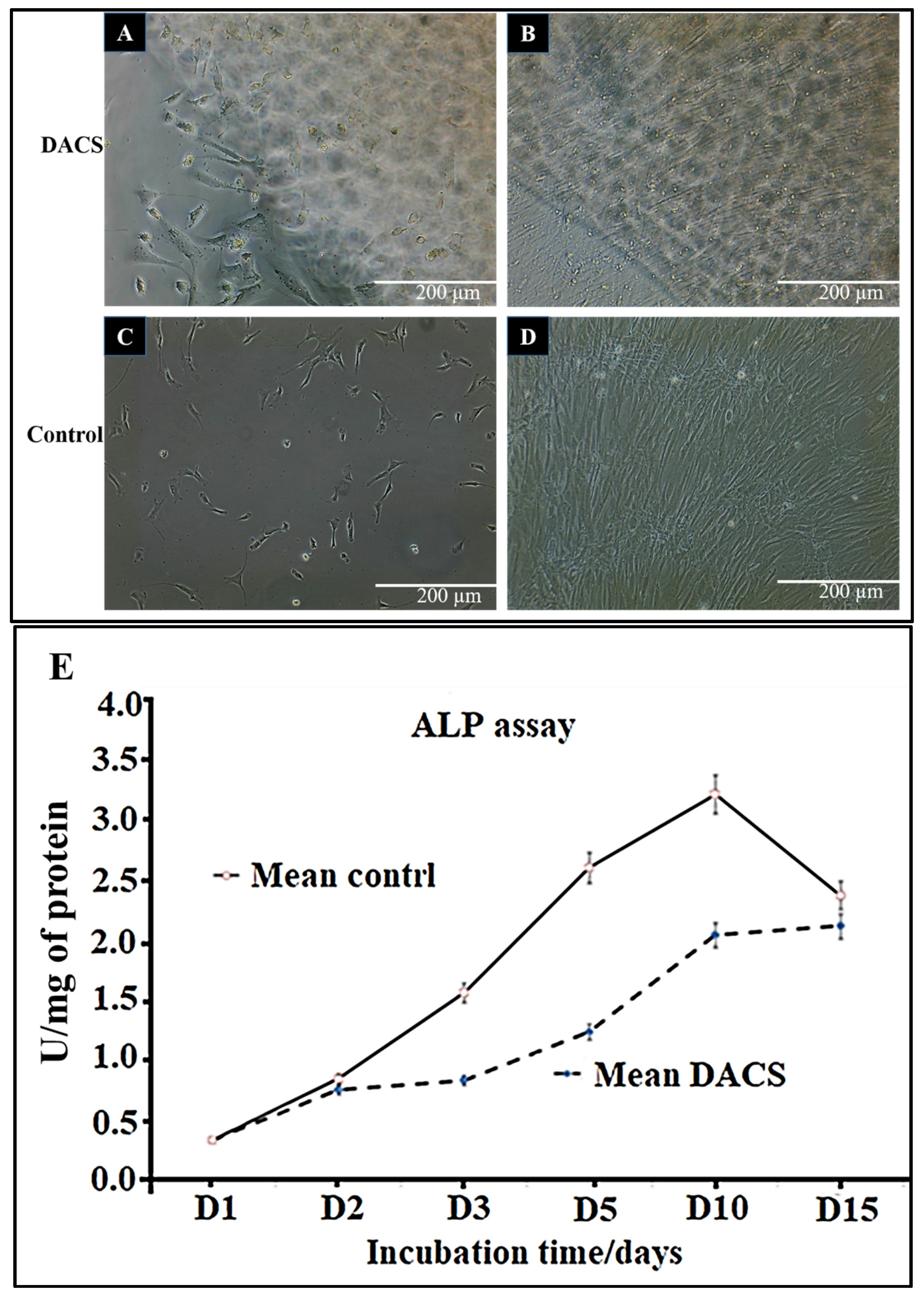
2.8. Chondrocyte Viabilities on Monolayer Plate and on DACS
2.9. Alkaline Phosphatase Activity Analysis of Chondrocyte Lysates from Monolayer Control Plates and Cells Seeded on DACS
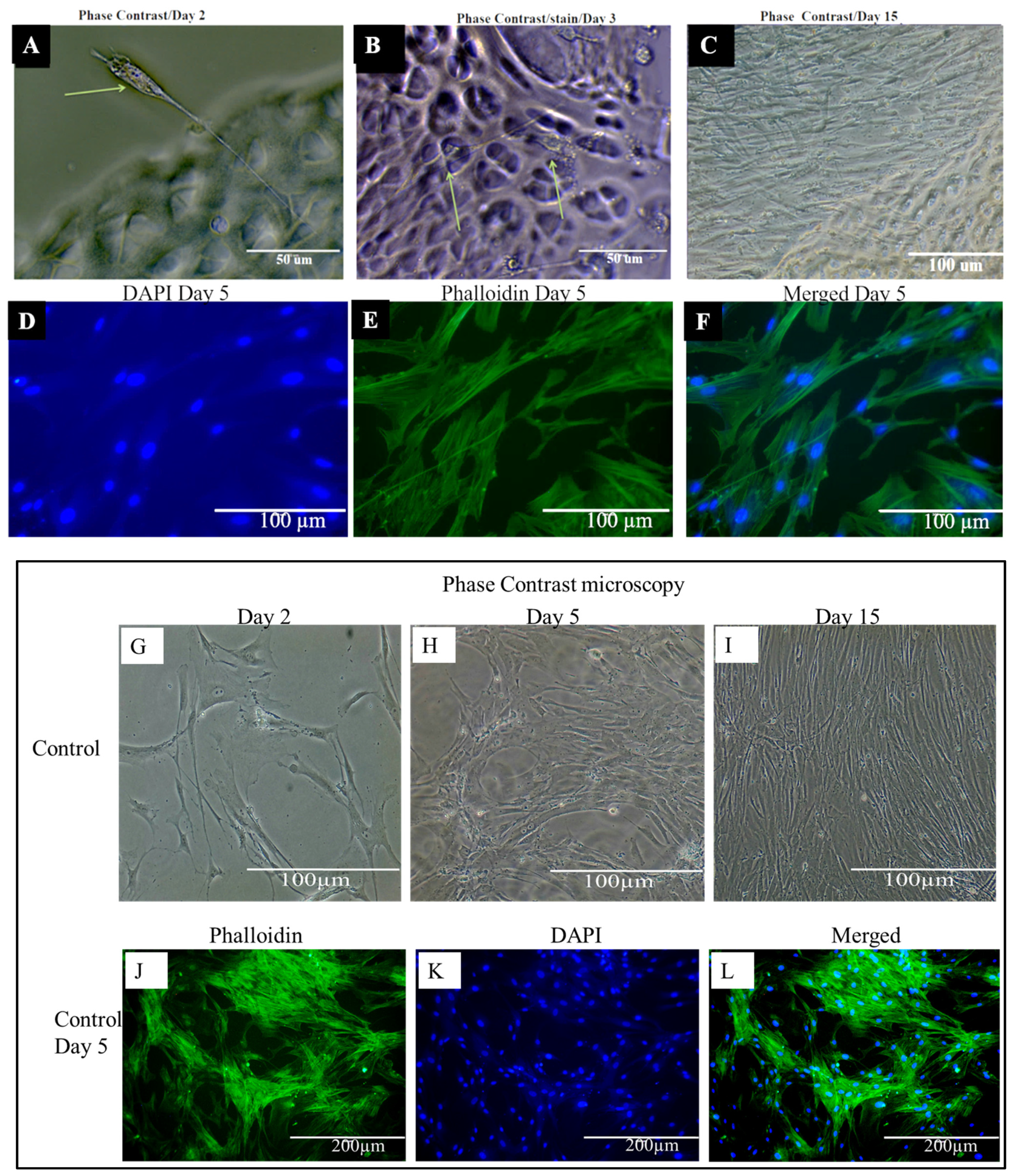
2.10. Phenotypic Characteristics of Chondrocytes Seeded on DACS
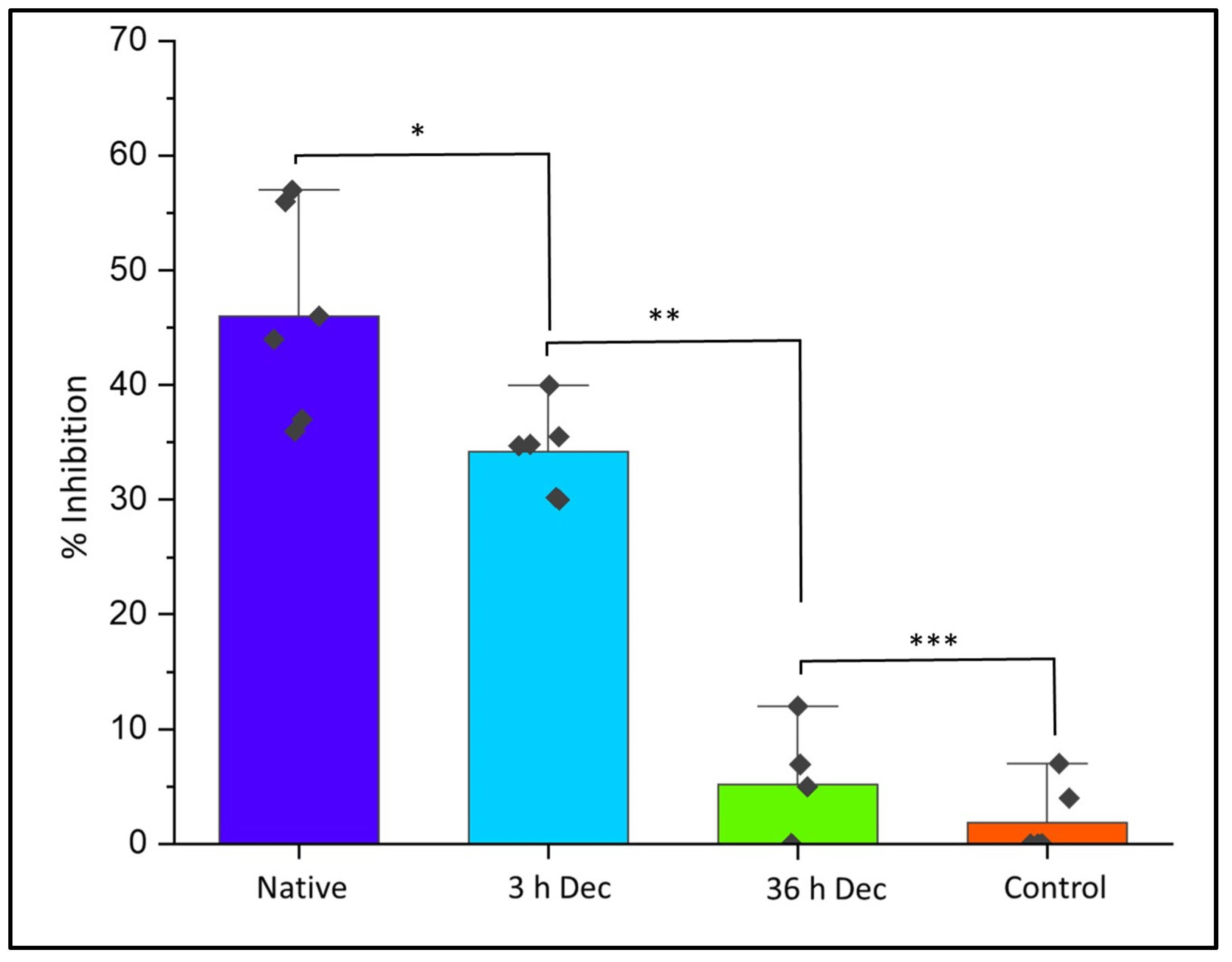
2.11. Evaluation of Cytotoxicity of DACS Scaffold at Different Time Points of Decellularization
2.11.1. Qualitative Analysis of Cytotoxicity of DACS on Chondrocytes
2.11.2. Quantitative Analysis of Cytotoxicity of DACS on Chondrocytes
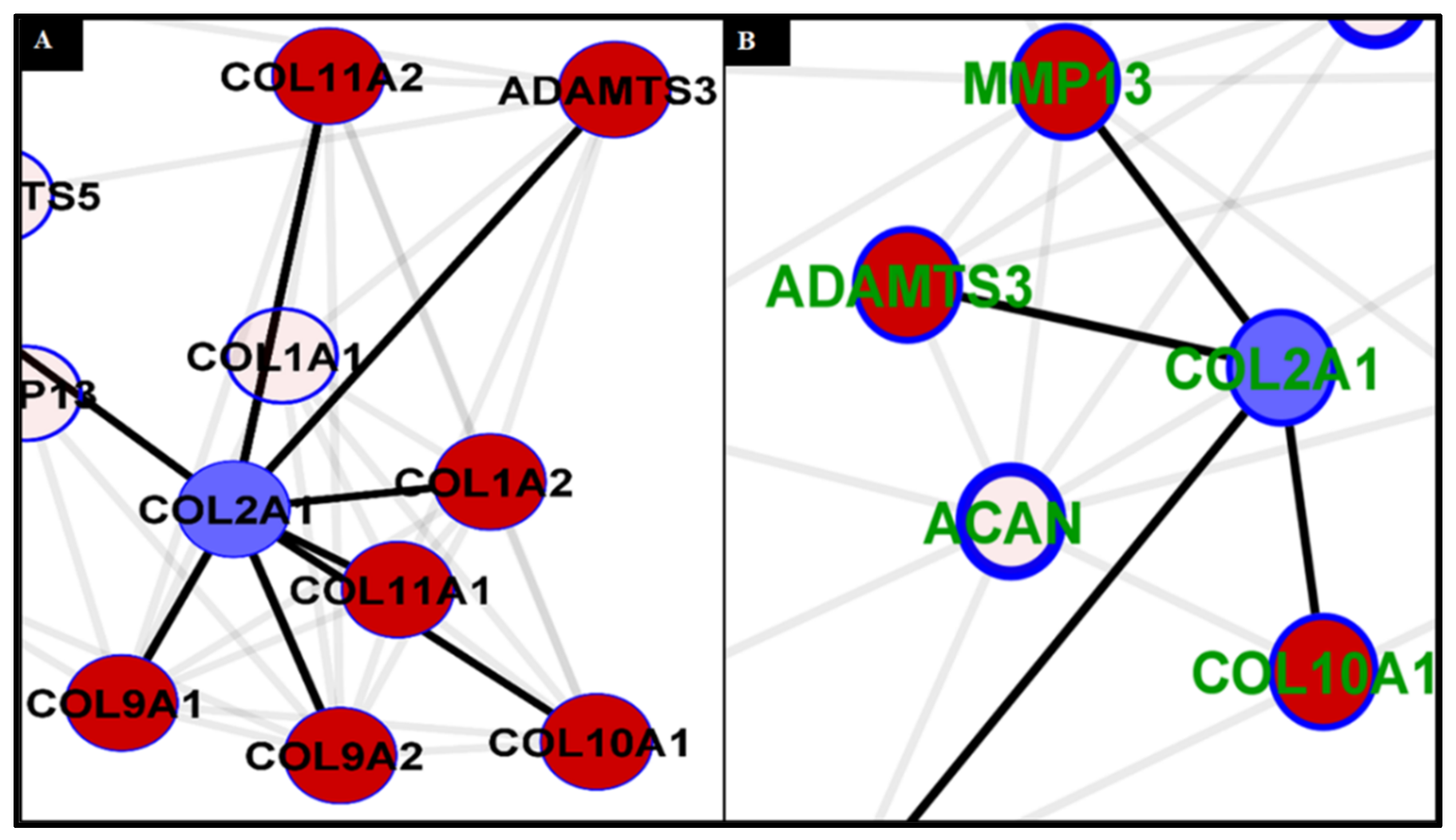
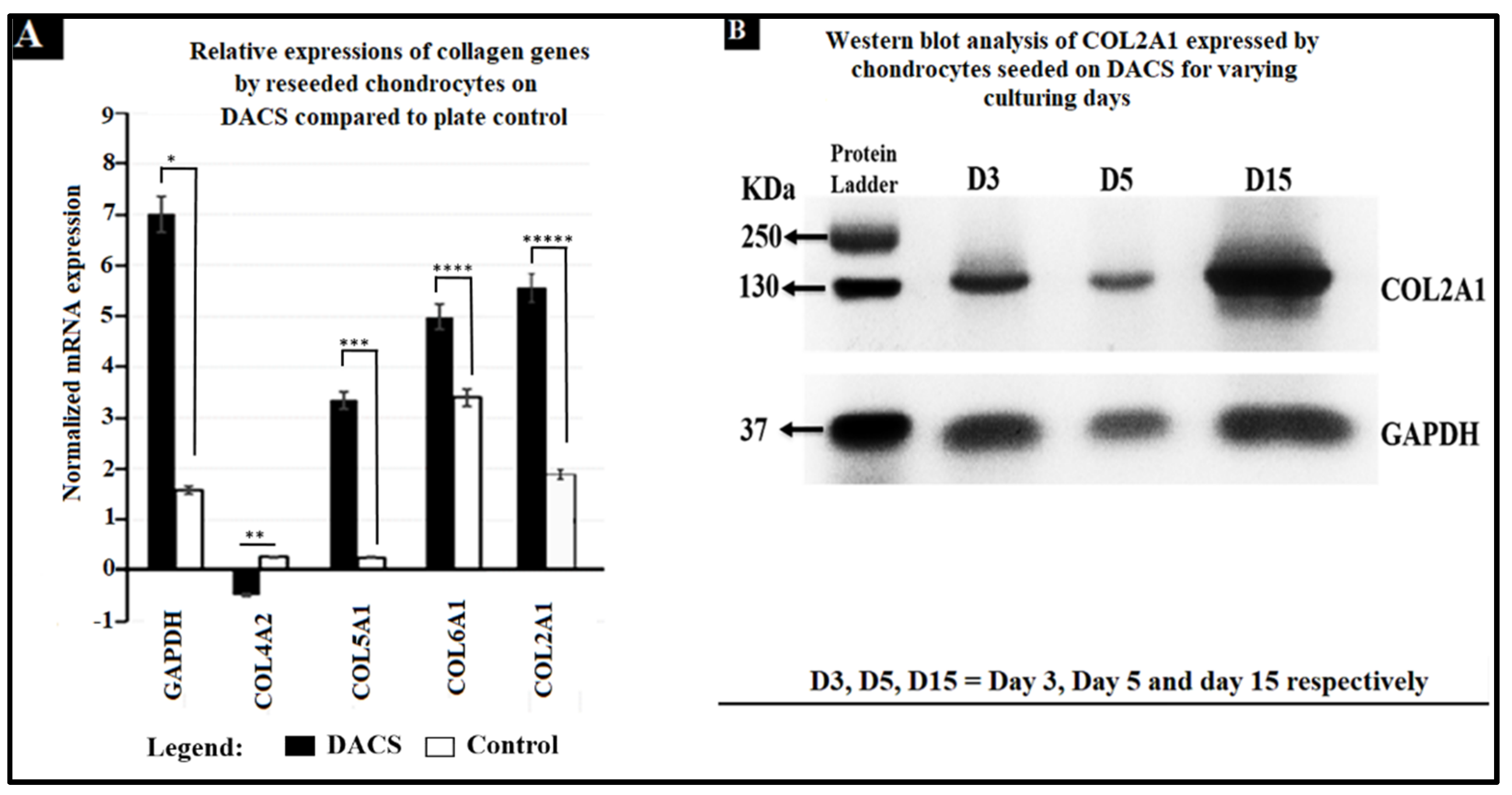
2.12. Analysis of Collagen Expression
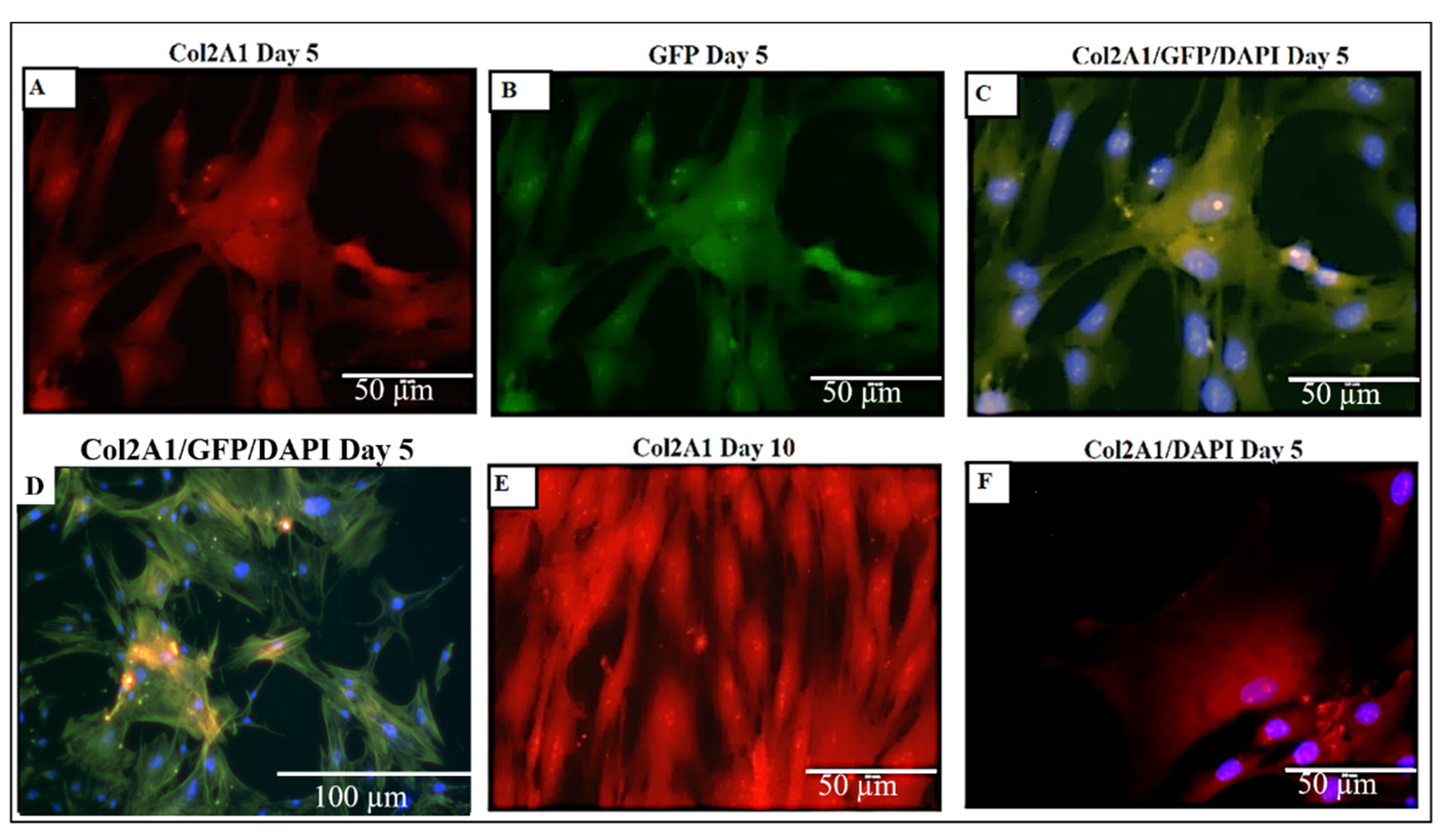
2.13. Higher Expression and Deposition of Col2A1 in DACS than Monolayer Control
3. Discussion
4. Materials and Methods
4.1. Decellularization of Avian Articular Cartilage as Scaffold (DACS)
4.1.1. Cryosectioning of Decellularized Avian Articular Cartilage
4.1.2. Decellularization of Sectioned Cartilages
4.1.3. Scanning Electron Microscopy (SEM)
4.2. Lipid Extraction and Quantification
4.3. Sulfated GAGs Extraction and Quantification
4.4. Seeding of Chondrocytes on DACS Scaffold
4.5. Protein and Nucleic Acid Assessment of DACS
4.5.1. Protein Assessment of DACS
4.5.2. Nucleic Acid Assessment of DACS
4.6. Histochemistry
4.7. In Vitro Assay of DACS Biocompatibility via the Use of Alamar Blue for Cell Inhibition
4.8. Alkaline Phosphatase (ALP) Assay for Chondrocytic Proliferation
4.9. Gene Expression Analysis
4.10. Immunofluorescence Assay of Chondrocytes Seeded on Scaffolds
4.11. SDS-PAGE and Western Blotting
4.12. Image Acquisition and Processing
Pore Size Analysis
4.13. Biomechanical Behavior Testing
4.14. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- More, N.; Kapusetti, G. Piezoelectric material—A promising approach for bone and cartilage regeneration. Med. Hypotheses 2017, 108, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Bedi, A.; Feeley, T.B.; Williams, R.J. Management of Articular Cartilage Defects of the Knee. J. Bone Jt. Surg. 2010, 92, 994–1009. [Google Scholar] [CrossRef] [PubMed]
- Green, J.D.; Tollemar, V.; Dougherty, M.; Yan, Z.; Yin, L.; Ye, J.; Collier, Z.; Mohammed, M.K.; Haydon, R.C.; Luu, H.H.; et al. Multifaceted signaling regulators of chondrogenesis: Implications in cartilage regeneration and tissue engineering. Genes Dis. 2015, 4, 307–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.; Rim, Y.A.; Lee, J.; Ju, J.H. Current therapeutic strategies for stem cell-based cartilage regeneration. Stem Cells Int. 2018, 2018, 8490489. [Google Scholar] [CrossRef] [Green Version]
- Koelling, S.; Miosge, N. Stem cell therapy for cartilage regeneration in osteoarthritis. Expert Opin. Biol. Ther. 2009, 9, 1399–1405. [Google Scholar] [CrossRef]
- Lee, K.B.; Hui, J.H.; Song, I.C.; Ardany, L.; Lee, E.H. Injectable mesenchymal stem cell therapy for large cartilage defects—A porcine model. Stem Cells 2007, 25, 2964–2971. [Google Scholar] [CrossRef] [Green Version]
- Mardones, R.; Jofré, C.M.; Minguell, J.J. Cell therapy and tissue engineering approaches for cartilage repair and/or regeneration. Int. J. Stem Cells 2015, 8, 48. [Google Scholar] [CrossRef]
- Frank, R.M.; Cotter, E.J.; Nassar, I.; Cole, B. Failure of bone marrow stimulation techniques. Sports Med. Arthrosc. Rev. 2017, 25, 2–9. [Google Scholar] [CrossRef]
- Young, A.; Dixey, J.; Cox, N.; Davies, P.; Devlin, J.; Emery, P.; Gallivan, S.; Gough, A.; James, D.; Prouse, P.; et al. How does functional disability in early rheumatoid arthritis (RA) affect patients and their lives? Results of 5 years of follow-up in 732 patients from the Early RA Study (ERAS). Rheumatology 2000, 39, 603–611. [Google Scholar] [CrossRef] [Green Version]
- Mollon, B.; Kandel, R.; Chahal, J.; Theodoropoulos, J. The clinical status of cartilage tissue regeneration in humans. Osteoarthr. Cartil. 2013, 21, 1824–1833. [Google Scholar] [CrossRef] [Green Version]
- Nerem, R.M. Tissue engineering: The hope, the hype, and the future. Tissue Eng. 2006, 12, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Mir, T.A.; Iwanaga, S.; Kurooka, T.; Toda, H.; Sakai, S.; Nakamura, M. Biofabrication offers future hope for tackling various obstacles and challenges in tissue engineering and regenerative medicine: A Perspective. Int. J. Bioprinting 2019, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Zafar, M.S.; Zohaib, K.; Khalid, A. Oral tissue engineering progress and challenges. Tissue Eng. Regen. Med. 2015, 12, 387–397. [Google Scholar] [CrossRef]
- Jones, C.W.; Smolinski, D.; Keogh, A.; Kirk, T.B.; Zheng, M.H. Confocal laser scanning microscopy in orthopaedic research. Prog. Histochem. Cytochem. 2005, 40, 1–71. [Google Scholar] [CrossRef] [PubMed]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The Basic Science of Articular Cartilage: Structure, Composition, and Function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Choi, K.H.; Choi, B.H.; Park, S.R.; Kim, B.J.; Min, B.H. The chondrogenic differentiation of mesenchymal stem cells on an extracellular matrix scaffold derived from porcine chondrocytes. Biomaterials 2010, 31, 5355–5365. [Google Scholar] [CrossRef]
- Sutherland, A.J.; Converse, G.L.; Hopkins, R.A.; Detamore, M.S. The bioactivity of cartilage extracellular matrix in articular cartilage regeneration. Adv. Healthc. Mater. 2015, 4, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Correa, D.; Lietman, S.A. Articular cartilage repair: Current needs, methods and research directions. Semin. Cell Dev. Biol. 2017, 62, 67–77. [Google Scholar] [CrossRef]
- Roessler, P.P.; Pfister, B.; Gesslein, M.; Figiel, J.; Heyse, T.J.; Colcuc, C.; Lorbach, O.; Efe, T.; Schüttler, K.F. Short-Term Follow up after Implantation of a Cell-Free Collagen Type I Matrix for the Treatment of Large Cartilage Defects of the Knee. Int. Orthop. 2015, 39, 2473–2479. [Google Scholar] [CrossRef]
- Schüttler, K.-F.; Götschenberg, A.; Klasan, A.; Stein, T.; Pehl, A.; Roessler, P.P.; Figiel, J.; Heyse, T.J.; Efe, T. Cell-Free Cartilage Repair in Large Defects of the Knee: Increased Failure Rate 5 Years after Implantation of a Collagen Type I Scaffold. Arch. Orthop. Trauma. Surg. 2019, 139, 99–106. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Afif, I.Y.; Maula, M.I.; Winarni, T.I.; Tauviqirrahman, M.; Akbar, I.; Basri, H.; van der Heide, E.; Jamari, J. Tresca Stress Simulation of Metal-on-Metal Total Hip Arthroplasty during Normal Walking Activity. Materials 2021, 14, 7554. [Google Scholar] [CrossRef] [PubMed]
- Jamari, J.; Ammarullah, M.I.; Saad, A.P.M.; Syahrom, A.; Uddin, M.; van der Heide, E.; Basri, H. The effect of bottom profile dimples on the femoral head on wear in metal-on-metal total hip arthroplasty. J. Funct. Biomater. 2021, 12, 38. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.H.; Kang, D.; Lim, M.W.; Kang, C.S.; Sung, H.J. Risk assessment of growth hormones and antimicrobial residues in meat. Toxicol. Res. 2010, 26, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.A.; Lee, P.F.; Barac, Y.; Hochman-Mendez, C.; Sampaio, L.C. Decellularization of whole hearts for cardiac regeneration. In Emerging Technologies for Heart Diseases; Academic Press: Cambridge, MA, USA, 2020; pp. 291–310. [Google Scholar]
- Zia, S.; Mozafari, M.; Natasha, G.; Tan, A.; Cui, Z.; Seifalian, A.M. Hearts beating through decellularized scaffolds: Whole-organ engineering for cardiac regeneration and transplantation. Crit. Rev. Biotechnol. 2016, 36, 705–715. [Google Scholar] [CrossRef]
- Schwarz, S.; Elsaesser, A.F.; Koerber, L.; Goldberg-Bockhorn, E.; Seitz, A.M.; Bermueller, C.; Durselen, L.; Ignatius, A.; Breiter, R.; Rotter, N. Processed xenogenic cartilage as innovative biomatrix for cartilage tissue engineering: Effects on chondrocyte differentiation and function. J. Tissue Eng. Regen. Med. 2012, 9, E239–E251. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Peng, J.; Lu, S.; Liu, S.; Zhang, L.; Huang, J.; Sui, X.; Zhao, B.; Wang, A.; Xu, W.; et al. In vivo cartilage repair using adipose-derived stem cell-loaded decellularized cartilage ECM scaffolds. J. Tissue Eng. Regen. Med. 2014, 8, 442–453. [Google Scholar] [CrossRef]
- Hench, L.L.; Jones, J.R. Artificial Organs and Tissue Engineering; Woodhead Publishing Ltd.: Cambridge, UK, 2005; pp. 275–284. [Google Scholar]
- Kim, B.S.; Das, S.; Jang, J.; Cho, D.W. Decellularized extracellular matrix-based bioinks for engineering tissue-and organ-specific microenvironments. Chem. Rev. 2020, 120, 10608–10661. [Google Scholar] [CrossRef]
- Kim, Y.S.; Majid, M.; Melchiorri, A.J.; Mikos, A.G. Applications of decellularized extracellular matrix in bone and cartilage tissue engineering. Bioeng. Transl. Med. 2019, 4, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Elena, G.; Roger, O.; Carolina, T.; Mateu, P.R.; Patricia, P.; Francisco, F.A.; Campistol, J.M.; Samitier, J.; Montserrat, N. Tissue engineering by decellularization and 3D bioprinting. Mater. Today 2017, 4, 66–178. [Google Scholar]
- Lim, T.; Tang, Q.; Zhu, Z.; Feng, Y.; Zhan, S.; Wei, X.; Zhang, C. A decellularized scaffold derived from squid cranial cartilage for use in cartilage tissue engineering. J. Mater. Chem. B 2020, 8, 4516. [Google Scholar] [CrossRef]
- Luo, Z.; Bian, Y.; Su, W.; Shi, L.; Li, S.; Song, Y.; Zheng, G.; Xie, A.; Xue, J. Comparison of various reagents for preparing a decellularized porcine cartilage scaffold. Am. J. Transl. Res. 2019, 11, 1417–1427. [Google Scholar] [PubMed]
- Sullivan, D.C.; Sayed-Hadi, M.S.; Daniel, B.D.; Pedro, M.B.; Tamer, A.; Anthony, A.; James, J.Y. Decellularization methods of porcine kidneys for whole organ engineering using a high-throughput system. Biomaterials 2012, 33, 7756–7764. [Google Scholar] [CrossRef] [PubMed]
- Burk, J.; Ina, E.; Dagmar, B.; Johannes, K.; Cornelia, K.; Bastian, P.; Karsten, W.; Walter, B. Freeze-thaw cycles enhance decellularization of large tendons. Tissue Eng. Part C Methods 2014, 20, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Nguyen, H.-Q.-D.; Weng, Y.-C. Characterization of porcine urinary bladder matrix hydrogels from sodium dodecyl sulfate decellularization method. Polymers 2020, 12, 3007. [Google Scholar] [CrossRef]
- Schaner, P.J.; Niels, D.M.; Thomas, N.T.; Irving, M.S.; Nicholas, A.T.; Rhoda, F.L.; Anthony, R.C.; Paul, J.D. Decellularized vein as a potential scaffold for vascular tissue engineering. J. Vasc. Surg. 2004, 40, 146–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temenoff, J.S.; Mikos, A.G. Tissue engineering for regeneration of articular cartilage. Biomaterials 2000, 21, 431–440. [Google Scholar] [CrossRef]
- Price, D. Legal and Ethical Aspects of Organ Transplantation; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Easterbrook, C.; Maddern, G. Porcine and bovine surgical products: Jewish, Muslim, and Hindu perspectives. Arch Surg. 2008, 143, 366–370. [Google Scholar] [CrossRef] [Green Version]
- Hussein, K.H.; Park, K.M.; Kang, K.S.; Woo, H.M. Biocompatibility evaluation of tissue-engineered decellularized scaffolds for biomedical application. Mater. Sci. Eng. C 2016, 67, 766–778. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, J.; Huang, X.; Fang, W.; Tao, Y.; Zhao, T.; Liang, C.; Hua, J.; Chen, Q.; Li, F. Injectable decellularized nucleus pulposus-based cell delivery system for differentiation of adipose-derived stem cells and nucleus pulposus regeneration. Acta Biomater. 2018, 81, 115–128. [Google Scholar] [CrossRef]
- Hussein, K.H.; Park, K.M.; Kim, H.M.; Teotia, P.K.; Ghim, J.H.; Woo, H.M. Construction of a biocompatible decellularized porcine hepatic lobe for liver bioengineering. Int. J. Artif. Organs 2015, 38, 96–104. [Google Scholar] [CrossRef]
- Fernández-Pérez, J.; Mark, A. The impact of decellularization methods on extracellular matrix derived hydrogels. Sci. Rep. 2019, 9, 14933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wess, T.J.; Wess, L.; Hocking, P.M. The structure of avian cartilage: A combined X-ray and biochemical analysis. J. Comp. Pathol. 1997, 116, 145–155. [Google Scholar] [CrossRef]
- Bielajew, B.J.; Hu, J.C.; Athanasiou, K.A. Collagen: Quantification, biomechanics and role of minor subtypes in cartilage. Nat. Rev. Mater. 2020, 5, 730–747. [Google Scholar] [CrossRef] [PubMed]
- Atia, A.J.; Azumah, A.D.; Deepa, B.; Dean, D. Tuning Phage for Cartilage Regeneration. In Bacteriophages; IntechOpen: London, UK, 2021. [Google Scholar]
- Somaiah, C.; Kumar, A.; Mawrie, D.; Sharma, A.; Patil, S.D.; Bhattacharyya, J.; Swaminathan, R.; Jaganathan, B.G. Collagen promotes higher adhesion, survival and proliferation of mesenchymal stem cells. PLoS ONE 2015, 10, e0145068. [Google Scholar]
- Xiang, J.X.; Zheng, X.L.; Gao, R.; Wu, W.Q.; Zhu, X.L.; Li, J.H.; Lv, Y. Liver regeneration using decellularized splenic scaffold: A novel approach in tissue engineering. Hepatobiliary Pancreat. Dis. Int. 2015, 14, 502–508. [Google Scholar] [CrossRef]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Wei, F.; Liu, S.; Chen, M.; Tian, G.; Zha, K.; Yang, Z.; Jiang, S.; Li, M.; Sui, X.; Chen, Z.; et al. Host Response to Biomaterials for Cartilage Tissue Engineering: Key to Remodeling. Front Bioeng Biotechnol. 2021, 9, 664592. [Google Scholar] [CrossRef]
- Boddupalli, A.; Zhu, L.; Bratlie, K.M. Methods for Implant Acceptance and Wound Healing: Material Selection and Implant Location Modulate Macrophage and Fibroblast Phenotypes. Adv. Heal. Mater. 2016, 5, 2575–2594. [Google Scholar] [CrossRef]
- Ayariga, J.A.; Huffman, A.M.; Napier, A.; Robertson, B.K.; Abugri, D. In silico Elucidation of Dihydroquinine Mechanism of Action against Toxoplasma gondii. bioRxiv 2022. [Google Scholar] [CrossRef]
- Huffman, A.M.; Ayariga, J.A.; Napier, A.; Robertson, B.K.; Abugri, D. Inhibition of Toxoplasma gondii growth by Dihydroquinine and its Mechanisms of Action. Front. Cell. Infect. Microbiol. 2022, 225. [Google Scholar] [CrossRef]
- Michler, R.E. Xenotransplantation: Risks, Clinical Potential, and Future Prospects. Emerg. Infect. Dis. 1996, 2, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucia, K.Y.C.; Victor, Y.L.L.; Vivian, T.; William, W.L.; Kenneth, M.C. Decellularized bovine intervertebral disc as a natural scaffold for xenogenic cell studies. Acta Biomater. 2013, 2, 5262–5272. [Google Scholar]
- Spaniol, K.; Witt, J.; Mertsch, S.; Borrelli, M.; Geerling, G.; Schrader, S. Generation and Characterisation of Decellularised Human Corneal Limbus. Graefes Arch. Clin. Exp. Ophthalmol. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 2018, 256, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Philips, C.; Campos, F.; Roosens, A.; del Carmen Sanchez-Quevedo, M.; Declercq, H.; Carriel, V. Qualitative and quantitative evaluation of a novel detergent-based method for decellularization of peripheral nerves. Ann. Biomed. Eng. 2018, 46, 1921–1937. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, N.; Gong, D.; Xia, C.; Xu, Z. Comparison of detergent-based decellularization protocols for the removal of antigenic cellular components in porcine aortic valve. Xenotransplantation 2018, 25, e12380. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, J.; Wu, Q.; Zhou, Y.; Li, Y.; Wu, X.; Shi, Y.; Li, L.; Bu, H. Method for perfusion decellularization of porcine whole liver and kidney for use as a scaffold for clinical-scale bioengineering engrafts. Xenotransplantation 2015, 22, 48–61. [Google Scholar] [CrossRef]
- Pelosi, P.; Rocco, P.R. Effects of mechanical ventilation on the extracellular matrix. Intensive Care Med. 2008, 34, 631–639. [Google Scholar] [CrossRef]
- Ibrahim, J.; Nguyen, A.H.; Rehman, A.; Ochi, A.; Jamal, M.; Graffeo, C.S.; Henning, J.R.; Zambirinis, C.P.; Fallon, N.C.; Barilla, R.; et al. Dendritic cell populations with different concentrations of lipid regulate tolerance and immunity in mouse and human liver. Gastroenterology 2012, 143, 1061–1072. [Google Scholar] [CrossRef] [Green Version]
- Golub, E.E.; Boesze-Battaglia, K. The role of alkaline phosphatase in mineralization. Curr. Opin. Orthop. 2007, 18, 444–448. [Google Scholar] [CrossRef]
- Gronthos, S.; Fitter, S.; Diamond, P.; Simmons, P.J.; Itescu, S.; Zannettino, A.C. A novel monoclonal antibody (STRO-3) iden-tifies an isoform of tissue nonspecific alkaline phosphatase expressed by multipotent bone marrow stromal Stem Cells. Stem Cells Dev. 2007, 16, 953–964. [Google Scholar] [CrossRef]
- Narisawa, S.; Yadav, M.C.; Millán, J.L. In vivo overexpression of tissue-nonspecific alkaline phosphatase increases skeletal mineralization and affects the phosphorylation status of osteopontin. J. Bone Miner. Res. 2013, 28, 1587–1598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, V. Alkaline phosphatase: Structure, expression and its function in bone mineralization. Gene 2020, 754, 144855. [Google Scholar]
- Whyte, M.P.; Landt, M.; Ryan, L.M.; Mulivor, R.A.; Henthorn, P.S.; Fedde, K.N.; Mahuren, J.D.; Coburn, S.P. Alkaline phosphatase: Placental and tissue-nonspecific isoenzymes hydrolyze phosphoethanolamine, inorganic pyrophosphate, and pyridoxal 5′-phosphate. Substrate accumulation in carriers of hypophosphatasia corrects during pregnancy. J. Clin. Investig. 1995, 95, 1440–1445. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Chi, G.; Li, P.; Lv, S.; Xu, J.; Xu, Z.; Xia, Y.; Tan, Y.; Xu, J.; Li, L.; et al. Effects of matrix stiffness on the morphology, adhesion, proliferation and osteogenic differentiation of mesenchymal Stem Cells. Int. J. Med. Sci. 2018, 15, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjipanayi, E.; Mudera, V.; Brown, R.A. Close dependence of fibroblast proliferation on collagen scaffold matrix stiffness. J. Tissue Eng. Regen. Med. 2009, 3, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Koerber, L.; Elsaesser, A.F.; Goldberg-Bockhorn, E.; Seitz, A.M.; Dürselen, L.; Ignatius, A.; Walther, P.; Breiter, R.; Rotter, N. Decellularized cartilage matrix as a novel biomatrix for cartilage tissue-engineering applications. Tissue Eng. Part A 2012, 18, 2195–2209. [Google Scholar] [CrossRef]
- Cukierman, E.; Pankov, R.; Stevens, D.R.; Yamada, K.M. Taking cell-matrix adhesions to the third dimension. Science 2001, 294, 1708–1712. [Google Scholar] [CrossRef] [PubMed]
- Sicchieri, L.G.; Crippa, G.E.; de Oliveira, P.T.; Beloti, M.M.; Rosa, A.L. Pore size regulates cell and tissue interactions with PLGA–CaP scaffolds used for bone engineering. J. Tissue Eng. Regen. Med. 2012, 6, 155–162. [Google Scholar] [CrossRef]
- Ross, E.A.; Abrahamson, D.R.; John, P.; Clapp, W.L.; Williams, M.J.; Terada, N.; Hamazaki, T.; Ellison, G.W.; Batich, C.D. Mouse stem cells seeded into decellularized rat kidney scaffolds endothelialize and remodel basement membranes. Organogenesis 2012, 8, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.H.; Houchin, T.L.; Shea, L.D. Gene delivery from polymer scaffolds for tissue engineering. Expert Rev. Med. Devices 2004, 1, 127–138. [Google Scholar] [CrossRef]
- Jang, J.H.; Rives, C.B.; Shea, L.D. Plasmid delivery in vivo from porous tissue-engineering scaffolds: Transgene expression and cellular transfection. Mol. Ther. 2005, 12, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Hsiong, S.X.; Mooney, D.J. Regeneration of vascularized bone. Periodontology 2000 2006, 41, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Rebar, E.J.; Huang, Y.; Hickey, R.; Nath, A.K.; Meoli, D.; Nath, S.; Chen, B.; Xu, L.; Liang, Y.; Jamieson, A.C.; et al. Induction of angiogenesis in a mouse model using engineered transcription factors. Nat. Med. 2002, 8, 1427–1432. [Google Scholar] [CrossRef]
- Debnath, J.; Brugge, J.S. Modelling glandular epithelial cancers in three-dimensional cultures. Nat. Rev. Cancer 2005, 5, 675–688. [Google Scholar] [CrossRef] [PubMed]
- Weaver, V.M.; Fischer, A.H.; Peterson, O.W.; Bissell, M.J. The importance of the microenvironment in breast cancer progression: Recapitulation of mammary tumorigenesis using a unique human mammary epithelial cell model and a three-dimensional culture assay. Biochem. Cell Biol. 1996, 74, 833–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Yi, S.; Ding, F.; Gong, L.; Gu, X. Extracellular matrix scaffolds for tissue engineering and regenerative medicine. Curr. Stem Cell Res. Ther. 2017, 12, 233–246. [Google Scholar] [CrossRef]
- Badylak, S.F.; Taylor, D.; Uygun, K. Whole-organ tissue engineering: Decellularization and recellularization of three-dimensional matrix scaffolds. Annu. Rev. Biomed. Eng. 2011, 13, 27. [Google Scholar] [CrossRef]
- Gilpin, A.; Yang, Y. Decellularization strategies for regenerative medicine: From processing techniques to applications. BioMed Res. Int. 2017, 2017, 9831534. [Google Scholar] [CrossRef] [Green Version]
- Babensee, J.E.; Anderson, J.M.; McIntire, L.V.; Mikos, A.G. Host response to tissue engineered devices. Adv. Drug Deliv. Rev. 1998, 33, 111–139. [Google Scholar] [CrossRef]
- Daly, K.A.; Liu, S.; Agrawal, V.; Brown, B.N.; Huber, A.; Johnson, S.A.; Reing, J.; Sicari, B.; Wolf, M.; Zhang, X.; et al. The host response to endotoxin-contaminated dermal matrix. Tissue Eng. 2012, 18, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Valentin, J.E.; Stewart-Akers, A.M.; Gilbert, T.W.; Badylak, S.F. Macrophage participation in the degradation and remodeling of extracellular matrix scaffolds. Tissue Eng. Part A 2009, 15, 1687. [Google Scholar] [CrossRef] [Green Version]
- Badylak, S.F.; Vorp, D.A.; Spievack, A.R.; Simmons-Byrd, A.; Hanke, J.; Freytes, D.O.; Thapa, A.; Gilbert, T.W.; Nieponice, A. Esophageal reconstruction with ECM and muscle tissue in a dog model. J. Surg. Res. 2005, 128, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Mobasseri, R.; Tian, L.; Soleimani, M.; Ramakrishna, S.; Naderi-Manesh, H. Peptide modified nanofibrous scaffold promotes human mesenchymal stem cell proliferation and long-term passaging. Mater. Sci. Eng. C Mater. Biol. Appl. 2018, 84, 80–89. [Google Scholar] [CrossRef]
- Hussein, K.H.; Saleh, T.; Ahmed, E.; Kwak, H.H.; Park, K.M.; Yang, S.R.; Kang, B.J.; Choi, K.Y.; Kang, K.S.; Woo, H.M. Biocompatibility and hemocompatibility of efficiently decellularized whole porcine kidney for tissue engineering. J. Biomed. Mater. Res. A. 2018, 106, 2034–2047. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Liu, S.; Liu, Y.; Sun, C.; Cheng, G.; Luan, Y.; Li, K.; Wang, J.; Xie, X.; Zhao, S. Porcine kidneys as a source of ECM scaffold for kidney regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 56, 451–456. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, N.; Hynds, R.E.; Gowers, K.; Tait, A.; Butler, C.R.; Hopper, C.; Burns, A.J.; Birchall, M.A.; Lowdell, M.; Janes, S.M. Using a Three-Dimensional Collagen Matrix to Deliver Respiratory Progenitor Cells to Decellularized Trachea In Vivo. Tissue Eng. Part C Methods 2019, 25, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Thakkar, S.; Fernandes, H.; Moroni, L. Decellularized Extracellular Matrix Scaffolds for Cartilage Regeneration. Methods Mol. Biol. 2015, 1340, 133–151. [Google Scholar] [PubMed]
- Zhao, Q.; Eberspaecher, H.; Lefebvre, V.; de Crombrugghe, B. Parallel expression of Sox9 and Col2a1 in cells undergoing chondrogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1997, 209, 377–386. [Google Scholar]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; De Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef]
- Canty, E.G.; Kadler, K.E. Procollagen trafficking, processing and fibrillogenesis. J. Cell Sci. 2005, 118, 1341–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kannu, P.; Bateman, J.; Savarirayan, R. Clinical phenotypes associated with type II collagen mutations. Paediatr. Child Health 2012, 48, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Cheah, K.S.; Stoker, N.G.; Griffin, J.R.; Grosveld, F.G.; Solomon, E. Identification and characterization of the human type II collagen gene (COL2A1). Proc. Natl. Acad. Sci. USA 1985, 82, 2555–2559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, B.R. Matrix molecules and their ligands. In Principles of Tissue Engineering; Academic Press: Cambridge, MA, USA, 2014; pp. 189–208. [Google Scholar]
- Glorieux, F.H. Pediatric Bone: Biology & Diseases; Elsevier: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Paradise, C.R. The Role of Bromodomain-Containing Protein 4 in Epigenetic Control of Skeletal Development. Doctoral Dissertation, College of Medicine-Mayo Clinic, Rochester, MN, USA, 2020. [Google Scholar]
- Le Goff, C.; Somerville, R.P.; Kesteloot, F.; Powell, K.; Birk, D.E.; Colige, A.C.; Apte, S.S. Regulation of Procollagen Amino-Propeptide Processing during Mouse Embryogenesis by Specialization of Homologous ADAMTS Proteases: Insights on Collagen Biosynthesis and Dermatosparaxis. Development 2006, 133, 1587–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAlinden, A.; Traeger, G.; Hansen, U.; Weis, M.A.; Ravindran, S.; Wirthlin, L.; Eyre, D.R.; Fernandes, R.J. Molecular properties and fibril ultrastructure of types II and XI collagens in cartilage of mice expressing exclusively the α1 (IIA) collagen isoform. Matrix Biol. 2014, 34, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Huang, J.H.; Sampson, E.R.; Kim, K.O.; Zuscik, M.J.; O’Keefe, R.J.; Chen, D.; Rosier, R.N. Smurf2 induces degradation of GSK-3β and upregulates β-catenin in chondrocytes: A potential mechanism for Smurf2-induced degeneration of articular cartilage. Exp. Cell Res. 2009, 315, 2386–2398. [Google Scholar] [CrossRef] [Green Version]
- Byers, P.H. Collagens: Building blocks at the end of the development line. Clin. Genet. 2000, 58, 270–279. [Google Scholar] [CrossRef]
- Desancé, M.; Contentin, R.; Bertoni, L.; Gomez-Leduc, T.; Branly, T.; Jacquet, S.; Betsch, J.M.; Batho, A.; Legendre, F.; Audigié, F.; et al. Chondrogenic differentiation of defined equine mesenchymal stem cells derived from umbilical cord blood for use in cartilage repair therapy. Int. J. Mol. Sci. 2018, 19, 537. [Google Scholar] [CrossRef] [Green Version]
- Neuhold, L.A.; Killar, L.; Zhao, W.; Sung, M.L.A.; Warner, L.; Kulik, J.; Turner, J.; Wu, W.; Billinghurst, C.; Meijers, T.; et al. Postnatal expression in hyaline cartilage of constitutively active human collagenase-3 (MMP-13) induces osteoarthritis in mice. J. Clin. Investig. 2001, 107, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Pelletier, J.P.; Boileau, C.; Brunet, J.; Boily, M.; Lajeunesse, D.; Reboul, P.; Laufer, S.; Martel-Pelletier, J. The inhibition of subchondral bone resorption in the early phase of experimental dog osteoarthritis by licofelone is associated with a reduction in the synthesis of MMP-13 and cathepsin K. Bone 2004, 34, 527–538. [Google Scholar] [CrossRef]
- Troeberg, L.; Nagase, H. Proteases involved in cartilage matrix degradation in osteoarthritis. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2012, 1824, 133–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, C.D.; Maity, S.N.; Lu, J.F.; Zhang, J.; Liang, S.; Coustry, F.; de Crombrugghe, B.; Yasuda, H. Identification of SOX9 interaction sites in the genome of chondrocytes. PLoS ONE 2010, 5, e10113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldwin, C.T.; Reginato, A.M.; Smith, C.; Jimenez, S.A.; Prockop, D.J. Structure of cDNA clones coding for human type II procollagen. The alpha 1(II) chain is more similar to the alpha 1(I) chain than two other alpha chains of fibrillar collagens. Prockop. Biochem. J. 1989, 262, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Kung, L.H.W.; Mullan, L.; Soul, J.; Wang, P.; Mori, K.; Bateman, J.F.; Briggs, M.D.; Boot-Handford, R.P. Cartilage endoplasmic reticulum stress may influence the onset but not the progression of experimental osteoarthritis. Arthritis Res. Ther. 2019, 21, 206. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.M.; Beier, F. Chondrocyte hypertrophy in skeletal development, growth, and disease. Birth Defects Res. C Embryo Today 2014, 102, 74–82. [Google Scholar] [CrossRef]
- Volz, M.; Schaumburger, J.; Frick, H.; Grifka, J.; Anders, S. A randomized controlled trial demonstrating sustained benefit of Autologous Matrix-Induced Chondrogenesis over microfracture at five years. Int. Orthop. 2017, 41, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-K.; Bae, H.C.; Ro, D.H.; Lee, S.; Lee, M.C.; Han, H.-S. Enhancement of Cartilage Regeneration of Synovial Stem Cells/Hydrogel by Using Transglutaminase-4. Tissue Eng. Part A 2021, 27, 761–770. [Google Scholar] [CrossRef]
- Dang, Q.T.; Huynh, T.D.; Inchingolo, F.; Dipalma, G.; Inchingolo, A.D.; Cantore, S.; Paduanelli, G.; Nguyen, K.C.D.; Ballini, A.; Isacco, C.G.; et al. Human Chondrocytes from Human Adipose Tissue-Derived Mesenchymal Stem Cells Seeded on a Dermal-Derived Collagen Matrix Sheet: Our Preliminary Results for a Ready to Go Biotechnological Cartilage Graft in Clinical Practice. Stem Cells Int. 2021, 2021, 6664697. [Google Scholar] [CrossRef]
- Li, D.; Zhang, L.; Liang, J.; Deng, W.; Wei, Q.; Wang, K. Biofilm Formation by Pseudomonas aeruginosa in a Novel Septic Arthritis Model. Front. Cell. Infect. Microbiol. 2021, 11, 884. [Google Scholar] [CrossRef]
- Castro, R.D.J.C.; Sobrinho, F.S.; da Gama, M.A.S.; de Castro Barra, P.M.; Antoniassi, R.; de Oliveira, M.A. Analysis of the main conjugated linoleic acid (CLA) precursors (C18: 2 n-6 and C18: 3 n-3) in Brachiaria ruzizienses by capillary zone electrophoresis. Cent. Eur. J. Chem. 2013, 11, 1286–1296. [Google Scholar]
- Le Vien, N.T.; Nguyen, P.B.; Cuong, L.D.; An, T.T.T.; Dao, D.T.A. Optimization of papain hydrolysis conditions for release of glycosaminoglycans from the chicken keel cartilage. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2017; Volume 1878, p. 020009. [Google Scholar]
- Burkhardt, D.; Hwa, S.Y.; Ghosh, P. A novel microassay for the quantitation of the sulfated glycosaminoglycan content of histological sections: Its application to determine the effects of Diacerhein on cartilage in an ovine model of osteoarthritis. Osteoarthr. Cartil. 2001, 9, 238–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’brien, J.; Wilson, I.; Orton, T.; Pognan, F. Investigation of the Alamar Blue (resazurin) fluorescent dye for the assessment of mammalian cell cytotoxicity. Eur. J. Biochem. 2000, 267, 5421–5426. [Google Scholar] [CrossRef] [PubMed]
- Rampersad, S.N. Multiple applications of Alamar Blue as an indicator of metabolic function and cellular health in cell viability bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef]
- Schmitt, A.; Csiki, R.; Tron, A.; Saldamli, B.; Tübel, J.; Florian, K.; Siebenlist, S.; Balmayor, E.; Burgkart, R. Optimized protocol for whole organ decellularization. Eur. J. Med. Res. 2017, 22, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, E.R.; Parry, J.; Dadsetan, M.; Bravo, D.; Riester, S.M.; van Wijnen, A.J.; Yaszemski, M.J.; Kakar, S. Chondrocyte attachment, proliferation, and differentiation on three-dimensional polycaprolactone fumarate scaffolds. Tissue Eng. Part A 2017, 23, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.S.; Zhou, Z.L.; Luo, J.W.; Zhang, H.; Hou, J.F. The Ihh signal is essential for regulating proliferation and hypertrophy of cultured chicken chondrocytes. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2013, 166, 117–122. [Google Scholar] [CrossRef]
- Ayariga, J.A.; Dean, M.; Nyairo, E.; Thomas, V.; Dean, D. PLA/HA Multiscale Nano-/Micro-Hybrid 3D Scaffolds Provide Inductive Cues to Stems Cells to Differentiate into an Osteogenic Lineage. JOM 2021, 73, 3787–3797. [Google Scholar] [CrossRef]
- Huang, H.; Ayariga, J.; Ning, H.; Nyairo, E.; Dean, D. Freeze-printing of pectin/alginate scaffolds with high resolution, overhang structures and interconnected porous network. Addit. Manuf. 2021, 46, 102120. [Google Scholar] [CrossRef]
- Baiocchini, A.; Montaldo, C.; Conigliaro, A.; Grimaldi, A.; Correani, V.; Mura, F.; Ciccosanti, F.; Rotiroti, N.; Brenna, A.; Montalbano, M.; et al. Extracellular matrix molecular remodeling in human liver fibrosis evolution. PLoS ONE 2016, 11, e0151736. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, A.; Nakajima, A.; Nakajima, F.; Goto, K.I.; Yamazaki, M. Molecular basis for affected cartilage formation and bone union in fracture healing of the streptozotocin-induced diabetic rat. Bone 2008, 43, 832–839. [Google Scholar] [CrossRef]
- Hoppe, A.; Boccaccini, A.R. 7—Bioactive glass foams for tissue engineering applications. Biomed. Foam. Tissue Eng. Appl. 2014, 191–212. [Google Scholar] [CrossRef]
- Belviso, I.; Romano, V.; Sacco, A.M.; Ricci, G.; Massai, D.; Cammarota, M.; Catizone, A.; Nurzynska, S.D.; Terzini, M.; Aldieri, A.; et al. Decellularized Human Dermal Matrix as a Biological Scaffold for Cardiac Repair and Regeneration. Front. Bioeng. Biotechnol. 2020, 20, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Input Template ID |
|---|---|---|
| FW COL2A1 | 5′-GTAGAGACCCGGACCCGC-3′ | NM_001844.5 |
| RV COL2A1 | 5′-ACTCTCCGAAGGGGATCTCA-3′ | NM_001844.5 |
| FW COL5A1 | 5′-TTCAAGCGTGGGAAACTGCT-3′ | NM_000093.5 |
| RV COL5A1 | 5′-GGGAGAAGCCTTCACTGTCC-3′ | NM_000093.5 |
| FW COL4A2 | 5′-ATAGGAGGGCCCAAGGGATT-3′ | NM_001846.4 |
| RV COL4A2 | 5′-CAGGGTCCCCTCTATCACCA-3′ | NM_001846.4 |
| FW COL6A1 | 5′-ATTGCCAAGGACTTCGTCGT-3′ | NM_001848.3 |
| RV COL6A1 | 5′-ACATTGAGCTGGTCTGAGCC-3′ | NM_001848.3 |
| FW GAPDH | 5′-CCATGGGGAAGGTGAAGGTC-3′ | NM_002046.7 |
| RV GAPDH | 5′-AGTGATGGCATGGACTGTGG-3′ | NM_002046.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayariga, J.A.; Huang, H.; Dean, D. Decellularized Avian Cartilage, a Promising Alternative for Human Cartilage Tissue Regeneration. Materials 2022, 15, 1974. https://doi.org/10.3390/ma15051974
Ayariga JA, Huang H, Dean D. Decellularized Avian Cartilage, a Promising Alternative for Human Cartilage Tissue Regeneration. Materials. 2022; 15(5):1974. https://doi.org/10.3390/ma15051974
Chicago/Turabian StyleAyariga, Joseph Atia, Hanxiao Huang, and Derrick Dean. 2022. "Decellularized Avian Cartilage, a Promising Alternative for Human Cartilage Tissue Regeneration" Materials 15, no. 5: 1974. https://doi.org/10.3390/ma15051974
APA StyleAyariga, J. A., Huang, H., & Dean, D. (2022). Decellularized Avian Cartilage, a Promising Alternative for Human Cartilage Tissue Regeneration. Materials, 15(5), 1974. https://doi.org/10.3390/ma15051974