

Serial Cultivation of an MSC-Like Cell Line with Enzyme-Free Passaging Using a Microporous Titanium Scaffold

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
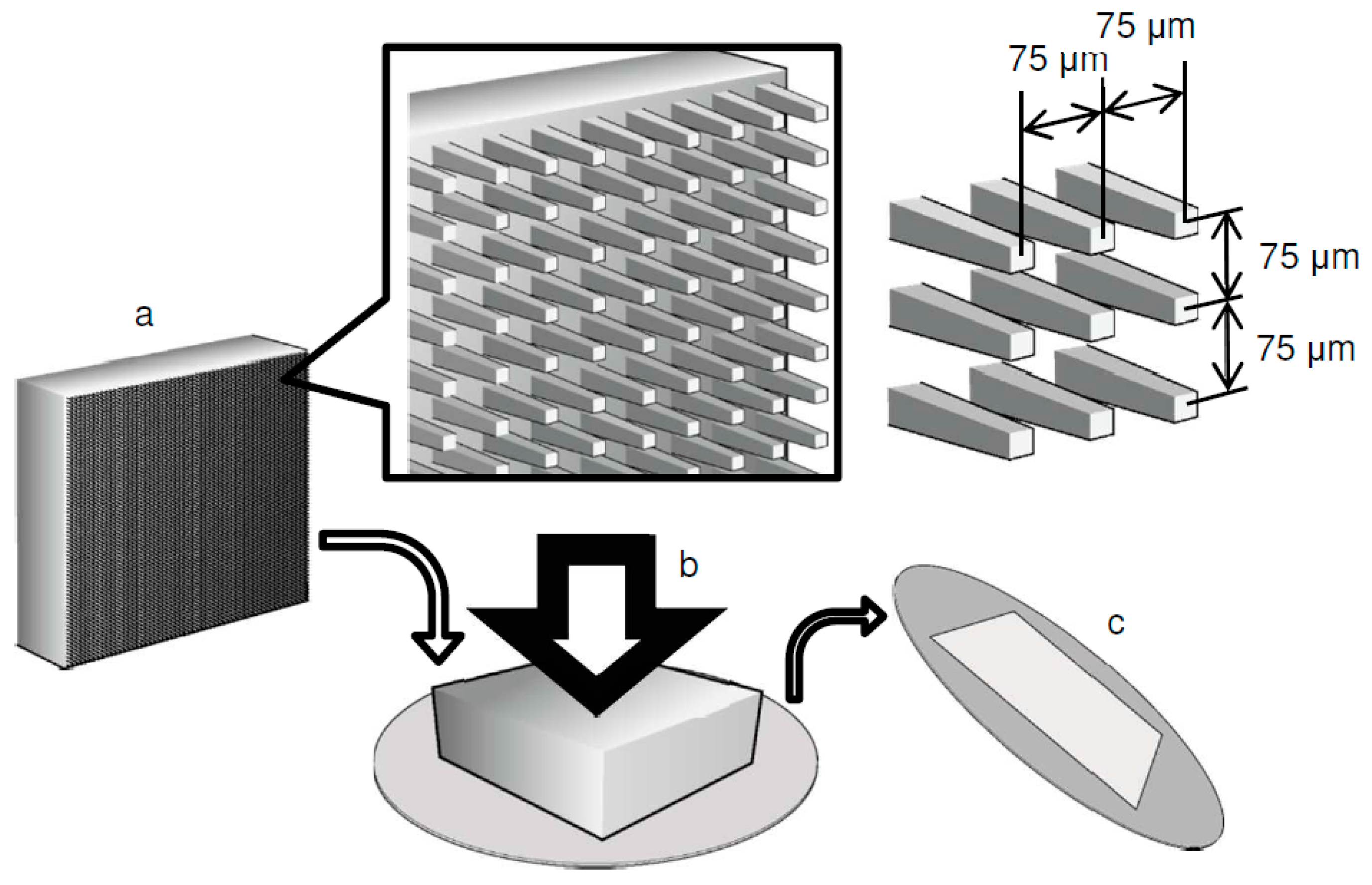
2.1. Preparation of a Microarrayed Pin Needle Punch (MPNP) and Micropiercing
2.2. Cell Culture
2.3. Scanning Electron Microscopy (SEM) Observations of the Cell Morphology
2.4. Immunofluorescence Assay
2.5. Statistical Analyses
3. Results
3.1. Fabrication of a Microperforated Ti Membrane
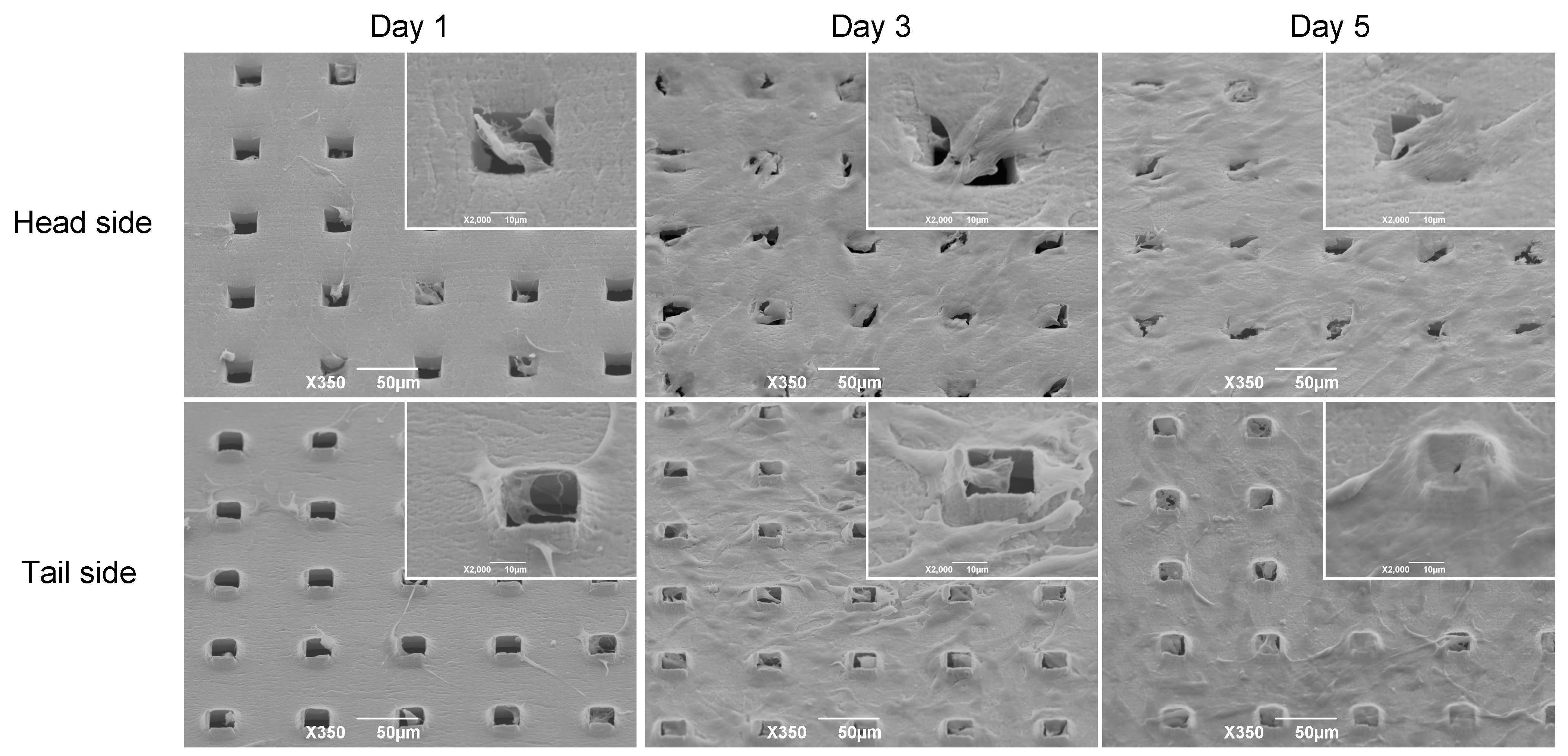
3.2. Morphology of C3H10T1/2 Cells on the Microperforated Ti Membrane
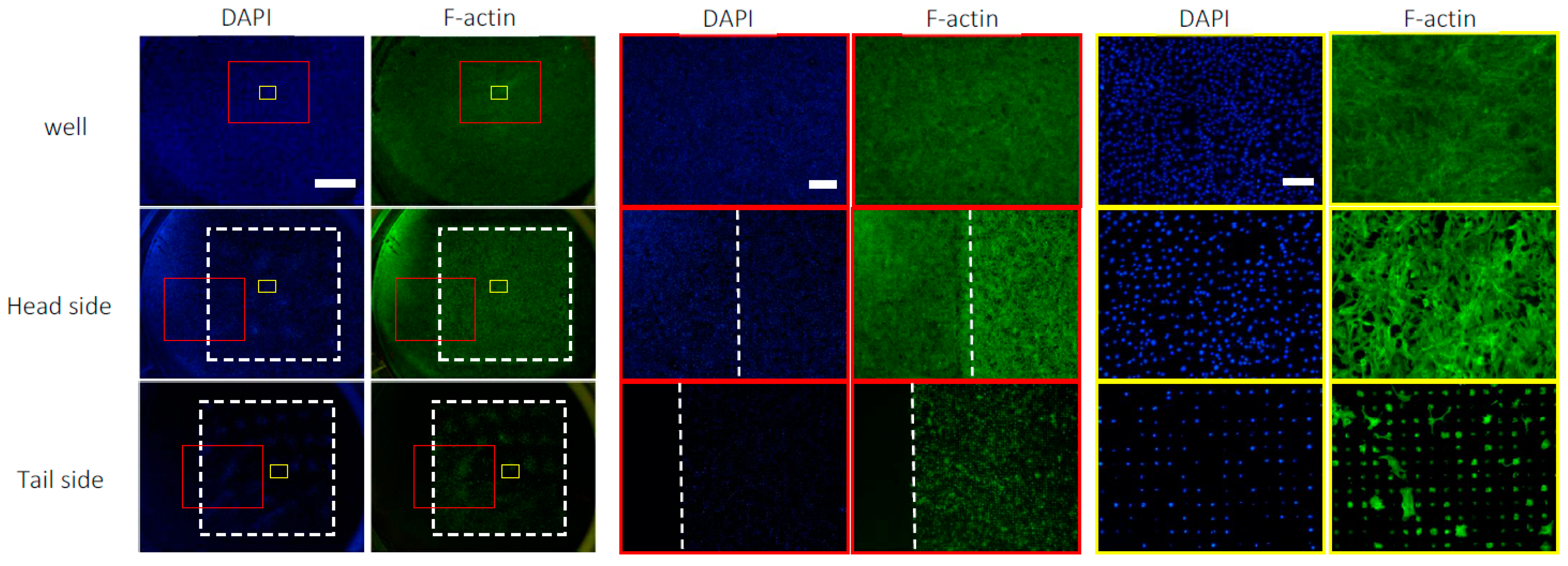
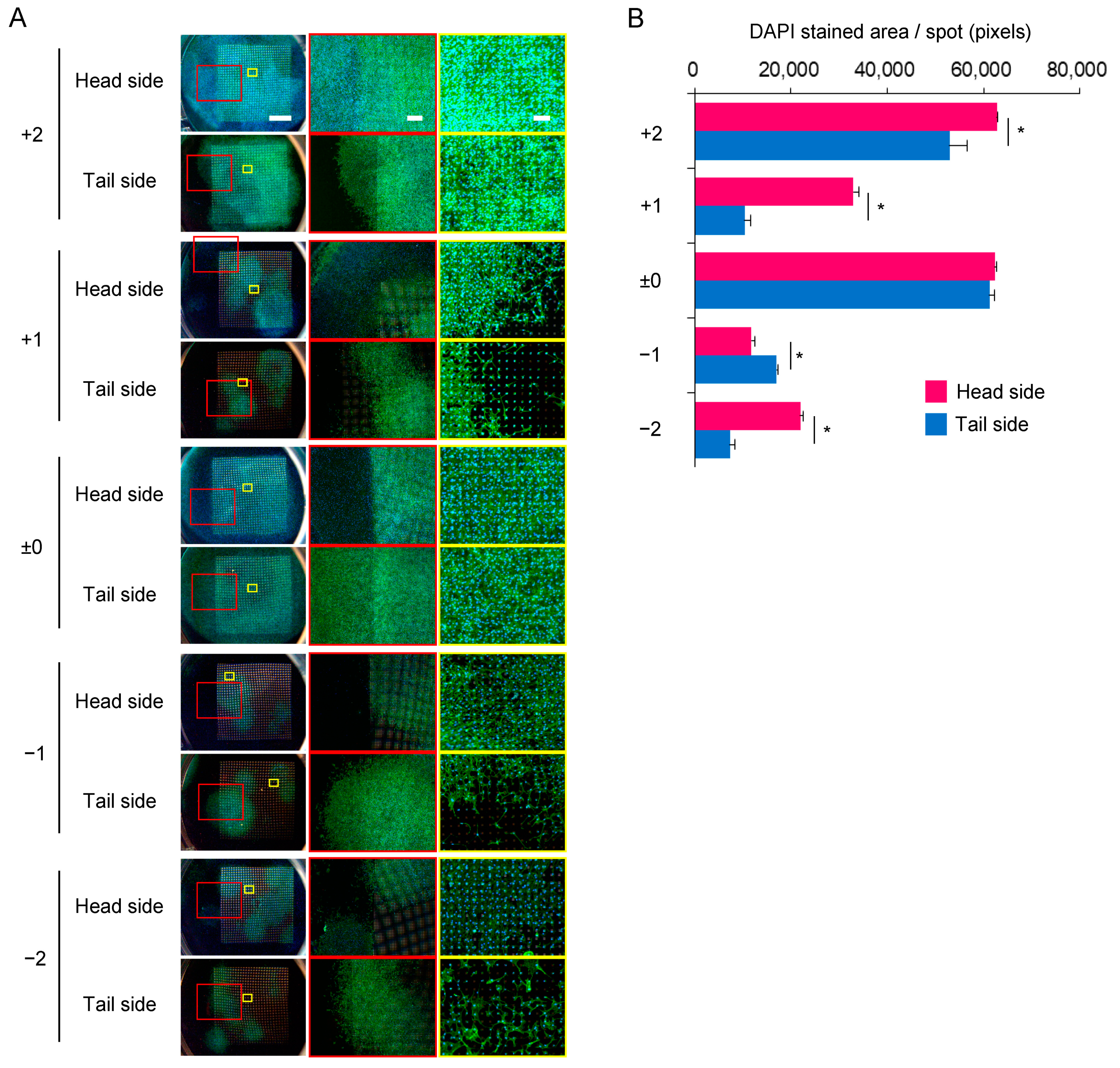
3.3. Distribution of C3H10T1/2 Cells on the Microperforated Ti Membrane
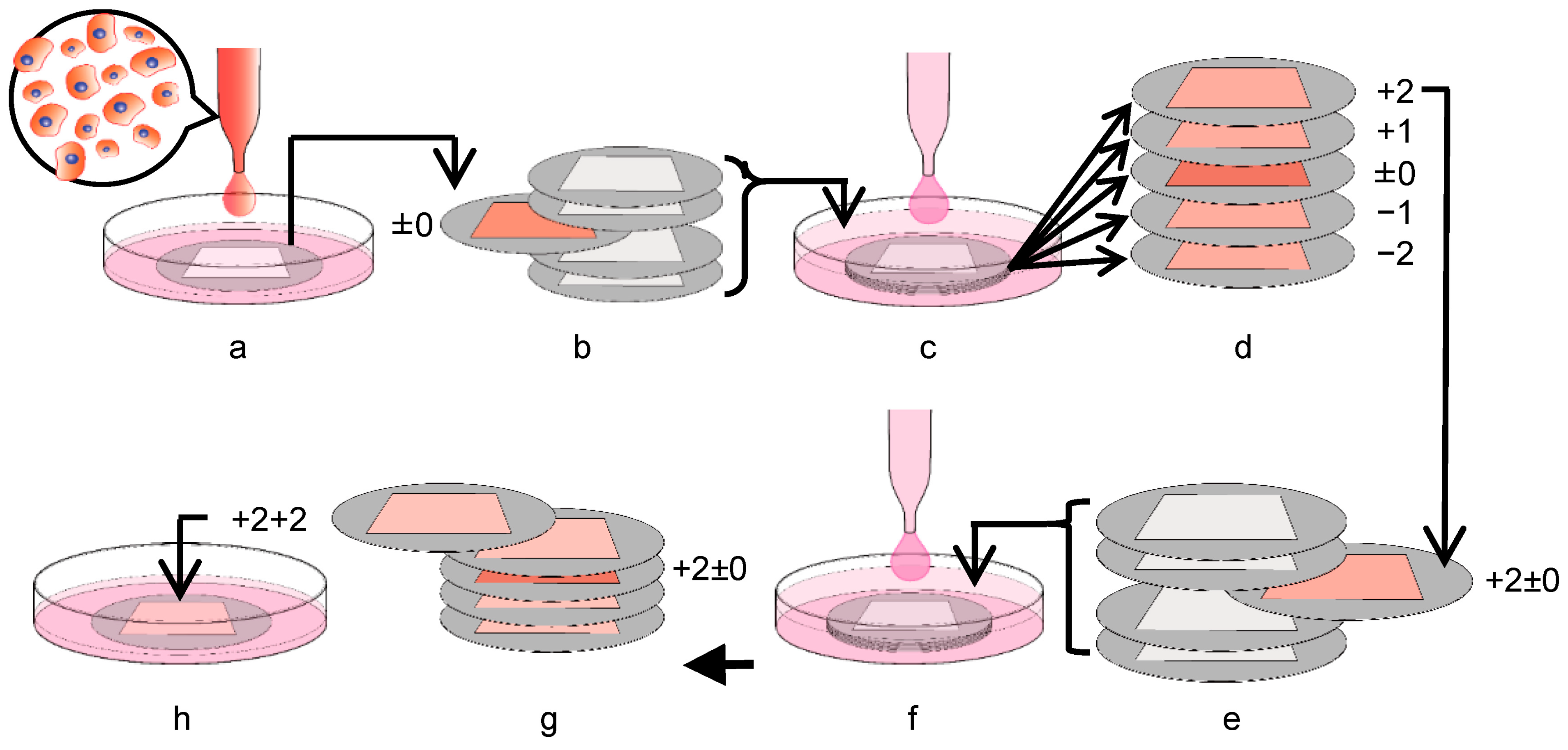
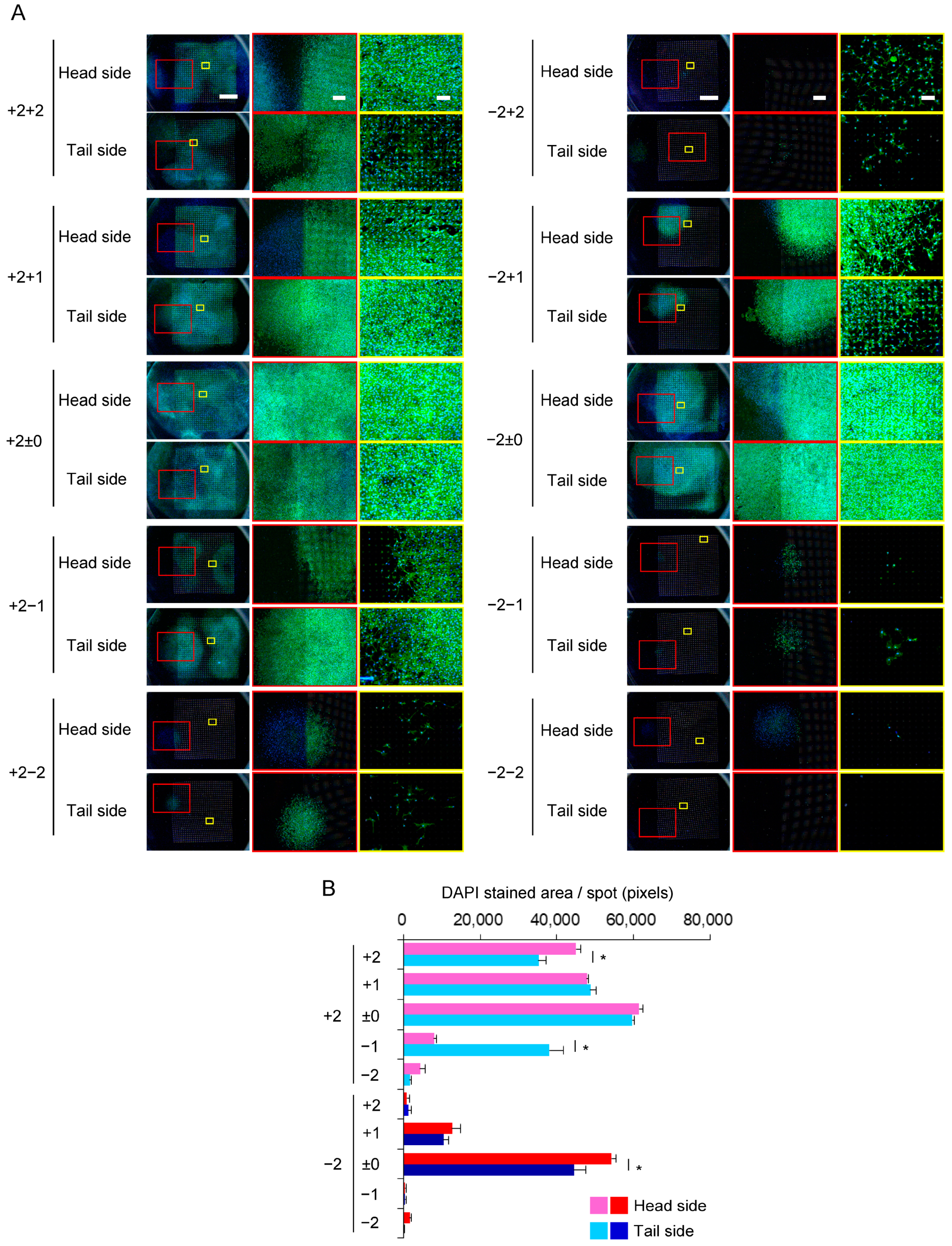
3.4. Expansion of C3H10T1/2 Cells in the Multi-Layered Culture
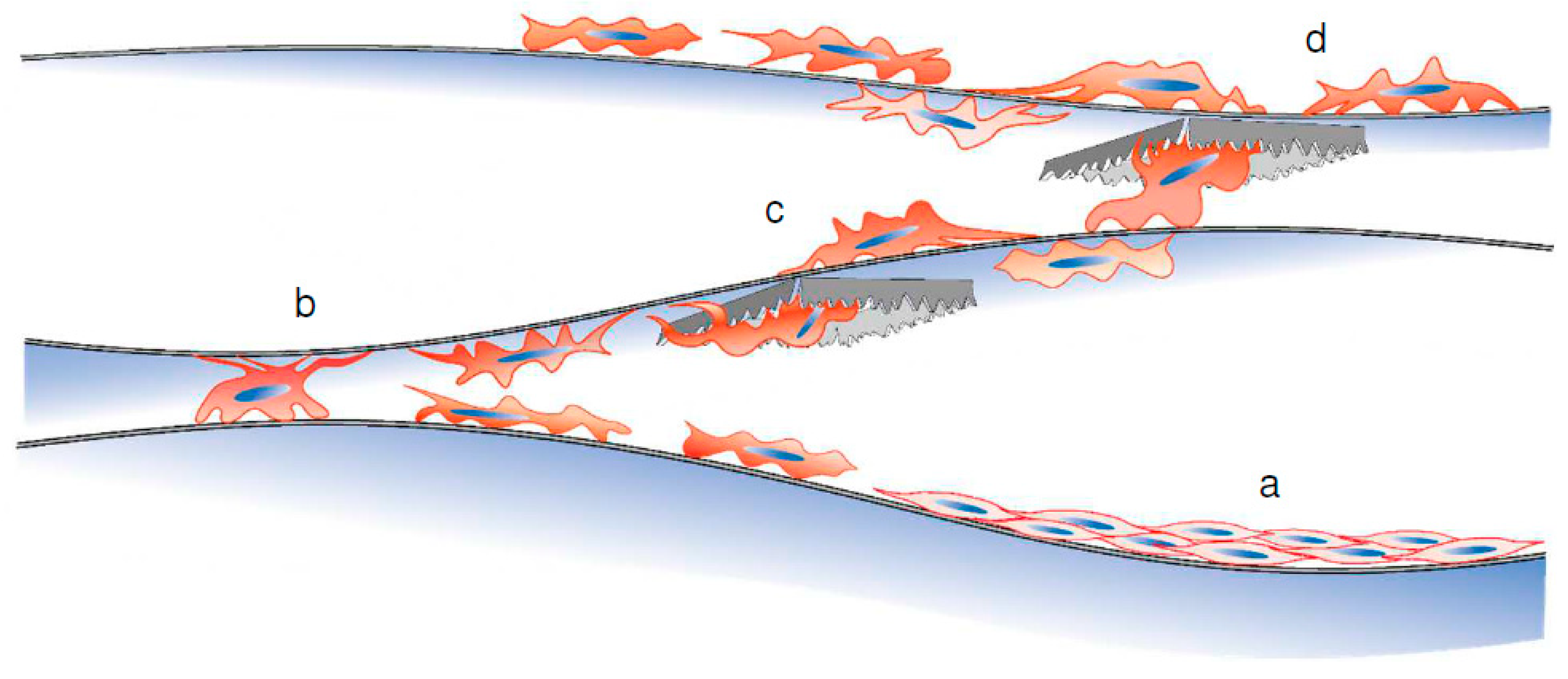
3.5. Transfer of C3H10T1/2 Cells from the Ti Membrane to a Conventional Culture Plate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lechanteur, C.; Briquet, A.; Giet, O.; Delloye, O.; Baudoux, E.; Beguin, Y. Clinical-scale expansion of mesenchymal stromal cells: A large banking experience. J. Transl. Med. 2016, 14, 145. [Google Scholar] [CrossRef] [Green Version]
- Resnick, I.B.; Barkats, C.; Shapira, M.Y.; Stepensky, P.; Bloom, A.I.; Shimoni, A.; Mankuta, D.; Varda-Bloom, N.; Rheingold, L.; Yeshurun, M.; et al. Treatment of severe steroid resistant acute GVHD with mesenchymal stromal cells (MSC). Am. J. Blood. Res. 2013, 3, 225–238. [Google Scholar]
- Heng, B.C.; Cowan, C.M.; Basu, S. Comparison of enzymatic and nonenzymatic means of dissociating adherent monolayers of mesenchymal stem cells. Biol. Proced. Online 2009, 11, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Pervin, B.; Aydın, G.; Visser, T.; Uçkan-Çetinkaya, D.; Aerts-Kaya, F.S. CXCR4 expression by mesenchymal stromal cells is lost after use of enzymatic dissociation agents, but preserved by use of non-enzymatic methods. Int. J. Hematol. 2021, 113, 5–9. [Google Scholar] [CrossRef]
- Pilbauerová, N.; Soukup, T.; Kleplová, T.S.; Suchánek, J. Enzymatic Isolation, Amplification and Characterization of Dental Pulp Stem Cells. Folia Biol. 2019, 65, 124–133. [Google Scholar]
- Garitaonandia, I.; Amir, H.; Boscolo, F.S.; Wambua, G.K.; Schultheisz, H.L.; Sabatini, K.; Morey, R.; Waltz, S.; Wang, Y.C.; Tran, H.; et al. Increased risk of genetic and epigenetic instability in human embryonic stem cells associated with specific culture conditions. PLoS ONE 2015, 10, e0118307. [Google Scholar] [CrossRef]
- Watanabe, K.; Ueno, M.; Kamiya, D.; Nishiyama, A.; Matsumura, M.; Wataya, T.; Takahashi, J.B.; Nishikawa, S.; Nishikawa, S.; Muguruma, K.; et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nat. Biotechnol. 2007, 25, 681–686. [Google Scholar] [CrossRef]
- Badur, M.G.; Zhang, H.; Metallo, C.M. Enzymatic passaging of human embryonic stem cells alters central carbon metabolism and glycan abundance. Biotechnol. J. 2015, 10, 1600–1611. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Walsh, P.; Clarke, D.L.; Rowley, J.A.; Fellner, T. Scalable passaging of adherent human pluripotent stem cells. PLoS ONE 2014, 9, e88012. [Google Scholar] [CrossRef] [Green Version]
- Ghorbani, A.; Jalali, S.A.; Varedi, M. Isolation of adipose tissue mesenchymal stem cells without tissue destruction: A non-enzymatic method. Tissue Cell 2014, 46, 54–58. [Google Scholar] [CrossRef]
- Kuriyama, T.; Fukuma, Y.; Imashiro, C.; Kabayama, K.; Kurashina, Y.; Takemura, K. Detachment of RAW264.7 macrophages from a culture dish using ultrasound excited by a Langevin transducer. J. Biosci. Bioeng. 2021, 131, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Batista, U.; Garvas, M.; Nemec, M.; Schara, M.; Veranič, P.; Koklic, T. Effects of different detachment procedures on viability, nitroxide reduction kinetics and plasma membrane heterogeneity of V-79 cells. Cell Biol. Int. 2010, 34, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Yan, Y.; Wang, X.; Fu, Q.; Wang, W.; Wu, J.; Yang, G.; Ren, J.; Wang, Y. Impact of cell dissociation on identification of breast cancer stem cells. Cancer Biomark 2012, 12, 125–133. [Google Scholar] [CrossRef]
- Nakashima, Y.; Akaike, M.; Kounoura, M.; Hayashi, K.; Morita, K.; Oki, Y.; Nakanishi, Y. Evaluation of osteoblastic cell behavior upon culture on titanium substrates photo-functionalized by vacuum ultra-violet treatment. Exp. Cell Res. 2022, 410, 112944. [Google Scholar] [CrossRef] [PubMed]
- Gardin, C.; Ferroni, L.; Erdoğan, Y.K.; Zanotti, F.; De Francesco, F.; Trentini, M.; Brunello, G.; Ercan, B.; Zavan, B. Nanostructured modifications of titanium surfaces improve vascular regenerative properties of exosomes derived from mesenchymal stem cells: Preliminary in vitro results. Nanomaterials 2021, 11, 3452. [Google Scholar] [CrossRef]
- Zhang, J.; Sakisaka, Y.; Ishihata, H.; Maruyama, K.; Nemoto, E.; Chiba, S.; Nagamine, M.; Hasegawa, H.; Yamada, S. Evaluation of preosteoblast MC3T3-E1 cells cultured on a microporous titanium membrane fabricated using a precise mechanical punching process. Materials 2020, 13, 5288. [Google Scholar] [CrossRef]
- Hasegawa, H.; Masui, S.; Ishihata, H. New microperforated pure titanium membrane created by laser processing for guided regeneration of bone. Br. J. Oral Maxillofac. Surg. 2018, 56, 642–643. [Google Scholar] [CrossRef]
- Ishihara, K.; Kaneyasu, M.; Fukazawa, K.; Zhang, R.; Teramura, Y. Induction of mesenchymal stem cell differentiation by co-culturing with mature cells in double-layered 2-methacryloyloxyethyl phosphorylcholine polymer hydrogel matrices. J. Mater. Chem. B 2022, 10, 2561–2569. [Google Scholar] [CrossRef]
- Hanawa, T. Biocompatibility of titanium from the viewpoint of its surface. Sci. Technol. Adv. Mater. 2022, 23, 457–472. [Google Scholar] [CrossRef]
- Cohen, D.J.; Cheng, A.; Sahingur, K.; Clohessy, R.M.; Hopkins, L.B.; Boyan, B.D.; Schwartz, Z. Performance of laser sintered Ti-6Al-4V implants with bone-inspired porosity and micro/nanoscale surface roughness in the rabbit femur. Biomed. Mater. 2017, 12, 025021. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, P.; Jin, J.; Li, L.; He, S.Y.; Zhou, P.; Jiang, Q.; Wen, C. In silico and in vivo studies of the effect of surface curvature on the osteoconduction of porous scaffolds. Biotechnol. Bioeng. 2022, 119, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Yilgor, P.; Yilmaz, G.; Onal, M.B.; Solmaz, I.; Gundogdu, S.; Keskil, S.; Sousa, R.A.; Reis, R.L.; Hasirci, N.; Hasirci, V. An in vivo study on the effect of scaffold geometry and growth factor release on the healing of bone defects. J. Tissue Eng. Regen. Med. 2013, 7, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, E.W.; Wang, Y.B.; Shuai, K.G.; Gao, F.; Bai, Y.J.; Cheng, Y.; Xiong, X.L.; Zheng, Y.F.; Wei, S.C. In vitro and in vivo evaluation of SLA titanium surfaces with further alkali or hydrogen peroxide and heat treatment. Biomed. Mater. 2011, 6, 025001. [Google Scholar] [CrossRef] [Green Version]
- Gil-Redondo, J.C.; Weber, A.; Zbiral, B.; Vivanco, M.D.; Toca-Herrera, J.L. Substrate stiffness modulates the viscoelastic properties of MCF-7 cells. J. Mech. Behav. Biomed. Mater. 2022, 125, 104979. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Doss, B.; Lim, C.T.; Voituriez, R.; Ladoux, B. Single cell rigidity sensing: A complex relationship between focal adhesion dynamics and large-scale actin cytoskeleton remodeling. Cell Adh. Migr. 2016, 10, 554–567. [Google Scholar] [CrossRef] [Green Version]
- Hopp, I.; Michelmore, A.; Smith, L.E.; Robinson, D.E.; Bachhuka, A.; Mierczynska, A.; Vasilev, K. The influence of substrate stiffness gradients on primary human dermal fibroblasts. Biomaterials 2013, 34, 5070–5077. [Google Scholar] [CrossRef]
- Lang, N.P. Oral Implants: The Paradigm Shift in Restorative Dentistry. J. Dent. Res. 2019, 98, 1287–1293. [Google Scholar] [CrossRef]
- Gao, H.; Li, X.; Wang, C.; Ji, P.; Wang, C. Mechanobiologically optimization of a 3D titanium-mesh implant for mandibular large defect: A simulated study. Mater. Sci. Eng. C 2019, 104, 109934. [Google Scholar] [CrossRef]
- Ilea, A.; Vrabie, O.G.; Băbțan, A.M.; Miclăuş, V.; Ruxanda, F.; Sárközi, M.; Barbu-Tudoran, L.; Mager, V.; Berce, C.; Boşca, A.B.; et al. Osseointegration of titanium scaffolds manufactured by selective laser melting in rabbit femur defect model. J. Mater. Sci. Mater. Med. 2019, 30, 26. [Google Scholar] [CrossRef]
- Fukuhira, Y.; Kitazono, E.; Hayashi, T.; Kaneko, H.; Tanaka, M.; Shimomura, M.; Sumia, Y. Biodegradable honeycomb-patterned film composed of poly(lactic acid) and dioleoylphosphatidylethanolamine. Biomaterials 2006, 27, 1797–1802. [Google Scholar] [CrossRef]
- Yamazaki, H.; Gotou, S.; Ito, K.; Kohashi, S.; Goto, Y.; Yoshiura, Y.; Sakai, Y.; Yabu, H.; Shimomura, M.; Nakazawa, K. Micropatterned culture of HepG2 spheroids using microwell chip with honeycomb-patterned polymer film. J. Biosci. Bioeng. 2014, 118, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Langhans, S.A. Three-dimensional in vitro cell culture models in drug discovery and drug repositioning. Front. Pharmacol. 2018, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Campos Marín, A.; Brunelli, M.; Lacroix, D. Flow perfusion rate modulates cell deposition onto scaffold substrate during cell seeding. Biomech. Model. Mechanobiol. 2018, 17, 675–687. [Google Scholar] [CrossRef] [Green Version]
- De Peppo, G.M.; Marcos-Campos, I.; Kahler, D.J.; Alsalman, D.; Shang, L.; Vunjak-Novakovic, G.; Marolt, D. Engineering bone tissue substitutes from human induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, 8680–8685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porta, G.D.; Nguyen, B.N.B.; Campardelli, R.; Reverchon, E.; Fisher, J.P. Synergistic effect of sustained release of growth factors and dynamic culture on osteoblastic differentiation of mesenchymal stem cells. J. Biomed. Mater. Res. A 2015, 103, 2161–2171. [Google Scholar] [CrossRef]
- Shi, F.; Xiao, D.; Zhang, C.; Zhi, W.; Liu, Y.; Weng, J. The effect of macropore size of hydroxyapatite scaffold on the osteogenic differentiation of bone mesenchymal stem cells under perfusion culture. Regen. Biomater. 2021, 8, rbab050. [Google Scholar] [CrossRef]
- Fontoura, J.C.; Viezzer, C.; Dos Santos, F.G.; Ligabue, R.A.; Weinlich, R.; Puga, R.D.; Antonow, D.; Severino, P.; Bonorino, C. Comparison of 2D and 3D cell culture models for cell growth, gene expression and drug resistance. Mater. Sci. Eng. C 2020, 107, 110264. [Google Scholar] [CrossRef]
- Raghav, P.K.; Mann, Z.; Ahlawat, S.; Mohanty, S. Mesenchymal stem cell-based nanoparticles and scaffolds in regenerative medicine. Eur. J. Pharmacol. 2022, 918, 174657. [Google Scholar] [CrossRef]
- Leber, J.; Barekzai, J.; Blumenstock, M.; Pospisil, B.; Salzig, D.; Czermak, P. Microcarrier choice and bead-to-bead transfer for human mesenchymal stem cells in serum-containing and chemically defined media. Process Biochem. 2017, 59, 255–265. [Google Scholar] [CrossRef]
- Ranjan, V.D.; Zeng, P.; Li, B.; Zhang, Y. In vitro cell culture in hollow microfibers with porous structures. Biomater. Sci. 2020, 8, 2175–2188. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakisaka, Y.; Ishihata, H.; Maruyama, K.; Nemoto, E.; Chiba, S.; Nagamine, M.; Hasegawa, H.; Hatsuzawa, T.; Yamada, S. Serial Cultivation of an MSC-Like Cell Line with Enzyme-Free Passaging Using a Microporous Titanium Scaffold. Materials 2023, 16, 1165. https://doi.org/10.3390/ma16031165
Sakisaka Y, Ishihata H, Maruyama K, Nemoto E, Chiba S, Nagamine M, Hasegawa H, Hatsuzawa T, Yamada S. Serial Cultivation of an MSC-Like Cell Line with Enzyme-Free Passaging Using a Microporous Titanium Scaffold. Materials. 2023; 16(3):1165. https://doi.org/10.3390/ma16031165
Chicago/Turabian StyleSakisaka, Yukihiko, Hiroshi Ishihata, Kentaro Maruyama, Eiji Nemoto, Shigeki Chiba, Masaru Nagamine, Hiroshi Hasegawa, Takeshi Hatsuzawa, and Satoru Yamada. 2023. "Serial Cultivation of an MSC-Like Cell Line with Enzyme-Free Passaging Using a Microporous Titanium Scaffold" Materials 16, no. 3: 1165. https://doi.org/10.3390/ma16031165
APA StyleSakisaka, Y., Ishihata, H., Maruyama, K., Nemoto, E., Chiba, S., Nagamine, M., Hasegawa, H., Hatsuzawa, T., & Yamada, S. (2023). Serial Cultivation of an MSC-Like Cell Line with Enzyme-Free Passaging Using a Microporous Titanium Scaffold. Materials, 16(3), 1165. https://doi.org/10.3390/ma16031165