
Influence of Chlorination and Choice of Materials on Fouling in Cooling Water System under Brackish Seawater Conditions

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
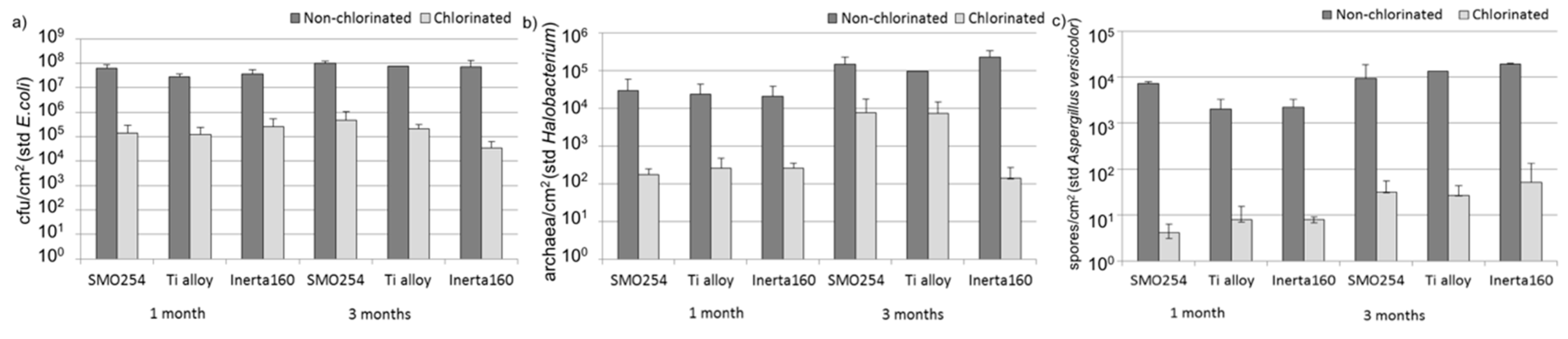
2.1. Biofouling
2.1.1. Quantification of Micro-Organisms
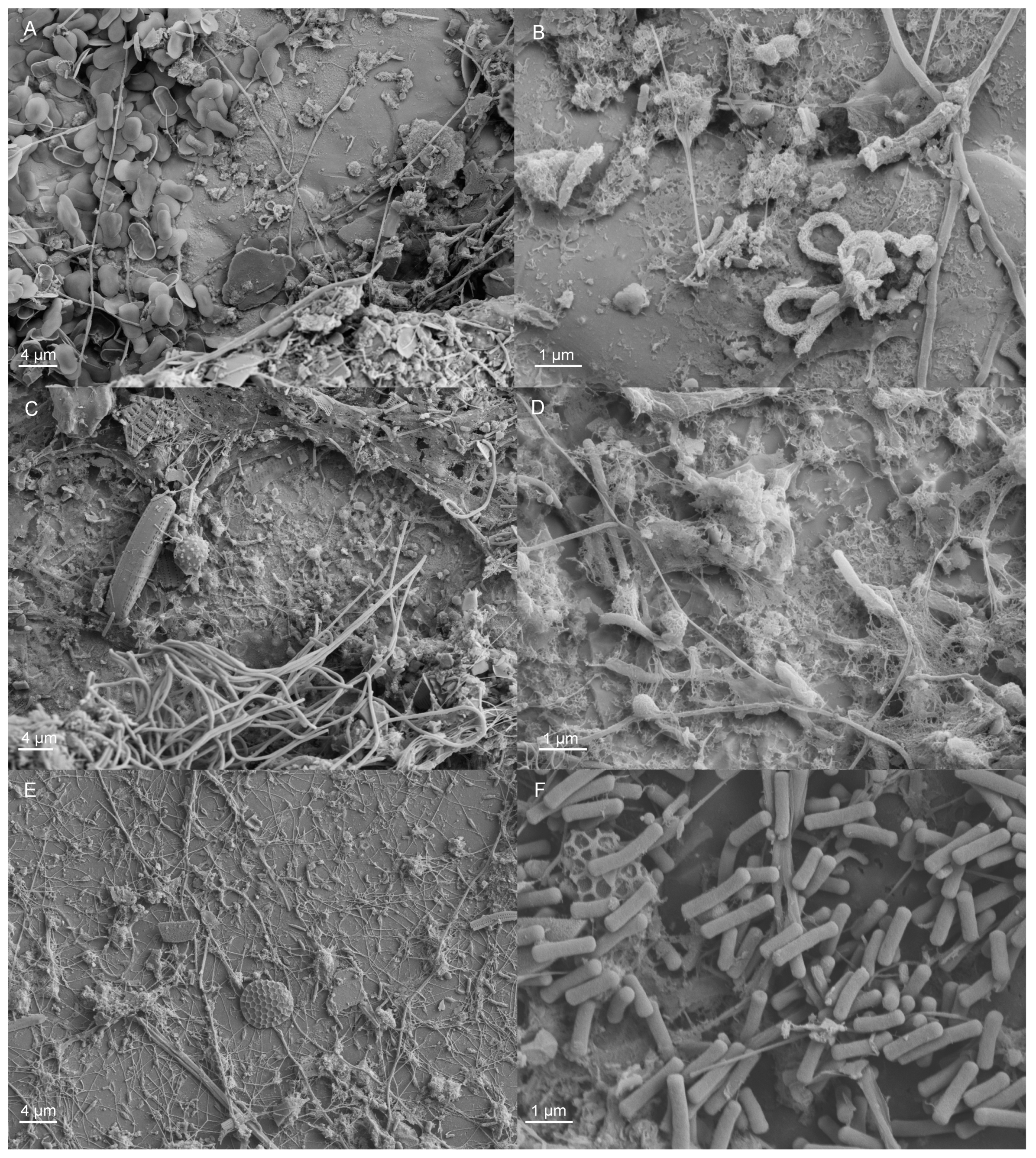
2.1.2. Microscopy
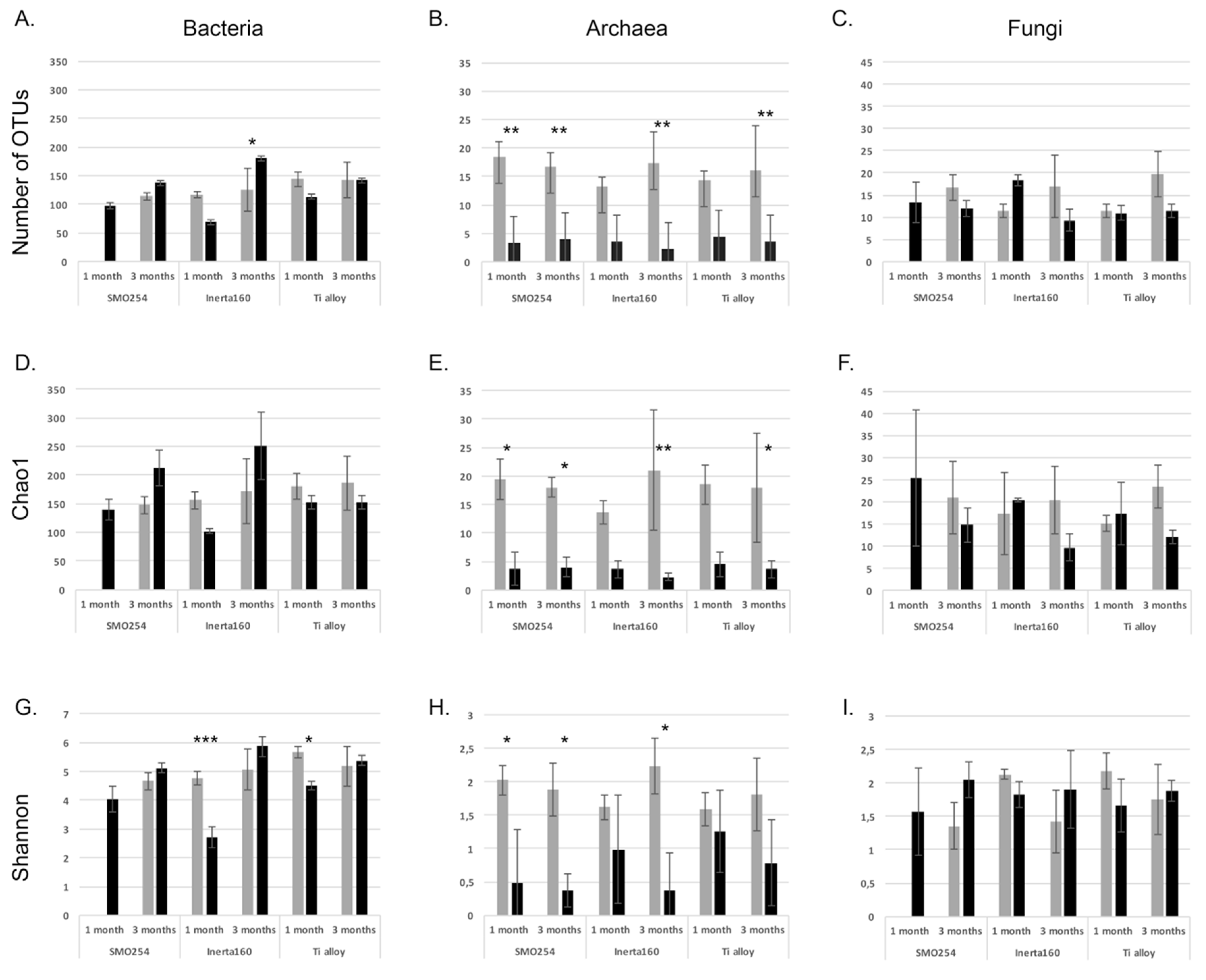
2.1.3. Effect of Chlorination on the Richness and Diversity of the Biofilm
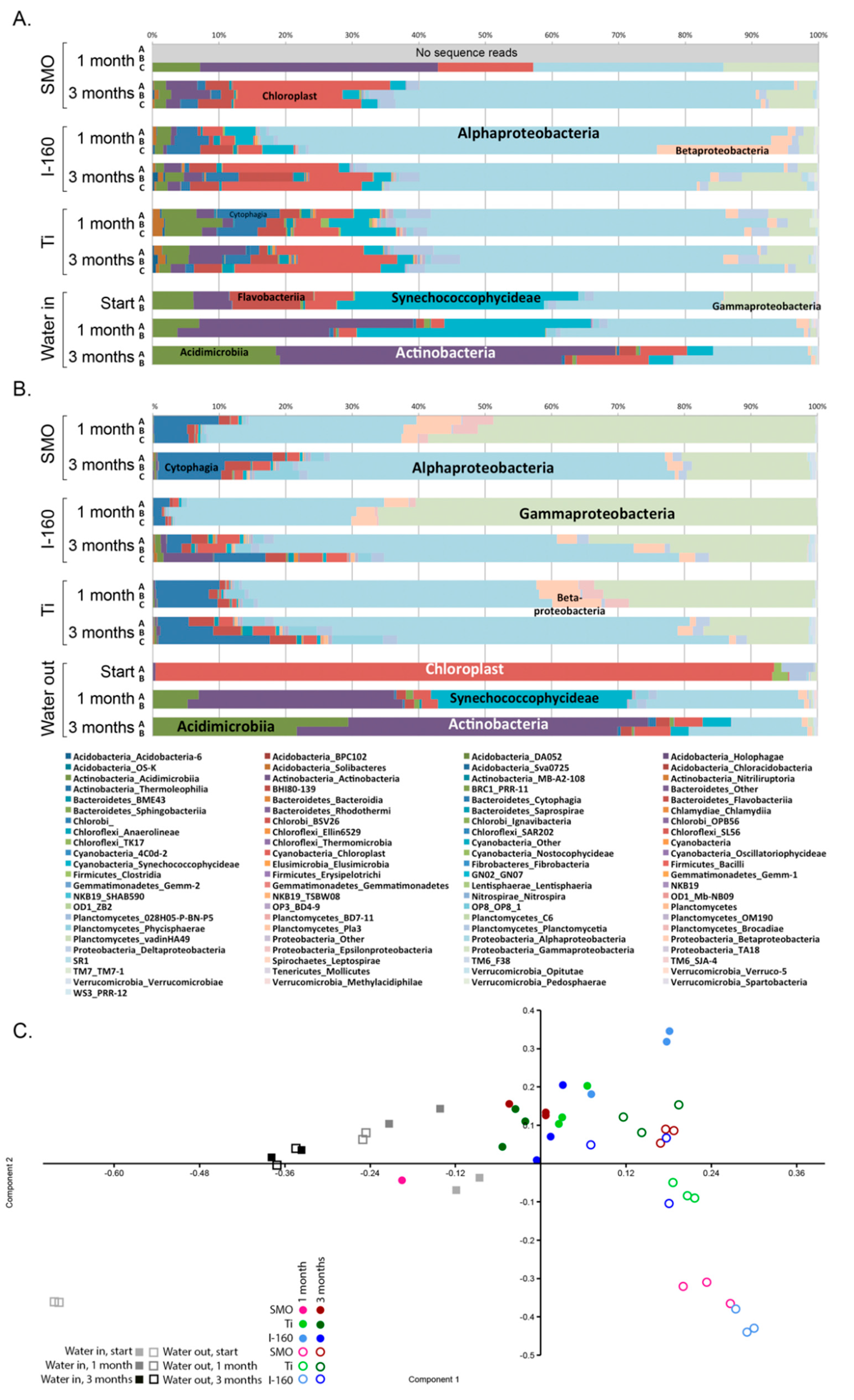
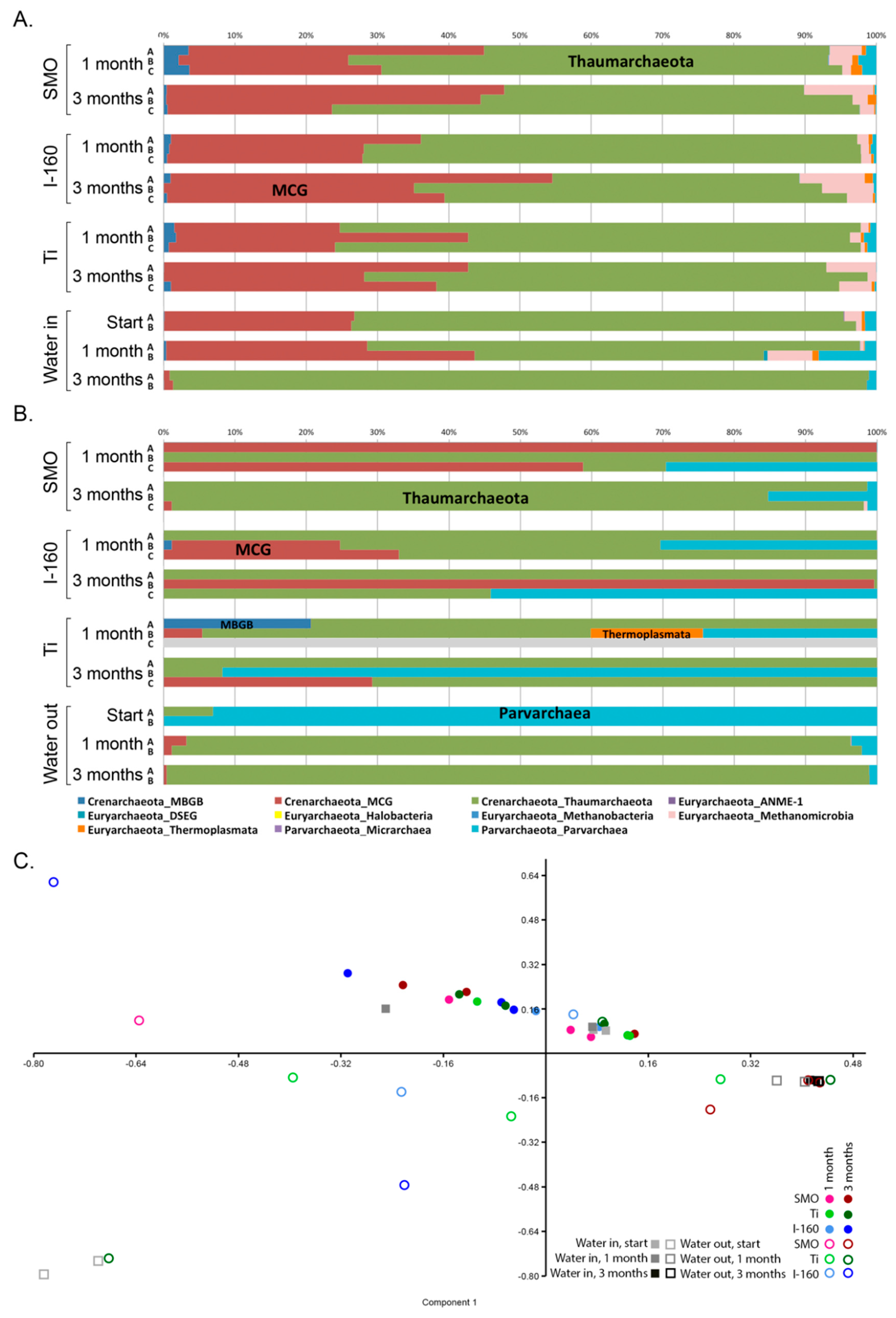
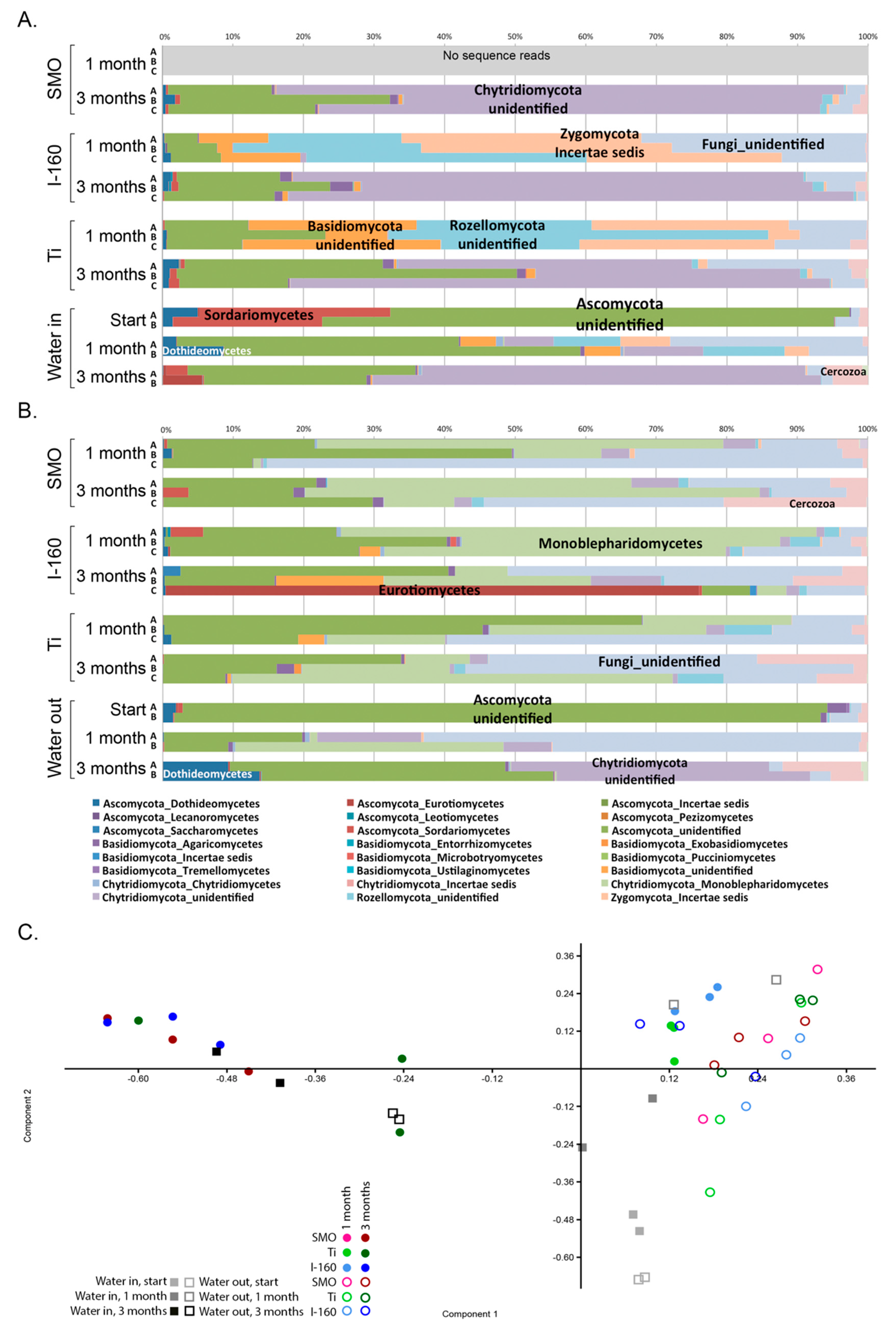
2.1.4. Microbial Community Composition
2.2. Inorganic Fouling
2.3. Materials and Their Performance
2.4. Seawater
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Experiment Setup
4.3. Surface Characterization
4.4. DNA Extraction
4.5. Quantitative PCR
4.6. Amplicon Library
4.7. Sequence Processing and Analysis
4.8. Statistics Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| BLAST | Basic Local Alignment Search Tool |
| bp | Base pair (in DNA) |
| DNA | Deoxyribonucleic acid |
| FE-SEM | Field emission scanning electron microscopy |
| EDS | Energy-dispersive spectroscopy |
| ITS | fungal internal transcribed spacer |
| MCG | Miscellaneous crenarchaeotal group |
| OTU | Operational taxonomic unit |
| PCA | Principal component analysis |
| PCR | Polymerase chain reaction |
| STD | Standard deviation |
| Ti | in this paper, Ti-6Al-4V |
| TOC | Total organic carbon |
References
- Fleming, H.C. Microbially Influenced Corrosion of Materials; Heitz, E., Fleming, H.C., Sand, W., Eds.; Springer-Verlag: New York, NY, USA, 1996; pp. 6–14. [Google Scholar]
- Sawall, Y.; Richter, C.; Ramette, A. Effects of eutrophication, seasonality and macrofouling on the diversity of bacterial biofilms in equatorial coral reefs. PLoS ONE 2012, 7, e39951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salta, M.; Wharton, J.A.; Blache, Y.; Stokes, K.R.; Briand, J.F. Marine biofilms on artificial surfaces: Structure and dynamics. Environ. Microbiol. 2013, 15, 2879–2893. [Google Scholar] [CrossRef] [PubMed]
- Cristiani, P.; Perboni, G.; Debenedetti, A. Effect of chlorination on the corrosion of Cu/Ni 70/30 condenser tubing. Electrochim. Acta 2008, 54, 100–107. [Google Scholar] [CrossRef]
- Cristiani, P.; Perboni, P. Antifouling strategies and corrosion control in cooling circuits. Bioelectrochemistry 2014, 97, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Murthy, P.S.; Venkatesan, R.; Nair, K.V.K.; Inbakandan, D.; Jahan, S.S.; Peter, D.M.; Ravindan, M. Evaluation of sodium hypochlorite for fouling control in plate heat exchangers for seawater application. Int. Biodeter. Biodegrad. 2005, 55, 161–170. [Google Scholar] [CrossRef]
- Yebra, D.M.; Kiil, S.; Dam-Johansen, K. Antifouling technology—Past, present and future steps towards efficient and environmentally friendly antifouling coatings. Prog. Org. Coat. 2004, 50, 75–104. [Google Scholar] [CrossRef]
- Ferrari, M.; Benedetti, A. Superhydrophobic surfaces for applications in seawater. Adv. Coll. Interf. Sci. 2015, 222, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Gurrappa, I. Characterization of titanium alloy Ti-6Al-4V for chemical, marine and industrial applications. Mater. Charact. 2003, 51, 131–139. [Google Scholar] [CrossRef]
- Hedrich, S.; Schlömann, M.; Johnson, D.B. The iron-oxidizing proteobacteria. Microbiology 2011, 157, 1551–1564. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.W.; Wu, M.; Oren, A. The Family Kordiimonadaceae. In The Prokaryotes; Springer: Berlin, Germany, 2014; pp. 307–312. [Google Scholar]
- Tonon, L.A.C.; Moreira, A.P.B.; Thompson, F. The Family Erythrobacteraceae. In The Prokaryotes; Springer: Berlin, Germany, 2014; pp. 213–235. [Google Scholar]
- Geszvain, K.; Butterfield, C.; Davis, R.; Madison, A.; Lee, S.; Parker, D.; Soldatova, A.; Spiro, T.G.; Luther, G.W.; Tebo, B. The molecular biogeochemistry of manganese (II) oxidation. Biochem. Soc. Trans. 2012, 40, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Maki, J.S.; Rittschof, D.; Teo, S.L.M. Early marine bacterial biofilm on a copper-based antifouling paint. Int. Biodeter. Biodegrad. 2013, 83, 71–76. [Google Scholar] [CrossRef]
- Holmström, C.; Egan, S.; Franks, A.; McCloy, S.; Kjelleberg, S. Antifouling activities expressed by marine surface associated Pseudoalteromonas species. FEMS Microbiol. Ecol. 2002, 41, 47–58. [Google Scholar] [CrossRef]
- Bernbom, N.; Ng, Y.Y.; Olsen, S.M.; Gram, L. Pseudoalteromonas spp. Serve as Initial Bacterial Attractants in Mesocosms of Coastal Waters but Have Subsequent Antifouling Capacity in Mesocosms and when Embedded in Paint. Appl. Environ. Microbiol. 2013, 79, 6885–6893. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.; Song, Z.; Yang, L.; Jiang, J.; He, J. Effect of marine Pseudoalteromonas sp. on the microstructure and corrosion behaviour of 2205 duplex stainless steel. Corros. Sci 2014, 84, 103–112. [Google Scholar] [CrossRef]
- Liang, R.; Grizzle, R.S.; Duncan, K.E.; McInerney, M.J.; Suflita, J.M. Roles of thermophilic thiosulfate-reducing bacteria and methanogenic archaea in the biocorrosion of oil pipelines. Front. Microb. 2014, 5, 89. [Google Scholar] [CrossRef] [PubMed]
- Lenhart, T.R.; Duncan, K.E.; Beech, I.B.; Sunner, J.A.; Smith, W.; Bonifay, V.; Biri, B.; Suflita, J.M. Identification and characterization of microbial biofilm communities associated with corroded oil pipeline surfaces. Biofouling 2014, 30, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Fang, H.H.P.; Ko, B.C.B. Methanogen population in a marine biofilm corrosive to mild steel. Appl. Microbiol. Biotechnol. 2003, 63, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Könneke, M.; Bernhard, A.E.; José, R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Hügler, M.; Sievert, S.M. Beyond the Calvin cycle: Autotrophic carbon fixation in the ocean. Mar. Sci. 2011, 3, 261–289. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, K.G.; Schreiber, L.; Petersen, D.G.; Kjeldsen, K.U.; Lever, M.A.; Steen, A.D.; Stepanauskas, R.; Richter, M.; Kleindienst, S.; Lenk, S.; et al. Predominant archaea in marine sediments degrade detrital proteins. Nature 2013, 496, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Wang, L.; Li, L.; He, Y.; Yang, M.; Ding, Z. Effect of the fungus, Aspergillus niger, on the corrosion behaviour of AZ31B magnesium alloy in artificial seawater. Corros. Sci. 2015, 98, 249–259. [Google Scholar] [CrossRef]
- Naranjo, L.; Pernía, B.; Inojosa, Y.; Rojas, D.; D’Anna, L.; González, M.; Sisto, Á. First Evidence of Fungal Strains Isolated and Identified from Naphtha Storage Tanks and Transporting Pipelines in Venezuelan Oil Facilities. Adv. Microbiol. 2015, 5, 143–154. [Google Scholar] [CrossRef]
- Miyata, N.; Tani, Y.; Maruo, K.; Tsuno, H.; Sakata, M.; Iwahori, K. Manganese (IV) oxide production by Acremonium sp. strain KR21–2 and extracellular Mn (II) oxidase activity. Appl. Environ. Microbiol. 2006, 72, 6467–6473. [Google Scholar] [CrossRef] [PubMed]
- Little, B.; Staehle, R.; Davis, R. Fungal influenced corrosion of post-tensioned cables. Int. Biodeterior. Biodegrad. 2001, 47, 71–77. [Google Scholar] [CrossRef]
- Pereira, V.J.; Marques, R.; Marques, M.; Benoliel, M.J.; Crespo, M.B. Free chlorine inactivation of fungi in drinking water sources. Water Res. 2013, 47, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Jelic-Mrcelic, G.; Sliskovic, M.; Antolic, B. Biofouling communities on test panels coated with TBT and TBT-free copper based antifouling paints. Biofouling 2006, 22, 293–302. [Google Scholar] [CrossRef] [PubMed]
- De Messano, L.V.; Sathler, L.; Reznik, L.Y.; Coutinho, R. The effect of biofouling on localized corrosion of the stainless steels N08904 and UNS S32760. Int. Biodeterior. Biodegrad. 2009, 63, 607–614. [Google Scholar] [CrossRef]
- Coetser, S.E.; Cloete, T.E. Biofouling and biocorrosion in industrial water systems. Crit. Rev. Microbiol. 2005, 31, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Valster, R.M.; Wullings, B.A.; van der Kooij, D. Detection of protozoan hosts for Legionella pneumophila in engineered water systems by using a biofilm batch test. Appl. Environ. Microbiol. 2010, 76, 7144–7153. [Google Scholar] [CrossRef] [PubMed]
- Al-Ahmad, M.; Abdul Aleem, F.A.; Mutiri, A.; Ubaisy, A. Biofouling in RO membrane systems. Part 1: Fundamentals and control. Desalination 2000, 132, 173–179. [Google Scholar] [CrossRef]
- Demadis, K.D.; Neofotistou, E.; Mavredaki, E.; Tsiknakis, M.; Sarigiannidou, E.-M.; Katarachia, S.D. Inorganic foulants in membrane systems: Chemical control strategies and the contribution of “green chemistry”. Desalination 2005, 179, 281–295. [Google Scholar] [CrossRef]
- Euvrard, M.; Hadi, L.; Foissy, A. Influence of PPCA (phosphinopolycarboxylic acid) and DETPMP (diethylenetriaminepentamethylenephosphonic acid) on silica fouling. Desalination 2007, 205, 114–123. [Google Scholar] [CrossRef]
- Hellio, C.; Yebra, D. Advances in Marine Antifouling Coatings and Technologies, 1st ed.; Elsevier: Cambridge, UK, 2009. [Google Scholar]
- Al-Turaif, H.A. Surface morphology and chemistry of epoxy-based coatings after exposure to ultraviolet radiation. Prog. Org. Coat. 2013, 76, 677–681. [Google Scholar] [CrossRef]
- Daghia, F.; Zhang, F.; Cluzel, C.; Ladeveze, P. Thermo-mechano-oxidative behaviour at the ply’s scale: The effect of oxidation on the transverse cracking in carbon-epoxy composites. Compos. Struct. 2015, 134, 602–612. [Google Scholar] [CrossRef]
- Shams El Din, A.M.; El-Dahshan, M.E.; Tag El Din, A.M. Biofilm formation on stainless steels Part 2. The role of seasonal changes, seawater composition and surface roughness. Desalination 2003, 154, 267–276. [Google Scholar] [CrossRef]
- Muyzer, G.; De Waal, E.; Uitterlinden, A. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [PubMed]
- Bano, N.; Ruffin, S.; Ransom, B.; Hollibaugh, J.T. Phylogenetic composition of arctic ocean archaeal assemblages and comparison with antarctic assemblages. Appl. Environ. Microbiol. 2004, 70, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Barns, S.M.; Fundyga, R.E.; Jeffries, M.W.; Pace, N.R. Remarkable archaeal diversity detected in a Yellowstone National Park hot spring environment. Proc. Natl. Acad. Sci. USA 1994, 91, 1609–1613. [Google Scholar] [CrossRef] [PubMed]
- Haugland, R.; Vesper, S. Method of Identifying and Quantifying Specific Fungi and Bacteria. U.S. Patent 6,387,652, 14 May 2002. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucl. Acids Res. 2012, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Pena, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.; Gish, W.; Miller, W.; Myers, E.; Lipman, D. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria, cfu/mL | Archaeons, cells/mL | Fungi, spores/mL | ||||
|---|---|---|---|---|---|---|
| Non-Chlorinated | Chlorinated | Non-Chlorinated | Chlorinated | Non-Chlorinated | Chlorinated | |
| beginning | 7.7 × 106 | 1.8 × 103 | 3.2 × 103 | <1 | 2300 | 3 |
| 1 month | 8.3 × 106 | 5.6 × 106 | 8.8 × 103 | 6.8 × 103 | 500 | 380 |
| 3 months | 5.8 × 106 | 8.8 × 106 | 1.3 × 104 | 6.8 × 103 | 360 | 710 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajala, P.; Bomberg, M.; Huttunen-Saarivirta, E.; Priha, O.; Tausa, M.; Carpén, L. Influence of Chlorination and Choice of Materials on Fouling in Cooling Water System under Brackish Seawater Conditions. Materials 2016, 9, 475. https://doi.org/10.3390/ma9060475
Rajala P, Bomberg M, Huttunen-Saarivirta E, Priha O, Tausa M, Carpén L. Influence of Chlorination and Choice of Materials on Fouling in Cooling Water System under Brackish Seawater Conditions. Materials. 2016; 9(6):475. https://doi.org/10.3390/ma9060475
Chicago/Turabian StyleRajala, Pauliina, Malin Bomberg, Elina Huttunen-Saarivirta, Outi Priha, Mikko Tausa, and Leena Carpén. 2016. "Influence of Chlorination and Choice of Materials on Fouling in Cooling Water System under Brackish Seawater Conditions" Materials 9, no. 6: 475. https://doi.org/10.3390/ma9060475
APA StyleRajala, P., Bomberg, M., Huttunen-Saarivirta, E., Priha, O., Tausa, M., & Carpén, L. (2016). Influence of Chlorination and Choice of Materials on Fouling in Cooling Water System under Brackish Seawater Conditions. Materials, 9(6), 475. https://doi.org/10.3390/ma9060475