Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Measurements and Estimations
3. Statistical Analyses
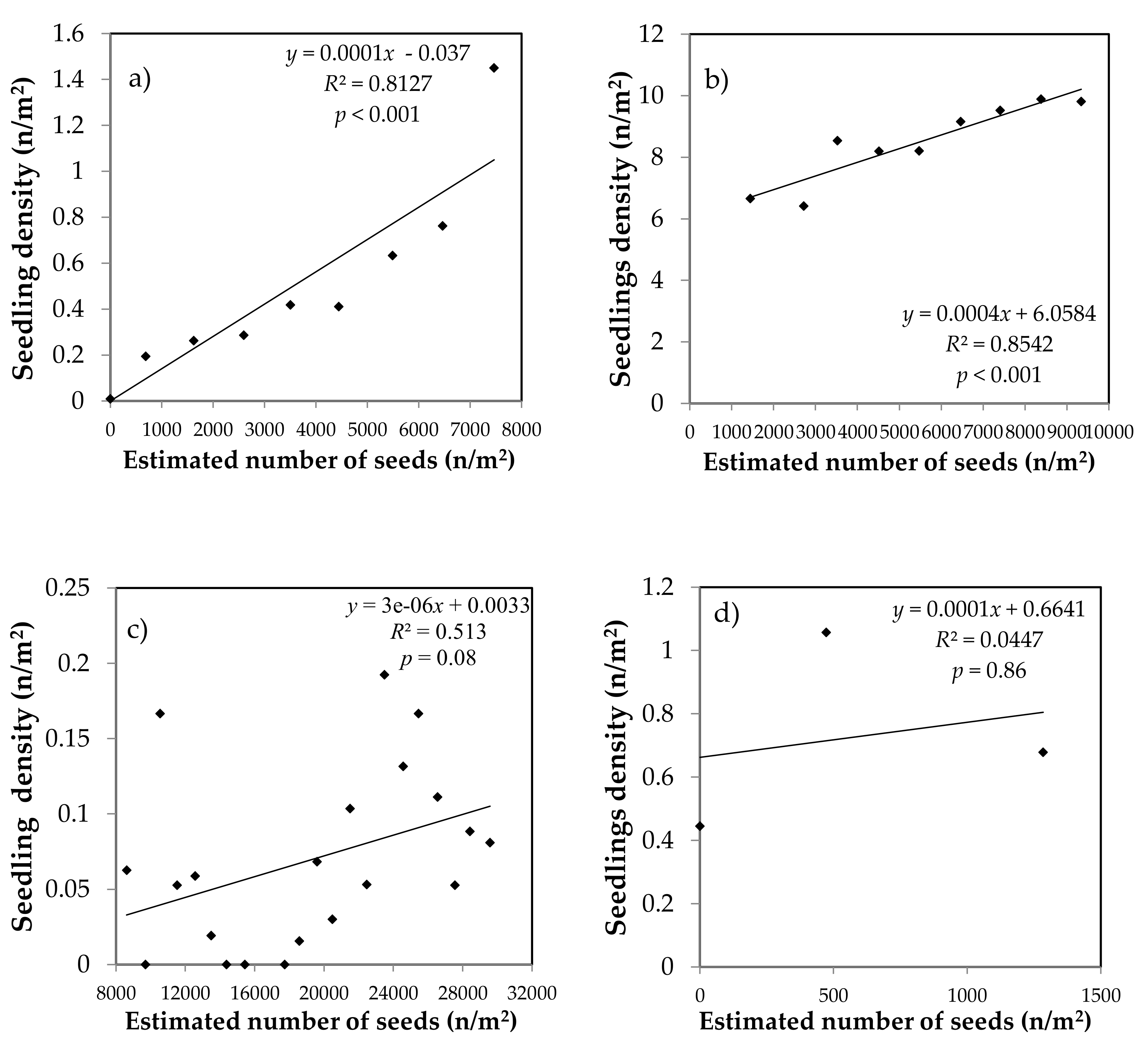
3.1. Relationship between the Estimated Amount of Dispersed Seeds and the Number of Seedlings
3.2. Comparison of Density and Stocking Coefficient of Seedlings
3.3. Percentage of Seedlings per Substrate Type
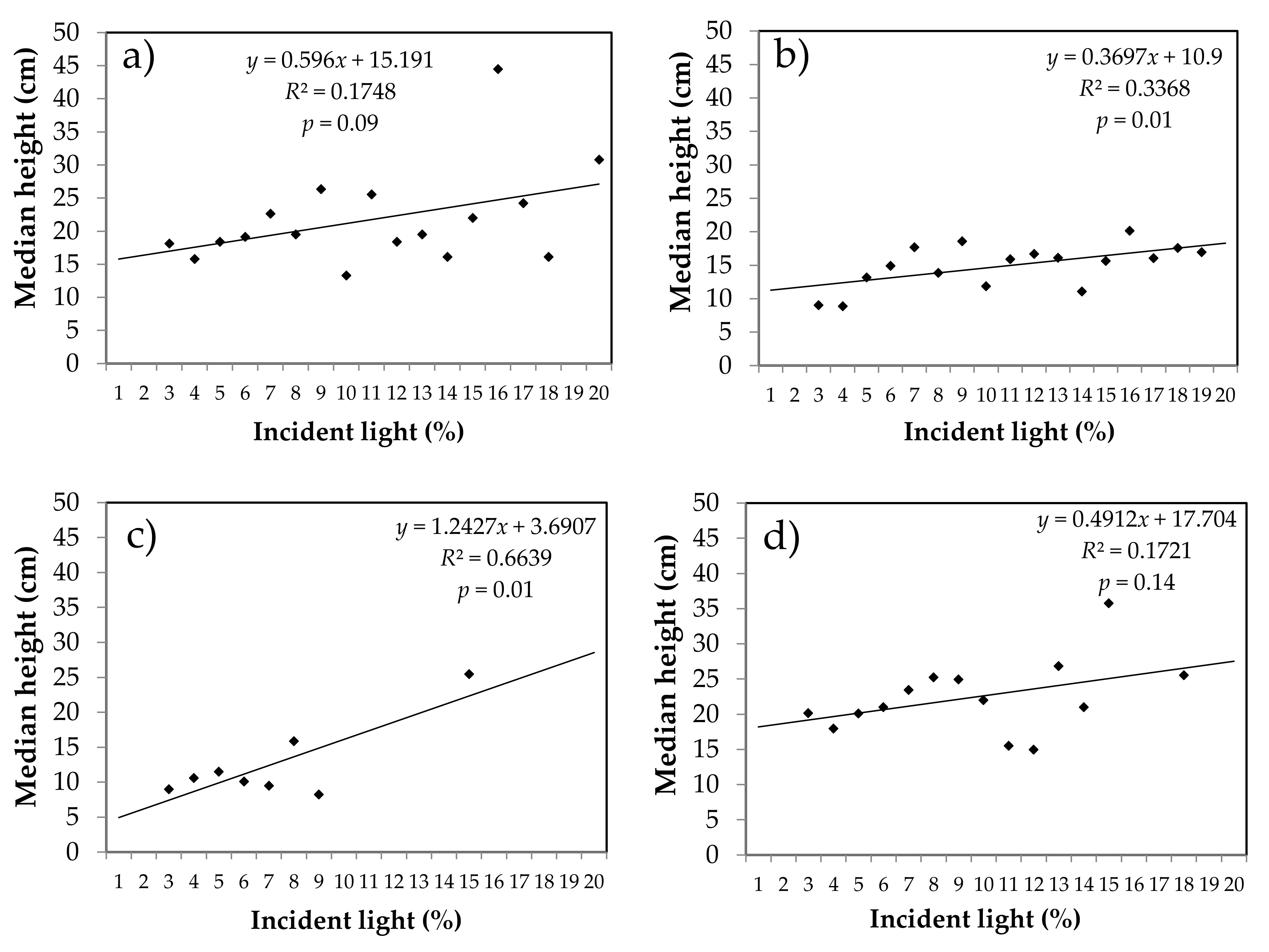
3.4. Relationship between Median Seedling Height and Incident Light
4. Results
4.1. Relationships between the Estimated Number of Dispersed Seeds and the Number of Seedlings
4.2. Stocking Coefficient and Density of Seedlings
4.3. Substrate Type Cover and Seedling Establishment in Plantations and Natural Stands
4.4. Relationship between the Percentage of Incident Light, Density of Seedlings and Their Height
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Olson, M.G.; Meyer, S.R.; Wagner, R.G.; Seymour, R.S. Commercial thinning stimulates natural regeneration in spruce-fir stands. Can. J. For. Res. 2014, 44, 173–181. [Google Scholar] [CrossRef]
- Bauhus, J.; Puettmann, K.; Messier, C. Silviculture for old-growth attributes. For. Ecol. Manag. 2009, 258, 525–537. [Google Scholar] [CrossRef]
- Lundqvist, L. Stand development in uneven-aged sub-alpine Picea abies stands after partial harvest estimated from repeated surveys. Forestry 2004, 77, 120–129. [Google Scholar] [CrossRef]
- Nilson, K.; Lundqvist, L. Effect of stand structure and density on development of natural regeneration in two Picea abies stands in Sweden. Scand. J. For. Res. 2001, 16, 253–259. [Google Scholar] [CrossRef]
- Lundqvist, L.; Chrimes, D.; Elfving, B.; Mörling, T.; Valinger, E. Stand development after different thinnings in two uneved-aged Picea abies forest in Sweden. For. Ecol. Manag. 2007, 238, 141–146. [Google Scholar] [CrossRef]
- Rive, A.C. Enhancing Natural Regeneration of white Spruce (Picea glauca) via Synchronization of a Mast Year with Site Manipulations in Abitibi, Quebec. Master’s Thesis, Biology Department, Concordia University, Montreal, QC, Canada, 2010. [Google Scholar]
- Calogeropoulos, C.; Greene, D.F.; Messier, C.; Brais, S. The effects of harvest intensity and seedbed type on germination and cumulative survivorship of white spruce and balsam fir in northwestern Quebec. Can. J. For. Res. 2004, 34, 1467–1476. [Google Scholar] [CrossRef]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Conifer seedling recruitment in a southeastern canadian boreal forest: The importance of substrate. J. Veg. Sci. 1998, 9, 575–582. [Google Scholar] [CrossRef]
- Messier, C.; Doucet, R.; Ruel, J.-C.; Claveau, Y.; Kelly, C.; Lechowicz, M.J. Functional ecology of advance regeneration in relation to light in boreal forests. Can. J. For. Res. 1999, 29, 812–823. [Google Scholar] [CrossRef]
- Blanco, J.A.; Welham, C.; (Hamish) Kimmins, J.P.; Seely, B.; Mailly, D. Guidelines for modeling natural regeneration in boreal forests. For. Chron. 2009, 85, 427–439. [Google Scholar] [CrossRef]
- Gärtner, S.M.; Lieffers, V.J.; Macdonald, S.E. Ecology and management of natural regeneration of white spruce in the boreal forest. Environ. Rev. 2011, 19, 461–478. [Google Scholar] [CrossRef]
- Greene, D.F.; Johnson, E.A. Estimating the mean annual seed production of trees. Ecology 1994, 75, 642647. [Google Scholar] [CrossRef]
- Greene, D.F. Sexual recruitment of trees in strip cuts in eastern Canada. Can. J. For. Res. 2000, 30, 1256–1263. [Google Scholar] [CrossRef]
- Hély, C.; Bergeron, Y.; Flannigan, M.D. Coarse woody debris in the southeastern Canadian boreal forest: Composition and load variations in relation to stand replacement. Can. J. For. Res. 2000, 30, 674–687. [Google Scholar] [CrossRef]
- Christy, E.J.; Mack, R.N. Variation in demography of juvenile Tsuga heterophylla across the substratum mosaic. J. Ecol. 1984, 72, 75–91. [Google Scholar] [CrossRef]
- Kuuluvainen, T.; Kalmari, R. Regeneration microsites of Picea abies seedlings in a windthrow area of boreal old-growth forest in southern Finland. Ann. Bot. Fenn. 2003, 40, 401–413. [Google Scholar]
- Simard, M.-J.; Bergeron, Y.; Sirois, L. Substrate and litterfall effects on conifer seedling survivorship in southern boreal stands of Canada. Can. J. For. Res. 2003, 33, 672–681. [Google Scholar] [CrossRef]
- Carswell, F.E.; Doherty, J.E.; Allen, R.B.; Brignall-Theyer, M.E.; Richardson, S.J.; Wiser, S.K. Quantification of the effects of aboveground and belowground competition on growth of seedlings in a conifer– angiosperm forest. For. Ecol. Manag. 2012, 269, 188–196. [Google Scholar] [CrossRef]
- Niensteadt, H.; Zasada, J.C. Picea glauca (Moench) Voss. White spruce. In Silvics of North America; Burns, R.M., Honkola, B.H., Eds.; USDA Forest Service Agriculture Handbook; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, ISBN -13 978-0160271458. [Google Scholar]
- Frank, R.M. Abies balsamea (L.) Mill. Balsam fir. In Silvics of North America; Burns, R.M., Honkola, B.H., Eds.; USDA Forest Service Agriculture Handbook; USDA Forest Service: Washington, DC, USA, 1990; Volume 1, Conifers; pp. 26–35. ISBN -13: 978-0160271458. [Google Scholar]
- Groupe d’experts sur la Sylviculture Intensive de Plantations (GESIP). La Sylviculture Intensive de Plantations dans un Contexte d’aménagement écosystémique; Gouvernement du Québe: Québec City, QC, Canada, 2013; 112p, Rapport du groupe d’experts, sous la direction de M. Barrette et M. Leblanc; Québec; Numéro de publication: DAEF-0354; ISBN 978-2-550-69378-9. [Google Scholar]
- Kneeshaw, D.D.; Bergeron, Y. Ecological factors affecting the abundance of advance regeneration in Quebec’s southwestern boreal forest. Can. J. For. Res. 1996, 26, 888–898. [Google Scholar] [CrossRef]
- Robert, E.; Brais, S.; Harvey, B.D.; Greene, D. Seedling establishment and survival on decaying logs in boreal mixedwood stands following a mast year. Can. J. For. Res. 2012, 42, 1446–1455. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Koubaa, A. Softwoods of Eastern Canada; Special publication 1917-3539; SP-526F FPInnovations Institute: Vancouver, BC, Canada, 2009; ISBN 978-0-86488-538-8. [Google Scholar]
- Klopcic, M.; Bončina, A. Recruitment of tree species in mixed selection and irregular shelterwood forest stands. Ann. For. Sci. 2012, 69, 915–925. [Google Scholar] [CrossRef]
- Schütz, J.P.; Pommerening, A. Can Douglas fir (Pseudotsuga menziesii (Mirb.) Franco.) sustainable grow in complex forest structures? For. Ecol. Manag. 2013, 303, 175–183. [Google Scholar] [CrossRef]
- Saucier, J.-P.; Robitaille, A.; Grondin, P.; Bergeron, J.-F.; Gosselin, J. Les Régions Écologiques du Québec Meridional. (4e version). 2011. Available online: https://mffp.gouv.qc.ca/forets/inventaire/pdf/carte-regions-ecologiques.pdf (accessed on 20 February 2019).
- Alemdag, I.S. National site-index and height growth curves for white spruce growing in natural stands in Canada. Can J. For. Res. 1991, 21, 1466–1474. [Google Scholar] [CrossRef]
- Prégent, G. Tarif de Cubage, Tables de Rendement et Modèles de Croissance pour les Plantations d’épinette Blanche au Québec; Mémoire de recherche forestière No. 160; Ministère des Ressources naturelles et de la Faune, Direction de la recherche forestière, Gouvernement du Québec: Québec City, QC, Canada, 2010; 76p, ISBN 978-2-550-60589-8. [Google Scholar]
- Bédard, S. L’estimation du Potential de Croissance des Stations Forestières: Exemple du Sous-Domaine de la Sapinière à Bouleau Jaune de l’Est du Québec; Mémoire de recherche forestière No. 140; Forêt Québec. Direction de la recherche forestière: Québec City, QC, Canada, 2002; ISBN 2-550-38688-4. [Google Scholar]
- Ministère des Ressources naturelles. Le Guide Sylvicole du Québec—Tome 2—Les concepts et l’application de la Silviculture; Québec, Les Publications du Québec: Québec City, QC, Canada, 2013; 709p.
- Paquette, A.; Bouchard, A.; Cogliastro, A. A less restrictive technique for the estimation of understorey light under variable weather conditions. For. Ecol. Manag. 2007, 242, 800–804. [Google Scholar] [CrossRef]
- Royston, P. Remark AS R94: A remark on Algorithm AS 181: The W test for normality. Appl. Stat. 1995, 44, 547–551. [Google Scholar] [CrossRef]
- Zuur, A.F.; Saveliev, A.A.; Ieno, E.N. Zero Inflated Models and Generalized Linear Mixed Models with R. Newburgh; Highland Statistics Limited: Newburgh, UK, 2012; ISBN 9780957174108. [Google Scholar]
- Chou, Y.M.; Polansky, A.M.M.R.L. Transforming non normal data to normality in statistical process control. J. Qual. Technol. 1998, 30, 2. [Google Scholar] [CrossRef]
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2007; p. 38. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-97; The R Development Core Team: Vienna, Austria, 2010. [Google Scholar]
- Warnes, G.R.; Bolker, B.; Lumley, T.; Johnson, R.C.; Contributions from Randall C. Johnson Are Copyright. SAIC-Frederick, Inc. Funded by the Intramural Research Program, of the NIH, National Cancer Institute and Center for Cancer Research under NCI Contract NO1-CO-12400. Gmodels: Various R Programming Tools for Model Fitting. 2005 R Package Version 2.15.0. Available online: http://CRAN.Rproject.org/package=gmodels (accessed on 20 February 2019).
- Prévost, M. Effect of cutting intensity on microenvironmental conditions and regeneration dynamics in yellow birch–conifer stands. Can. J. For. Res. 2008, 38, 317–330. [Google Scholar] [CrossRef]
- Cameron, A. Determining the sustainable normal irregular condition: A provisional study on a transformed, irregular mixed species stand in Scotland. Scand. J. For. Res. 2007, 22, 13–21. [Google Scholar] [CrossRef]
- Kuehne, C.; Puettmann, K.J. Natural regeneration in thinned Douglas-stands in western Oregon. J. Sustain. For. 2008, 27, 246–274. [Google Scholar] [CrossRef]
- Carter, G.A.; Smith, W.K. Influence of shoot structure on light interception and photosynthesis in conifers. Plant Physiol. 1985, 79, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Lieffers, V.J.; Messier, C.; Stadt, K.J.; Gendron, F.; Comeau, P.G. Predicting and managing light in the understrory of boreal forests. Can. J. For. Res. 1999, 29, 796–811. [Google Scholar] [CrossRef]
- Noguchi, M.; Okuda, S.; Miyamoto, K.; Itou, T.; Inagaki, Y. Composition, size structure and local variation of naturally regenerated broad-leaved tree species in hinoki cypress plantations: A case study in Shikoku, southwestern Japan. Forestry 2011, 84, 493–504. [Google Scholar] [CrossRef]
- Bizzari, L.E.; Collins, C.D.; Brudvig, L.A.; Damschen, E.I. Historical agriculture and contemporary fire frequency alter soil properties in longleaf pine woodlands. For. Ecol. Manag. 2015, 349, 45–54. [Google Scholar] [CrossRef]
- McNabb, D.H.; Startsev, A.D.; Nguyen, H. Soil wetness and traffic level effects on bulk density and airfilled porosity of compacted forest soils. Soil Sci. Soc. Am. J. 2001, 65, 1238–1247. [Google Scholar] [CrossRef]
- Startsev, A.D.; McNabb, D.H. Effects of compaction on aeration and morphology of boreal forest soils in Alberta, Canada. Can. J. Soil Sci. 2009, 89, 45–56. [Google Scholar] [CrossRef]
- McLauchlan, K. The nature and longevity of agricultural impacts on soil carbon and nutrients: A review. Ecosystems 2006, 9, 1364–1382. [Google Scholar] [CrossRef]
- Lieffers, V.J.; Stadt, K.J.; Navratil, S. Age structure and growth of understory white spruce under aspen. Can. J. For. Res. 1996, 26, 1002–1007. [Google Scholar] [CrossRef]
- Leinonen, K.; Leikola, M.; Peltonen, A.; Räsänen, P.K. Kuusen luontainen uudistuminen Pirkka-Hämeen metsälautakunnassa. Summary: Natural regeneration of Norway spruce in Pirkka-Häme Forestry Board District, southern Finland. Acta For. Fenn. 1989, 209, 1–53. [Google Scholar]
- Chećko, E.; Jaroszewicz, B.; Olejniczak, K.; Kwiatkowska-Falińska, A.J. The importance of coarse woody debris for vascular plants in temperate mixed deciduous forests. Can. J. For. Res. 2015, 45, 1154–1163. [Google Scholar] [CrossRef]
- Cornett, M.W.; Reich, P.B.; Puettmann, K.J.; Frelich, L.E. Seedbed and moisture availability determine safe sites for early Thuja occidentalis (Cupressaceae) regeneration. Am. J. Bot. 2000, 87, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Waldron, R.M. Factors Affecting Natural White Spruce Regeneration on Prepared Seedbeds at the Riding Mountain Forest Experimental Area; Government of Canada, Department of Forestry and Rural Development, Forest Research Laboratory: Winnipeg, MB, Canada, 1966. [Google Scholar]
- Novák, J.; Slodičák, M. Structure and accumulation of litterfall under Norway spruce stands in connection with thinnings. J. For. Sci. 2004, 50, 101–108. [Google Scholar] [CrossRef]
- Eis, S. Development of white spruce and alpine fir seedlings on cut-over areas in the central interior of British Columbia. For. Chron. 1965, 41, 419–431. [Google Scholar] [CrossRef]
- Greene, D.F.; Zasada, J.C.; Sirois, L.; Kneeshaw, D.D.; Morin, H.; Charron, I.; Simard, M.-J. A review of the regeneration dynamics of North American boreal forest tree species. Can. J. For. Res. 1999, 29, 824–839. [Google Scholar] [CrossRef]
- Lundqvist, L.; Nilson, K. Regeneration dynamics in an uneven-aged virgin Norway spruce forest in northern Sweden. Scand. J. For. Res. 2007, 22, 304–309. [Google Scholar] [CrossRef]
- Greene, D.F.; Johnson, E.A. Seed mass and early survivorship of tree species in upland clearings and shelterwood. Can. J. For. Res. 1998, 28, 1307–1316. [Google Scholar] [CrossRef]
- Place, I.C.M. The Influence of Seed-Bed Conditions on the Regeneration of Spruce and Balsam Fir; Canadian Department of Northern Affairs and National Resources, Forestry Branch: Gatineau, QC, Canada, 1955. [Google Scholar]
- Wang, G.G.; Kemball, K.J. Balsam fir and white spruce seedling recruitment in response to understory release, seedbed type, and litter exclusion in trembling aspen stands. Can. J. For. Res. 2005, 35, 667–673. [Google Scholar] [CrossRef]
- Hellum, A.K. Grading seed by weight in white spruce. Tree Planters’ Notes 1976, 27, 16–24. [Google Scholar]
- Duchesneau, R.; Morin, H. Early seedling demography in balsam fir seedling banks. Can. J. For. Res. 1999, 29, 1502–1509. [Google Scholar] [CrossRef]
- Dumais, D.; Prévost, M. Management for red spruce conservation in Québec: The importance of some physiological and ecological characteristics—A review. For. Chron. 2007, 83, 378–392. [Google Scholar] [CrossRef]
- Parent, S.; Messier, C. Effets d’un gradient de lumière sur la croissance en hauteur et la morphologie de la cime du sapin baumier régénéré naturellement. Can. J. For. Res. 1995, 25, 878–885. [Google Scholar] [CrossRef]
- Weber, A.; Leckie, S.; Kimmins, J.P.; Gilbert, B.; Blanco, J.A.; Lo, Y.-H. Survival and growth as measures of shade tolerance of planted western redcedar, western hemlock and amabilis fir seedlings in hemlock-fir forests of northern Vancouver Island. For. Ecol. Manag. 2017, 386, 13–21. [Google Scholar] [CrossRef]
- Malcolm, D.C.; Mason, W.L.; Clarke, G.C. The transformation of conifer forests in Britain—Regeneration gap size and silvicultural systems. For. Ecol. Manag. 2001, 151, 7–23. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Site | Age range of Dominant Tree n = 6 | Years after Partial Cut | Stem Removal | Basal Area (m2/ha) | Mean Diameter at Breast Height (cm) | Stems/ha | Seedlings/ha | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % | Spruce | Fir | Spruce | Fir | Spruce | Fir | Spruce | Fir | ||||
| Natural | 1 | 37–40 | 10 | 14.0 | - | 37(0.5) | - | 15(4.1) | − | 1850 | 0 | 4000(4460) |
| 2 | 52–65 | 11 | 67.7 | 3(0) | 26(0.4) | 20 | 16(2.7) | 75(35) | 1325(247) | 875(3833) | 9575(10595) | |
| 3 | 51–71 | 10 | 45.5 | 5(0.5) | 38(0.6) | 20(3.2) | 16(3.4) | 150(141) | 1800(424) | 500(1329) | 4575(6638) | |
| 4 | 46–52 | 9 | 38.2 | 4(0.4) | 27(0.4) | 18(2.6) | 15(3.1) | 125(106) | 1400(247) | 575(1771) | 5300(10470) | |
| 5 | 37–52 | 9 | 36.9 | 16(0.7) | 15(0.7) | 15(4.2) | 17(4.5) | 700(494) | 775(671) | 350(1067) | 6125(6829) | |
| 6 | 48–60 | 9 | 53.0 | 2(0.3) | 25(0.7) | 21(2.3) | 16(4.7) | 75(35) | 1100(212) | 850(3019) | 11,075(16762) | |
| 7 | 55–74 | 10 | 27.9 | 12(1.0) | 26(1.0) | 20(4.7) | 19(5.6) | 300(353) | 875(318) | 2825(5920) | 16,750(16864) | |
| 8 | 62–71 | 9 | 52.1 | 5(1.0) | 36(0.5) | 20(4.7) | 14(4.1) | 150 | 2000 | 550(1161) | 16,850(14575) | |
| 9 | 60–85 | 9 | 11.6 | 7(1.1) | 32(0.8) | 19(6.9) | 16(5.1) | 200(212) | 1350(282) | 550(2089) | 14,425(14190) | |
| 10 | 60–77 | 14 | 73.5 | 4(1.4) | 26(0.7) | 20(6.2) | 19(4.6) | 75(35) | 825(212) | 1625(2667) | 42,050(29602) | |
| 11 | 73–104 | 10 | 74.0 | 8(1.2) | 26(0.7) | 25(4.9) | 18(3.6) | 150(0) | 975(106) | 1225(2809) | 25,950(22551) | |
| 12 | 70–75 | 23 | 60.4 | 7(0.9) | 33(0.7) | 20(5.4) | 18(3.6) | 200(70) | 1225(105) | 200(681) | 3200(8665) | |
| Plantation | 131 | 82 | 30–152 | 42.0 | 29(1.4) | 3(0.3) | 24(5.7) | 10(2.1) | 675(309) | 150(238) | 225(976) | 9350(10000) |
| 14 | 57 | 16–62 | 52.6 | 40(1) | - | 21(5.6) | - | 950(176) | - | 550(1899) | 1650(2664) | |
| 15 | 49 | 8 | 44.3 | 34(0.6) | - | 16(3.3) | - | 1475(247) | - | 0 | 1750(3380) | |
| 16 | 52 | 19−82 | 50.0 | 35(1.4) | - | 22(6.5) | - | 825(196) | - | 0 | 2150(3693) | |
| 17 | 57 | 12 | 65.6 | 29(1.3) | - | 25(6.2) | - | 650(212) | - | 50(351) | 150(596) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gagné, L.; Sirois, L.; Lavoie, L. Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands. Forests 2019, 10, 221. https://doi.org/10.3390/f10030221
Gagné L, Sirois L, Lavoie L. Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands. Forests. 2019; 10(3):221. https://doi.org/10.3390/f10030221
Chicago/Turabian StyleGagné, Laurent, Luc Sirois, and Luc Lavoie. 2019. "Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands" Forests 10, no. 3: 221. https://doi.org/10.3390/f10030221
APA StyleGagné, L., Sirois, L., & Lavoie, L. (2019). Seed Rain and Seedling Establishment of Picea glauca and Abies balsamea after Partial Cutting in Plantations and Natural Stands. Forests, 10(3), 221. https://doi.org/10.3390/f10030221