Productivity, Growth Patterns, and Cellulosic Pulp Properties of Hybrid Aspen Clones

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Location, Soil, and Climatic Condition
2.2. Planting Material
2.3. Study Design
2.4. Measurement of Tree Characteristics
2.5. Properties of Cellulosic Pulp
2.6. Properties of Paper Sheets
2.7. Data Analysis
3. Results
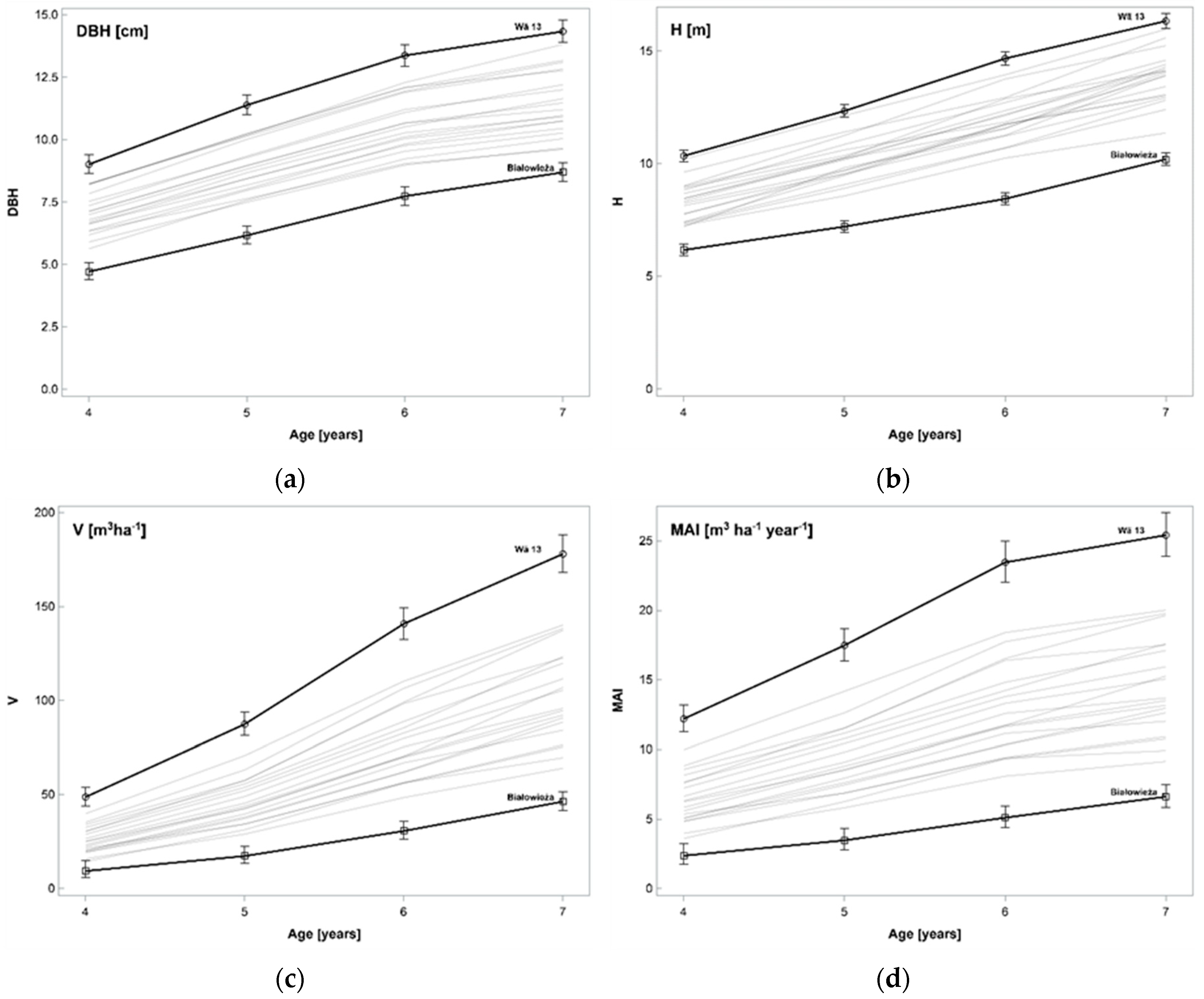
3.1. Biometric Characteristics and Productivity of Clones
3.2. Biometric Characteristics and Productivity for Species Combination
3.3. Properties of Cellulosic Pulp and Paper
3.4. Correlations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Worrell, R. European aspen (Populus tremula L.): A review with particular reference to Scotland I. Distribution, ecology and genetic variation. Forestry 1995, 68, 93–105. [Google Scholar] [CrossRef]
- Richardson, J.; Isebrands, J.G.; Ball, J.B. Ecology and Physiology of Poplars and Willows. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; FAO: Rome, Italy, 2014; pp. 92–123. [Google Scholar]
- Yu, Q.; Pulkkinen, P.; Rautio, M.; Haapanen, M.; Alén, R.; Stener, L.G.; Beuker, E.; Tigerstedt, P.M.A. Genetic control of wood physicochemical properties, growth, and phenology in hybrid aspen clones. Can. J. For. Res. 2001, 31, 1348–1356. [Google Scholar] [CrossRef]
- Bajpai, P. Pulp and Paper Industry: Energy Conservation; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Hart, J.F.; de Araujo, F.; Thomas, B.R.; Mansfield, S.D. Wood quality and growth characterization across intra- and inter-specific hybrid aspen clones. Forests 2013, 4, 786–807. [Google Scholar] [CrossRef]
- Einspahr, D.W.; Winton, L.L. Genetics of quaking aspen. Aspen Bibliogr. Paper 1976, 5026, 1–23. [Google Scholar]
- Einspahr, D.W. Production and utilization of triploid hybrid aspen (Populus tremuloides, Populus tremula). Iowa State J. Resour. 1984, 58, 401–409. [Google Scholar]
- Li, B.; Wyckoff, G.W.; Einspahr, D.W. Hybrid aspen performance and genetic gains. North J. Appl. For. 1993, 10, 117–122. [Google Scholar]
- Melchior, G.H. Breeding of aspen and hybrid aspen and their importance for practical use. Allg. For. Jagdztg. 1985, 156, 112–122. [Google Scholar]
- Zajączkowski, K. Hodowla Lasu: Plantacje Drzew Szybko Rosnących; PWRiL: Warsaw, Poland, 2013. [Google Scholar]
- Baranwal, V.K.; Mikkilineni, V.; Zehr, U.B.; Tyagi, A.K.; Kapoor, S. Heterosis: Emerging ideas about hybrid vigour. J. Exp. Bot. 2012, 63, 6309–6314. [Google Scholar] [CrossRef] [PubMed]
- Lippman, Z.B.; Zamir, D. Heterosis: Revisiting the magic. Trends Genet. 2007, 23, 60–66. [Google Scholar]
- Li, B.; Wu, R. Genetic causes of heterosis in juvenile aspen: A quantitative comparison across intra- and inter-specific hybrids. Theor. Appl. Genet. 1996, 93, 380–391. [Google Scholar] [CrossRef]
- Li, B.; Howe, G.T.; Wu, R. Developmental factors responsible for heterosis in aspen hybrids (Populus tremuloides × P. tremula). Tree Physiol. 1998, 18, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Wettstein-Westerheim, W. Die Kreuzungsmethode und die Beschreibung von F1-Bastarden bei Populus. Zeitschrift für Züchtung 1933, 18, 597–626. [Google Scholar]
- Chmielewski, W. Mieszańce Topolowe Hodowli Instytutu Badawczego Leśnictwa i ich Wstępna Ocena. Ph.D. Thesis, Forest Research Institute, Warsaw, Poland, 1966; p. 131. [Google Scholar]
- Jobling, J. Poplars for Wood Production and Amenity. In Forestry Commission Bulletin 92; Forestry Commission: London, UK, 1990. [Google Scholar]
- Liesebach, M.; Schneck, V. Züchtung, Zulassungen, Vermehrung. In Agrarholz—Schnellwachsende Bäume in der Landschaft; Böhm, C., Veste, M., Eds.; Springer: Berlin, Germany, 2018; pp. 119–145. [Google Scholar]
- Dickmann, D.I.; Stuart, K.W. The Culture of Poplars in Eastern North America; Michigan State University: East Lansing, MI, USA, 1983; p. 168. [Google Scholar]
- Näslund, M. Skogsförsöksanstaltens gallringsförsök i tallskog. Meddelanden från Statens Skogsförsöksanstalt 1936, 29, 1–169. [Google Scholar]
- Wróblewski, L.; Zasada, M. Wzory do określania miąższości grubizny dla modrzewia, osiki, grabu, topoli i lipy. Sylwan 2001, 11, 71–79. [Google Scholar]
- Czuraj, M. Tablice miąższości Kłód Odziomkowych i Drzew Stojących; PWRiL: Warsaw, Poland, 1991. [Google Scholar]
- Modrzejewski, K.; Olszewski, J.; Rutkowski, J. Analysis in Papermaking Industry; Editorial Office of the Lodz University of Technology: Łódź, Poland, 1969; pp. 60–89, 206–250. [Google Scholar]
- Przybysz, K.; Malachowska, E.; Martyniak, D.; Iłowska, J.; Kalinowska, H.; Przybysz, P. Yield of pulp, dimensional properties of fibers, and properties of paper produced from fast growing trees and grasses. BioResources 2018, 13, 1372–1387. [Google Scholar] [CrossRef]
- Johnsson, H. Hybridaspens ungdomsutveckling och ett försök till framtidsprognos. Svenska Skogsvårdsföreningens Tidskrift 1953, 51, 73–96. [Google Scholar]
- Jakobsen, B. Hybrid aspen (Populus tremula L. × Populus tremuloides Michx.). Forstlige Forsøgsvæsen Danmark 1976, 34, 317–338. [Google Scholar]
- Rytter, L. Nutrient content in stems of hybrid aspen as affected by tree age and tree size, and nutrient removal with harvest. Biomass Bioenergy 2002, 23, 13–25. [Google Scholar] [CrossRef]
- Karačić, A.; Verwijst, T.; Weih, M. Above-ground woody biomass production of short-rotation Populus plantations on agricultural land in Sweden. Scand. J. For. Res. 2003, 18, 427–437. [Google Scholar] [CrossRef]
- Rytter, L.; Stener, L.G. Productivity and thinning effects in hybrid aspen (Populus tremula × P. tremuloides Michx.) stands in southern Sweden. Forestry 2005, 78, 285–295. [Google Scholar] [CrossRef]
- Rytter, L. A management regime for hybrid aspen stands combining conventional forestry techniques with early biomass harvests to exploit their rapid early growth. For. Ecol. Manag. 2006, 236, 422–426. [Google Scholar] [CrossRef]
- Tullus, A.; Rytter, L.; Tullus, T.; Weih, M.; Tullus, H. Short-rotation forestry with hybrid aspen (Populus tremula L. × P. tremuloides Michx.) in Northern Europe. Scand. J. For. Res. 2012, 27, 10–29. [Google Scholar] [CrossRef]
- Johnsson, H. Das potential der hybrid aspen produktion (Populus tremula × tremuloides) in Südschweden. Holzzucht 1976, 30, 19–22. [Google Scholar]
- Poker, J. Ökologische und ökonomische Beurteilung unterschiedlicher Produktionsprogramme für Pappeln in Nordwestdeutschland. Ph.D. Thesis, University of Hamburg, Hamburg, Germany, 1984; p. 134. [Google Scholar]
- Perala, D.A. Stand equations for estimating aerial biomass, net productivity, and stem survival of young aspen suckers on good sites. Can. J. For. Res. 1973, 3, 288–292. [Google Scholar] [CrossRef]
- Christersson, L.; Sennerby-Forsse, L. The Swedish programme for intensive short-rotation forests. Biomass Bioenergy 1994, 6, 145–149. [Google Scholar] [CrossRef]
- Biernacik, D.; Filipiak, J.; Miętus, M.; Wójcik, R. Zmienność Warunków Klimatycznych w Polsce po Roku 1951. Rezultaty Projektu Klimat; In Klimat Polski na tle Klimatu Europy: Zmiany i Konsekwencje; Bednorz, E., Kolendowicz, L., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2010; pp. 9–21. [Google Scholar]
- Van de Pol, M.; Ens, B.J.; Heg, D.; Brouwer, L.; Krol, J.; Maier, M.; Exo, K.M.; Oosterbeek, K.; Lok, T.; Eising, C.M.; et al. Do changes in the frequency, magnitude and timing of extreme climatic events threaten the population viability of coastal birds? J. Appl. Ecol. 2010, 47, 720–730. [Google Scholar] [CrossRef]
- Brang, P.; Spathelf, P.; Larsen, J.B.; Bauhus, J.; Bončína, A.; Chauvin, C.; Drössler, L.; García-Güemes, C.; Heiri, C.; Kerr, G.; et al. Suitability of close-to-nature silviculture for adapting temperate European forests to climate change. Forestry 2014, 87, 492–503. [Google Scholar] [CrossRef] [Green Version]
- Dmyterko, E.; Mionskowski, M.; Bruchwald, A. Risk of the wind damage to the forests in Poland on the basis of a stand damage risk model. Sylwan 2015, 159, 361–371. [Google Scholar]
- Beniston, M.; Stephenson, D.B.; Christensen, O.B.; Ferro, C.A.T.; Frei, C.; Goyette, S.; Halsnaes, K.; Holt, T.; Jylhä, K.; Koffi, B.; et al. Future extreme events in European climate: An exploration of regional climate model projections. Clim. Chang. 2007, 81, 71–95. [Google Scholar] [CrossRef]
- Kjellström, E.; Nikulin, G.; Hansson, U.; Strandberg, G.; Ullerstig, A. 21st century changes in the European climate: Uncertainties derived from an ensemble of regional climate model simulations. Tellus Ser. A Dyn. Meteorol. Oceanogr. 2011, 63, 24–40. [Google Scholar] [CrossRef]
- Spiecker, H. Silvicultural management in maintaining biodiversity and resistance of forests in Europe—Temperate zone. J. Environ. Manag. 2003, 67, 55–65. [Google Scholar] [CrossRef]
- Mayer, P.; Brang, P.; Dobbertin, M.; Hallenbarter, D.; Renaud, J.P.; Walthert, L.; Zimmermann, S. Forest storm damage is more frequent on acidic soils. Ann. For. Sci. 2005, 62, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Updegraff, K.; Baughman, M.J.; Taff, S.J. Environmental benefits of cropland conversion to hybrid poplar: Economic and policy considerations. Biomass Bioenergy 2004, 27, 411–428. [Google Scholar] [CrossRef]
- Arevalo, C.B.; Bhatti, J.S.; Chang, S.X.; Sidders, D. Land use change effects on ecosystem carbon balance: From agricultural to hybrid poplar plantation. Agricult. Ecosyst. Environ. 2011, 141, 342–349. [Google Scholar] [CrossRef]
- Rytter, R.M. The potential of willow and poplar plantations as carbon sinks in Sweden. Biomass Bioenergy 2012, 36, 86–95. [Google Scholar] [CrossRef]
- Rytter, L.; Rytter, R.M. Growth and carbon capture of grey alder (Alnus incana (L.) Moench.). For. Ecol. Manag. 2016, 373, 56–65. [Google Scholar] [CrossRef]
- Jaworski, A. Hodowla Lasu: Sposoby Zagospodarowania, Odnowienie Lasu, Przebudowa i Przemiana Drzewostanów; PWRiL: Warsaw, Poland, 2011. [Google Scholar]
- Muhs, H.J. Growth Potential of Aspen Grown under Traditional Forest Management and Some Aspects of Using Aspen as an Alternative Crop in Agriculture. In Elaboration of a Problem-Oriented, Strategic Decision-Matrix for Research Project in Agriculture, Short Agricultural Surpluses; Schliephake, D., Krämer, P., Eds.; Dechema: Frankfurt/Main, Germany, 1986; pp. 94–102. [Google Scholar]
- Niemczyk, M.; Kaliszewski, A.; Jewiarz, M.; Wróbel, M.; Mudryk, K. Productivity and biomass characteristics of selected poplar (Populus spp.) cultivars under the climatic conditions of northern Poland. Biomass Bioenergy 2018, 111, 46–51. [Google Scholar] [CrossRef]
- Liesebach, M.; von Wuehlish, G.; Muhs, H.J. Aspen for short-rotation coppice plantations on agricultural sites in Germany: Effects of spacing and rotation time on growth and biomass production of aspen progenies. For. Ecol. Manag. 1999, 121, 25–39. [Google Scholar] [CrossRef]
- Thykesson, M.; Sjӧberg, L.A.; Ahlgren, P. Paper properties of grass and straw pulps. Ind. Crops Prod. 1998, 7, 351–362. [Google Scholar] [CrossRef]
- Francis, R.C.; Hanna, R.B.; Shin, S.J.; Brown, A.F.; Riemenschneider, D.E. Papermaking characteristics of three Populus clones grown in the north-central United States. Biomass Bioenergy 2006, 30, 803–808. [Google Scholar] [CrossRef]
- Gurnagul, N.; Page, D.H.; Seth, R.S. Dry sheet properties of Canadian hardwood kraft pulps. J. Pulp Pap. Sci. 1990, 16, 36–41. [Google Scholar]
- Niskanen, K. Mechanic of Paper Products; De Gruyter: Berlin, Germany, 2012. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Clone | Cross | Taxon Code | Background | Source |
|---|---|---|---|---|
| IBL 264/2/2 | Populus tremula 19 (Anin) × P. tremuloides 84 (Sweden) | TA × TE | Selected clone within a progeny crossed at the Forest Research Institute | Poland |
| IBL 55/8 | Populus tremula 38 (Białowieża) × P. alba var. nivea 31 (Sadłowice) | TA × A | Selected clone within a progeny crossed at the Forest Research Institute | |
| IBL 91/78 | Populus tremula 19 (Anin) × P. alba 30 (Grodzisk Maz.) | TA × A | Selected clone within a progeny crossed at the Forest Research Institute | |
| IBL 91/2 | Populus tremula 19 (Anin) × P. alba 30 (Grodzisk Maz.) | TA × A | Selected clone within a progeny crossed at the Forest Research Institute | |
| Białowieża (Clone mixture = reference) | Populus tremula | TA | Vegetative progeny of 30 plus trees) originated in wild populations in Białowieża Forest | |
| CA-2-75 | P. tremula × P. alba | TA × A | Germany | |
| Kh 83 | P. tremula (Wedesbüttel 3) × P. tremuloides (North Wisconsin Clone 13) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute | |
| ESCH 5 | P. tremula (Brauna 11) × P. tremuloides (New Hampshire Turesson 141) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Escherode | |
| ESCH 8 | P. tremula (Brauna 11) × P. tremuloides (New Hampshire Turesson 141) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Escherode | |
| L 176 (Se 3) | P. tremula (Brauna 11) × P. tremuloides (New Hampshire Turesson 141) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Seedorf | |
| Se 1 | P. tremula (Brauna 11) × P. tremuloides (New Hampshire Turesson 141) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Seedorf | |
| W 3 | P. tremula (Wedesbüttel 3) | TA | Selected clone in the stand Wedesbüttel (which was originated Tapiau) | |
| 174/10 | P. tremula (Wedesbüttel 18) × P. tremuloides (North Wisconsin Clone 13) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute | |
| Wä 1 | P. tremula (Großdubrau 1) × P. tremuloides (Ontario Maple) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Wächtersbach | |
| L 191 (Wä 14) | P. tremula (Großdubrau 1) × P. tremuloides (Ontario Maple) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Wächtersbach | |
| Astria | P. tremula × P. tremuloides | TA × TE | A triploid clone which is on the market as forest reproductive material of the category “tested” | |
| KH 73 | P. tremula (Wedesbüttel 3) × P. tremuloides (North Wisconsin Clone 13) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Klausheide | |
| Wä 13 | P. tremula (Großdubrau 1) × P. tremuloides (Ontario Maple) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Wächtersbach | |
| Se 4 | P. tremula (Brauna 11) × P. tremuloides (New Hampshire Turesson 141) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Seedorf | |
| Ihlendieksweg (Ihl 174/59) | P. tremula (Wedesbüttel 18) × P. tremuloides (North Wisconsin Clone 13) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute at the site Schmalenbeck | |
| 164A | P. tremula (Wedesbüttel 5) × P. tremuloides (T 13-58) | TA × TE | Selected clone within a progeny crossed at the Thünen Institute |
| Effect | DF | DBH | H | V | MAI |
|---|---|---|---|---|---|
| F Value (p-Value) | |||||
| Clone | 20 | 221.61 (<0.0001) | 359.61 (<0.0001) | 150.27 (<0.0001) | 266.33 (<0.0001) |
| Age | 3 | 2599.52 (<0.0001) | 7119.56 (<0.0001) | 2156.28 (<0.0001) | 1157.05 (<0.0001) |
| Clone × Age | 60 | 1.03 (0.4105) | 6.37 (<.0001) | 1.47 (0.0105) | 1.70 (0.0006) |
| Effect | DBH | H | V | MAI | |
|---|---|---|---|---|---|
| DF | F Value (p-Value) | ||||
| Taxon | 2 | 156.05 (<0.0001) | 274.46 (<0.0001) | 41.74 (<0.0001) | 73.97 (<0.0001) |
| Age | 3 | 817.24 (<0.0001) | 1593.65 (<0.0001) | 584.37 (<0.0001) | 309.17 (<0.0001) |
| Taxon × Age | 6 | 0.79 (0.5775) | 3.66 (0.0013) | 0.20 (0.9769) | 0.21 (0.9730) |
| Clone | Taxon | Kappa Number [-] | Screened Yield [%] | Cellulose [%, d.w] | Hemicellulose [% d.w.] | Lignin [%, d.w] | Extractives [%, d.w] | Minerals [%, d.w] |
|---|---|---|---|---|---|---|---|---|
| Wä 13 | TA × TE | 19.2 ± 0.3 a | 46.8 | 92.4 ± 0.3 a | 3.4 ± 0.6 a | 3.3 ± 0.2 a | 0.2 ± 0.1 a | 0.6 ± 0.2 a |
| IBL 91/78 | TA × A | 18.3 ± 0.1 b | 41.6 | 88.1 ± 0.9 b | 5.1 ± 1.2 b | 5.6 ± 0.8 b | 0.5 ± 0.1 b | 0.7 ± 0.3 a |
| W 3 | TA | 18.0 ± 0.1 b | 43.7 | 86.7 ± 0.6 c | 6.1 ± 1.3 b | 6.0 ± 0.8 b | 0.5 ± 0.1 b | 0.7 ± 0.2 a |
| Clone | Taxon | Mean Arithmetic Fiber Length [µm] | Mean Weighted Fiber Length [µm] | Mean Fiber Width [µm] | Coarseness [mg m−1] | Fine Content [% in Length] |
|---|---|---|---|---|---|---|
| Wä 13 | TA × TE | 619.5 ± 1.7 a | 712.8 ± 3.0 a | 24.6 ± 0.2 a | 0.0836 ± 0.001 a | 11.9 ± 0.4 a |
| IBL 91/78 | TA × A | 622.3 ± 1.0 a | 697.5 ± 2.1 b | 24.4 ± 0.2 a | 0.087 ± 0.001 a | 11.2 ± 0.3 a |
| W 3 | TA | 585 ± 1.8 b | 672.3 ± 1.9 c | 22.7 ± 0.1 b | 0.076 ± 0.001 b | 13.4 ± 0.5 b |
| Clone | Taxon | Apparent Density [g cm−3] | Breaking Length [m] | Strain [%] | Breaking Energy/TEA [J] | Tear Resistance [mN] | Tensile Index [N m g−1] | Tear Index [mN m2 g−1] | WRV [%] Beaten |
|---|---|---|---|---|---|---|---|---|---|
| Wä 13 | TA × TE | 0.79 ± 0.10 a | 11033 ± 367 a | 2.6 ± 0.1 a | 0.20 ± 0.01 a | 227 ± 11 a | 102.7 ± 3.4 a | 2.8 ± 0.1 a | 2.155 ± 0.005 a |
| IBL 91/78 | TA × A | 0.81 ± 0.01 b | 11467 ± 260 a | 2.7 ± 0.2 a | 0.22 ± 0.01 a | 220 ± 18 a | 106.8 ± 2.4 a | 2.8 ± 0.2 a | 2.107 ± 0.008 b |
| W 3 | TA | 0.82 ± 0.10 b | 11483 ± 328 a | 2.4 ± 0.2 a | 0.20 ± 0.02 a | 189 ± 15 b | 106.9 ± 3.1 a | 2.4 ± 0.2 b | 2.088 ± 0.017 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niemczyk, M.; Przybysz, P.; Przybysz, K.; Karwański, M.; Kaliszewski, A.; Wojda, T.; Liesebach, M. Productivity, Growth Patterns, and Cellulosic Pulp Properties of Hybrid Aspen Clones. Forests 2019, 10, 450. https://doi.org/10.3390/f10050450
Niemczyk M, Przybysz P, Przybysz K, Karwański M, Kaliszewski A, Wojda T, Liesebach M. Productivity, Growth Patterns, and Cellulosic Pulp Properties of Hybrid Aspen Clones. Forests. 2019; 10(5):450. https://doi.org/10.3390/f10050450
Chicago/Turabian StyleNiemczyk, Marzena, Piotr Przybysz, Kamila Przybysz, Marek Karwański, Adam Kaliszewski, Tomasz Wojda, and Mirko Liesebach. 2019. "Productivity, Growth Patterns, and Cellulosic Pulp Properties of Hybrid Aspen Clones" Forests 10, no. 5: 450. https://doi.org/10.3390/f10050450
APA StyleNiemczyk, M., Przybysz, P., Przybysz, K., Karwański, M., Kaliszewski, A., Wojda, T., & Liesebach, M. (2019). Productivity, Growth Patterns, and Cellulosic Pulp Properties of Hybrid Aspen Clones. Forests, 10(5), 450. https://doi.org/10.3390/f10050450