Conversion of Natural Evergreen Broadleaved Forests Decreases Soil Organic Carbon but Increases the Relative Contribution of Microbial Residue in Subtropical China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Soil Sampling, Litterfall, and Fine Root Biomass Measurements
2.3. Soil Analysis
2.4. Determination of Soil Amino Sugars
2.5. Statistics
3. Results
3.1. Soil Properties
3.2. Concentrations of Amino Sugars in the Different Forests
3.3. Amino Sugar Biomarker Ratios
3.4. Ratios of TAS to SOC in the Different Forest Type
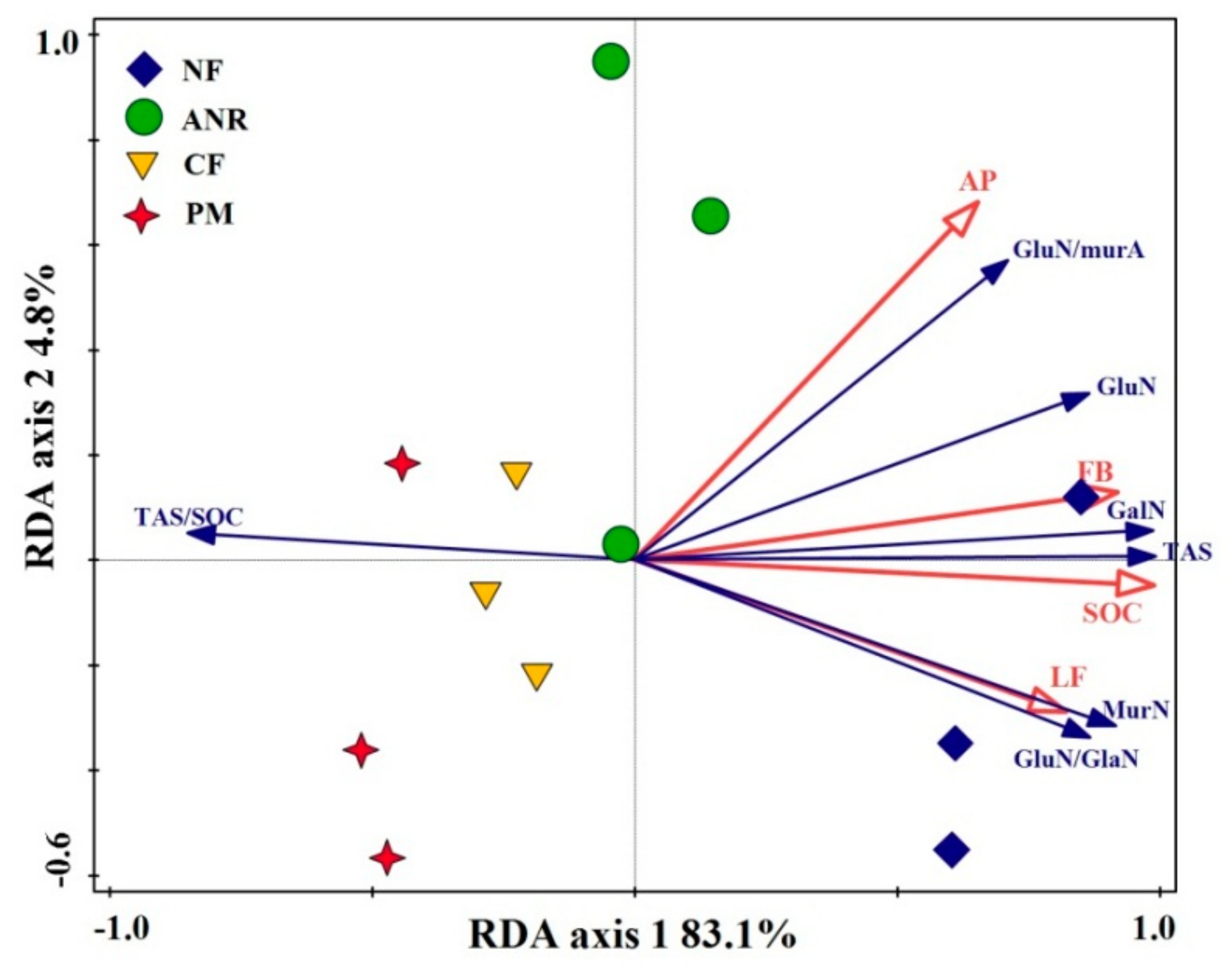
3.5. Correlation between Amino Sugars and Soil Properties
4. Discussion
4.1. Effects of Natural Forest Conversion on Concentrations of Soil Total C and Amino Sugars
4.2. Amino Sugar Biomarker Ratios
4.3. Contribution of TAS to SOC in the Different Forest Types
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Zhu, S.; Wang, Z.; Chen, D.; Dai, G.; Feng, B.; Su, X.; Hu, H.; Li, K.; Han, W.; et al. Divergent accumulation of microbial necromass and plant lignin components in grassland soils. Nat. Commun. 2018, 9, 3480. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Cheng, G.; Wixon, D.L.; Balser, T.C. An Absorbing Markov Chain approach to understanding the microbial role in soil carbon stabilization. Biogeochemistry 2011, 106, 303–309. [Google Scholar] [CrossRef]
- Schimel, J.P.; Schaeffer, S.M. Microbial control over carbon cycling in soil. Front. Microbiol. 2012, 3, 1–11. [Google Scholar] [CrossRef]
- Benner, R. Biosequestration of carbon by heterotrophic microorganisms. Nat. Rev. Microbiol. 2011, 9, 75. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Balser, T.C. Microbial production of recalcitrant organic matter in global soils: Implications for productivity and climate policy. Nat. Rev. Microbiol. 2011, 9, 75. [Google Scholar] [CrossRef]
- Zhang, X.; Amelung, W.; Yuan, Y.; Samson-Liebig, S.; Brown, L.; Zech, W. Land-use effects on amino sugars in particle size fractions of an Argiudoll. Appl. Soil Ecol. 1999, 11, 271–275. [Google Scholar] [CrossRef]
- Liang, C.; Balser, T.C. Warming and nitrogen deposition lessen microbial residue contribution to soil carbon pool. Nat. Commun. 2012, 3, 1222. [Google Scholar] [CrossRef] [Green Version]
- Raich, J.W. Effects of Forest Conversion on the Carbon Budget of a Tropical Soil. Biotropica 1983, 15, 177–184. [Google Scholar]
- Piquer-Rodríguez, M.; Torella, S.; Gavier-Pizarro, G.; Volante, J.; Somma, D.; Ginzburg, R.; Kuemmerle, T. Effects of past and future land conversions on forest connectivity in the Argentine Chaco. Landsc. Ecol. 2015, 30, 817–833. [Google Scholar]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land-use change: processes and potential. Chang. Boil. 2000, 6, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Lal, R. Forest soils and carbon sequestration. For. Ecol. Manag. 2005, 220, 242–258. [Google Scholar] [CrossRef]
- Schimel, J.; Balser, T.C.; Wallenstein, M. Microbial stress-response physiology and its implications for ecosystem function. Ecology 2007, 88, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Schimel, J. Microbial ecology: Linking omics to biogeochemistry. Nat. Microbiol. 2016, 1, 15028. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; Liang, C.; Lynch, L.; Xie, H.; Bao, X. Reforestation accelerates soil organic carbon accumulation: Evidence from microbial biomarkers. Soil Boil. Biochem. 2019, 131, 182–190. [Google Scholar] [CrossRef]
- Liang, C.; Balser, T.C. Preferential sequestration of microbial carbon in subsoils of a glacial-landscape toposequence, Dane County, WI, USA. Geoderma 2008, 148, 113–119. [Google Scholar] [CrossRef]
- Joergensen, R.G. Amino sugars as specific indices for fungal and bacterial residues in soil. Boil. Fertil. Soils 2018, 54, 559–568. [Google Scholar] [CrossRef]
- Amelung, W.; Miltner, A.; Zhang, X.; Zech, W. Fate of microbial residues during litter decomposition as affected by minerals. Soil Sci. 2001, 166, 598–606. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Wichern, F. Quantitative assessment of the fungal contribution to microbial tissue in soil. Soil Boil. Biochem. 2008, 40, 2977–2991. [Google Scholar] [CrossRef]
- Reay, M.K.; Knowles, T.D.J.; Jones, D.L.; Evershed, R.P. Development of Alditol Acetate Derivatives for the Determination of 15N-Enriched Amino Sugars by Gas Chromatography–Combustion–Isotope Ratio Mass Spectrometry. Anal. Chem. 2019, 91, 3397–3404. [Google Scholar] [CrossRef]
- Zhang, X.; Amelung, W. Gas chromatographic determination of muramic acid, glucosamine, mannosamine, and galactosamine in soils. Soil Boil. Biochem. 1996, 28, 1201–1206. [Google Scholar] [CrossRef]
- Amelung, W.; Brodowski, S.; Sandhage-Hofmann, A.; Bol, R. Combining Biomarker with Stable Isotope Analyses for Assessing the Transformation and Turnover of Soil Organic Matter. Adv. Agron. 2008, 100, 155–250. [Google Scholar]
- Parsons, J.W. Chemistry and Distribution of Amino Sugars in Soils and Soil Organisms. Soil Biochem. 1981, 5, 197–227. [Google Scholar]
- Guggenberger, G.; Frey, S.D.; Six, J.; Paustian, K.; Elliott, E.T. Bacterial and Fungal Cell-Wall Residues in Conventional and No-Tillage Agroecosystems. Soil Sci. Soc. J. 1999, 63, 1188–1198. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, H.; Xu, M.; Ma, Z.; Wang, Z.-L. Soil organic carbon stocks and CO2 effluxes of native and exotic pine plantations in subtropical China. CATENA 2015, 128, 167–173. [Google Scholar] [CrossRef]
- De Medeiros, E.V.; Duda, G.P.; Dos Santos, L.A.; de Sousa Lima, J.R.; de Almeida-Cortêz, J.S.; Hammecker, C.; Lardy, L.; Cournac, L. Soil organic carbon, microbial biomass and enzyme activities responses to natural regeneration in a tropical dry region in Northeast Brazil. CATENA 2017, 151, 137–146. [Google Scholar] [CrossRef]
- Yang, Y.-S.; Chen, G.-S.; Guo, J.-F.; Xie, J.-S.; Wang, X.-G. Soil respiration and carbon balance in a subtropical native forest and two managed plantations. Plant Ecol. 2007, 193, 71–84. [Google Scholar] [CrossRef]
- Murdiyarso, D.; Van Noordwijk, M.; Wasrin, U.R.; Tomich, T.P.; Gillison, A.N. Environmental benefits and sustainable land-use options in the Jambi transect, Sumatra. J. Veg. Sci. 2002, 13, 429–438. [Google Scholar] [CrossRef]
- Yang, Y.S.; Guo, J.F.; Chen, G.S.; He, Z.M.; Xie, J.S. Effect of slash burning on nutrient removal and soil fertility in Chinese fir and evergreen broadleaved forests of mid-subtropical China. Pedosphere 2003, 13, 87–96. [Google Scholar]
- Tian, D.; Yan, W.; Chen, X.; Deng, X.; Peng, Y.; Kang, W.; Peng, C. Variation in runoff with age of Chinese fir plantations in Central South China. Hydrol. Process. 2008, 22, 4870–4876. [Google Scholar] [CrossRef]
- Chen, G.; Yang, Y.; Yang, Z.; Xie, J.; Guo, J.; Gao, R.; Yin, Y.; Robinson, D. Accelerated soil carbon turnover under tree plantations limits soil carbon storage. Sci. Rep. 2016, 6, 19693. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Yang, Z.; Xu, C.; Xie, J.; Chen, G.; Lin, C.; Guo, J.; Liu, X.; Xiong, D.; et al. Large Ecosystem service benefits of assisted natural regeneration. J. Geophys. Res. Biogeosci. 2018, 123, 676–687. [Google Scholar] [CrossRef]
- Shono, K.; Cadaweng, E.A.; Durst, P.B. Application of Assisted Natural Regeneration to Restore Degraded Tropical Forestlands. Restor. Ecol. 2007, 15, 620–626. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Guariguata, M.R. Natural regeneration as a tool for large-scale forest restoration in the tropics: prospects and challenges. Biotropica 2016, 48, 716–730. [Google Scholar] [CrossRef]
- Shoo, L.P.; Catterall, C.P. Stimulating Natural Regeneration of Tropical Forest on Degraded Land: Approaches, Outcomes, and Information Gaps. Restor. Ecol. 2013, 21, 670–677. [Google Scholar] [CrossRef]
- Liu, L.; Gundersen, P.; Zhang, T.; Mo, J. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Boil. Biochem. 2012, 44, 31–38. [Google Scholar] [CrossRef]
- Tang, X.L.; Liu, S.G.; Zhou, G.Y.; Zhang, D.Q.; Zhou, C.Y. Soil-atmospheric exchange of CO2, CH4, and N2O in three subtropical forest ecosystems in southern China. Glob. Chang. Biol. 2006, 12, 546–560. [Google Scholar] [CrossRef]
- Wan, X.; Huang, Z.; He, Z.; Yu, Z.; Wang, M.; Davis, M.R.; Yang, Y. Soil C:N ratio is the major determinant of soil microbial community structure in subtropical coniferous and broadleaf forest plantations. Plant Soil 2015, 387, 103–116. [Google Scholar] [CrossRef]
- Odum, E.P. The Strategy of Ecosystem Development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odum, E.P. Trends Expected in Stressed Ecosystems. BioScience 1985, 35, 419–422. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, C.; Luo, Y. Effects of forest degradation on microbial communities and soil carbon cycling: A global meta-analysis. Glob. Ecol. Biogeogr. 2018, 27, 110–124. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, J.; Chen, G.; Yin, Y.; Gao, R.; Lin, C. Effects of forest conversion on soil labile organic carbon fractions and aggregate stability in subtropical China. Plant Soil 2009, 323, 153–162. [Google Scholar] [CrossRef]
- State Soil Survey Service of China (SSSSC). China Soil; China Agricultural Press: Beijing, China, 1998. (In Chinese) [Google Scholar]
- Yang, Y.-S.; Chen, G.-S.; Lin, P.; Xie, J.-S.; Guo, J.-F. Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in Subtropical China. Ann. For. Sci. 2004, 61, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Yang, Z.; Lin, C.; Giardina, C.P.; Xiong, D.; Lin, W.; Chen, S.; Xu, C.; Chen, G.; Xie, J.; et al. Will nitrogen deposition mitigate warming-increased soil respiration in a young subtropical plantation? Agric. Meteorol. 2017, 246, 78–85. [Google Scholar] [CrossRef]
- Sibbesen, E. An investigation of the anion-exchange resin method for soil phosphate extraction. Plant Soil 1978, 50, 305–321. [Google Scholar] [CrossRef]
- Powers, J.S.; Corre, M.D.; Twine, T.E.; Veldkamp, E. Geographic bias of field observations of soil carbon stocks with tropical land-use changes precludes spatial extrapolation. Proc. Natl. Acad. Sci. USA 2011, 108, 6318–6322. [Google Scholar] [CrossRef] [Green Version]
- Luyssaert, S.; Schulze, E.D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213. [Google Scholar] [CrossRef]
- Dybala, K.E.; Matzek, V.; Gardali, T.; Seavy, N.E. Carbon sequestration in riparian forests: A global synthesis and meta-analysis. Glob. Chang. Biol. 2019, 25, 57–67. [Google Scholar] [CrossRef]
- Silver, W.L.; Kueppers, L.M.; Lugo, A.E.; Ostertag, R.; Matzek, V. Carbon sequestration and plant community dynamics following reforestation of tropical pasture. Ecol. Appl. 2004, 14, 1115–1127. [Google Scholar] [CrossRef]
- Sheng, H.; Yang, Y.; Yang, Z.; Chen, G.; Xie, J.; Guo, J.; Zou, S. The dynamic response of soil respiration to land-use changes in subtropical China. Glob. Chang. Boil. 2010, 16, 1107–1121. [Google Scholar] [CrossRef]
- Schulze, E.D.; Wirth, C.; Heimann, M. Climate change: Managing forests after Kyoto. Science 2000, 289, 2058–2059. [Google Scholar] [CrossRef] [PubMed]
- Kiem, R.; Kögel-Knabner, I. Contribution of lignin and polysaccharides to the refractory carbon pool in C-depleted arable soils. Soil Boil. Biochem. 2003, 35, 101–118. [Google Scholar] [CrossRef]
- Glaser, B.; Turrión, M.-B.; Alef, K. Amino sugars and muramic acid—biomarkers for soil microbial community structure analysis. Soil Boil. Biochem. 2004, 36, 399–407. [Google Scholar] [CrossRef]
- Throckmorton, H.M.; Bird, J.A.; Monte, N.; Doane, T.; Firestone, M.K.; Horwath, W.R. The soil matrix increases microbial C stabilization in temperate and tropical forest soils. Biogeochemistry 2015, 122, 35–45. [Google Scholar] [CrossRef]
- Keiluweit, M.; Wanzek, T.; Kleber, M.; Nico, P.; Fendorf, S. Anaerobic microsites have an unaccounted role in soil carbon stabilization. Nat. Commun. 2017, 8, 1771. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.-H.; Li, X.G.; Wang, L.; Mou, X.M.; Kuzyakov, Y. Conversion of Tibetan grasslands to croplands decreases accumulation of microbially synthesized compounds in soil. Soil Boil. Biochem. 2018, 123, 10–20. [Google Scholar] [CrossRef]
- Ye, G.; Lin, Y.; Kuzyakov, Y.; Liu, D.; Luo, J.; Lindsey, S.; Wang, W.; Fan, J.; Ding, W. Manure over crop residues increases soil organic matter but decreases microbial necromass relative contribution in upland Ultisols: Results of a 27-year field experiment. Soil Biol. Biochem. 2019, 134, 15–24. [Google Scholar] [CrossRef]
- Chen, G.-S.; Yang, Y.-S.; Xie, J.-S.; Guo, J.-F.; Gao, R.; Qian, W. Conversion of a natural broad-leafed evergreen forest into pure plantation forests in a subtropical area: Effects on carbon storage. Ann. For. Sci. 2005, 62, 659–668. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, Z.; Wang, F.; Zou, B.; Chen, Y.; Zhao, J.; Mo, Q.; Li, Y.; Li, X.; Xia, H. Effects of nitrogen and phosphorus addition on soil microbial community in a secondary tropical forest of China. Biol. Fertil. Soils 2015, 51, 207–215. [Google Scholar] [CrossRef]
- Turner, B.L.; Brenes-Arguedas, T.; Condit, R. Pervasive phosphorus limitation of tree species but not communities in tropical forests. Nature 2018, 555, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Placella, S.A.; Daydé, F.; Bernard, L.; Robin, A.; Journet, E.-P.; Justes, E.; Hinsinger, P. Phosphorus availability and microbial community in the rhizosphere of intercropped cereal and legume along a P-fertilizer gradient. Plant Soil 2016, 407, 119–134. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Townsend, A.R.; Schmidt, S.K. Phosphorus Limitation of microbial processes in moist tropical forests: Evidence from short-term laboratory incubations and field studies. Ecosystems 2002, 5, 680–691. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Direct nitrogen and phosphorus limitation of arbuscular mycorrhizal fungi: a model and field test. New Phytol. 2002, 155, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.; Fujinuma, R.; Wei, L.; Balser, T.C. Tree species-specific effects on soil microbial residues in an upper Michigan old-growth forest system. Forestry 2007, 80, 65–72. [Google Scholar] [CrossRef]
- Shao, S.; Zhao, Y.; Zhang, W.; Hu, G.; Xie, H.; Yan, J.; Han, S.; He, H.; Zhang, X. Linkage of microbial residue dynamics with soil organic carbon accumulation during subtropical forest succession. Soil Boil. Biochem. 2017, 114, 114–120. [Google Scholar] [CrossRef]
- Guo, J.; Yang, Z.; Lin, C.; Liu, X.; Chen, G.; Yang, Y. Conversion of a natural evergreen broadleaved forest into coniferous plantations in a subtropical area: effects on composition of soil microbial communities and soil respiration. Boil. Fertil. Soils 2016, 52, 799–809. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in microbial community composition through two soil depth profiles. Soil Boil. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Steenwerth, K.L.; Drenovsky, R.E.; Lambert, J.J.; Kluepfel, D.A.; Scow, K.M.; Smart, D.R. Soil morphology, depth and grapevine root frequency influence microbial communities in a Pinot noir vineyard. Soil Boil. Biochem. 2008, 40, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Liang, C.; He, H.; Zhang, X. Variations in Soil Microbial Communities and Residues Along an Altitude Gradient on the Northern Slope of Changbai Mountain, China. PLoS ONE 2013, 8, e66184. [Google Scholar] [CrossRef] [PubMed]
- Sradnick, A.; Oltmanns, M.; Raupp, J.; Joergensen, R.G. Microbial residue indices down the soil profile after long-term addition of farmyard manure and mineral fertilizer to a sandy soil. Geoderma 2014, 226, 79–84. [Google Scholar] [CrossRef]
- Hedlund, K.; Griffiths, B.; Christensen, S.; Scheu, S.; Setälä, H.; Tscharntke, T.; Verhoef, H. Trophic interactions in changing landscapes: responses of soil food webs. Basic Appl. Ecol. 2004, 5, 495–503. [Google Scholar] [CrossRef]
- Moritz, L.K.; Liang, C.; Wagai, R.; Kitayama, K.; Balser, T.C. Vertical distribution and pools of microbial residues in tropical forest soils formed from distinct parent materials. Biogeochemistry 2009, 92, 83–94. [Google Scholar] [CrossRef]
- Turrión, M.B.; Glaser, B.; Zech, W. Effects of deforestation on contents and distribution of amino sugars within particle-size fractions of mountain soils. Boil. Fertil. Soils 2002, 35, 49–53. [Google Scholar] [CrossRef]
- Taylor, J.P.; Wilson, B.; Mills, M.S.; Burns, R.G. Comparison of microbial numbers and enzymatic activities in surface soils and subsoils using various techniques. Soil Boil. Biochem. 2002, 34, 387–401. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the Design of Ecological Field Experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | NF | ANR | CF | PM |
|---|---|---|---|---|
| Altitude (m) | 315 | 315 | 301 | 303 |
| Slope (°) | 35 | 28 | 30 | 35 |
| Canopy coverage (%) | 89 | 90 | 65 | 70 |
| Mean tree height (m) | 11.9 | 10.8 | 18.2 | 18.4 |
| Mean tree diameter at breast height (cm) | 20.0 | 14.3 | 15.6 | 16.3 |
| Stand density (stem ha−1) | 1955 | 3788 | 2858 | 1500 |
| Soil Property | 0–10 cm Layer | 10–20 cm Layer | ||||||
|---|---|---|---|---|---|---|---|---|
| NF | ANR | PM | CF | NF | ANR | PM | CF | |
| SOC (mg g−1) | 51.8 ± 3.0 A | 30.0 ± 2.7 B | 18.6 ± 0.9 C | 20.9 ± 1.3 bC | 27.0 ± 1.4 a | 12.7 ± 0.1 b | 10.7 ± 0.5 b | 11.7 ± 0.7 b |
| TN (mg g−1) | 2.75 ± 0.10 A | 1.91 ± 0.12 B | 1.13 ± 0.09 C | 1.29 ± 0.07 C | 1.47 ± 0.03 a | 1.06 ± 0.03 b | 0.75 ± 0.01 c | 0.86 ± 0.05 bc |
| C/N | 18.8 ± 0.6 A | 15.6 ± 0.4 B | 16.5 ± 1.1 BC | 16.2 ± 0.2 C | 18.3 ± 0.8 a | 12.0 ± 0.3 b | 14.3 ± 0.4 b | 13.7 ± 0.2 b |
| CEC (cmol kg−1) | 15.8 ± 1.7 A | 7.1 ± 1.2 B | 6.7 ± 0.6 B | 6.5 ± 0.5 B | 9.8 ± 0.4 a | 5.5 ± 0.2 b | 5.7 ± 0.3 b | 6.0 ± 0.2 b |
| pH | 4.08 ± 0.07 B | 4.44 ± 0.09 A | 4.47 ± 0.03 A | 4.52 ± 0.06 A | 4.10 ± 0.03 a | 4.28 ± 0.04 ab | 4.39 ± 0.02 b | 4.35 ± 0.02 b |
| NH4+ (mg kg−1) | 27.3 ± 0.8 A | 26.5 ± 4.7 A | 11.6 ± 0.9 B | 12.8 ± 1.0 B | 12.2 ± 0.2 a | 9.6 ± 0.8 b | 6.4 ± 0.1 b | 6.8 ± 0.2 b |
| NO3− (mg kg−1) | 0.86 ± 0.09 BC | 1.98 ± 0.33 A | 0.26 ± 0.03 C | 1.38 ± 0.18 AB | 1.19 ± 0.04 a | 1.14 ± 0.06 a | 0.32 ± 0.03 c | 0.73 ± 0.05 b |
| Sand (2–0.05 mm, %) | 16.4 ± 2.7 A | 15.2 ± 3.1 A | 11.1 ± 0.3 A | 12.3 ± 0.2 A | 16.3 ± 1.1 a | 17.5 ± 3.0 a | 13.3 ± 0.5 b | 16.6 ± 0.5 a |
| Silt (0.05–0.002 mm, %) | 37.3 ± 1.9 A | 34.9 ± 1.9 A | 39.0 ± 1.6 A | 37.5 ± 1.2 A | 40.6 ± 1.8 a | 36.6 ± 1.8 a | 40.8 ± 1.2 a | 41.1 ± 0.2 a |
| Clay (<0.002 mm, %) | 46.3 ± 3.3 A | 50.0 ± 1.9 A | 50.0 ± 1.9 A | 50.3 ± 1.4 A | 43.1 ± 2.8 a | 45.8 ± 1.7 a | 45.8 ± 1.7 a | 42.4 ± 0.6 a |
| Available P (mg kg−1) | 5.47 ± 0.45 A | 6.55 ± 0.56 A | 2.57 ± 0.52 B | 3.46 ± 0.35 B | 1.10 ± 0.14 b | 1.96 ± 0.20 a | 1.17 ± 0.10 b | 1.22 ± 0.06 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Chen, S.; Li, Y.; Wang, Q.; Zhong, X.; Yang, Z.; Lin, C.; Yang, Y. Conversion of Natural Evergreen Broadleaved Forests Decreases Soil Organic Carbon but Increases the Relative Contribution of Microbial Residue in Subtropical China. Forests 2019, 10, 468. https://doi.org/10.3390/f10060468
Yang L, Chen S, Li Y, Wang Q, Zhong X, Yang Z, Lin C, Yang Y. Conversion of Natural Evergreen Broadleaved Forests Decreases Soil Organic Carbon but Increases the Relative Contribution of Microbial Residue in Subtropical China. Forests. 2019; 10(6):468. https://doi.org/10.3390/f10060468
Chicago/Turabian StyleYang, Liuming, Silu Chen, Yan Li, Quancheng Wang, Xiaojian Zhong, Zhijie Yang, Chengfang Lin, and Yusheng Yang. 2019. "Conversion of Natural Evergreen Broadleaved Forests Decreases Soil Organic Carbon but Increases the Relative Contribution of Microbial Residue in Subtropical China" Forests 10, no. 6: 468. https://doi.org/10.3390/f10060468
APA StyleYang, L., Chen, S., Li, Y., Wang, Q., Zhong, X., Yang, Z., Lin, C., & Yang, Y. (2019). Conversion of Natural Evergreen Broadleaved Forests Decreases Soil Organic Carbon but Increases the Relative Contribution of Microbial Residue in Subtropical China. Forests, 10(6), 468. https://doi.org/10.3390/f10060468