
Spatially Explicit Kirtland’s Warbler Habitat Management Scheduling in Michigan’s Upper Peninsula †


Abstract
:1. Introduction
1.1. Kirtland’s Warbler Habitat Management
1.2. Operations Research-Based Methods to Address Spatial and Temporal Complexities
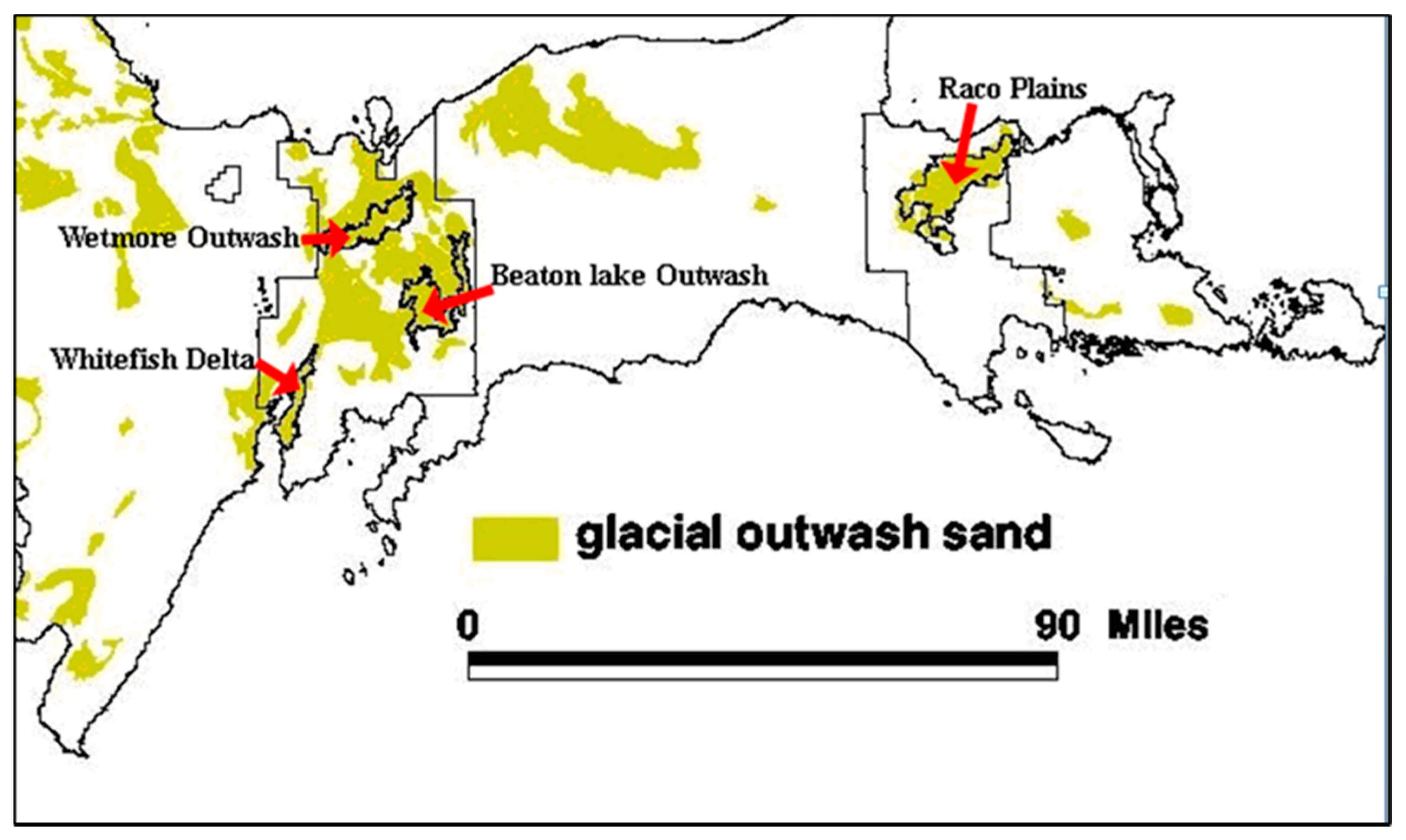
1.3. Study Application Area
1.4. Objectives
- How well does the modeling system used in this study perform in identifying a spatially explicit management strategy for arranging breeding habitat?
- In the context of multiple forest objectives, what are the trade-offs associated with managing for large habitat patches compared to managing habitat without spatial consideration?
- What are the financial trade-offs between alternative spatial management intensities?
2. Materials and Methods
2.1. Operations Research-Based Scheduling Methods for Spatial Arrangement
2.2. Planning Horizon and Time Periods
2.3. Stand Management Options and Costs
2.4. Objective Function, Ending Inventory, and Forest-Wide Constraints
2.5. Model Benchmarks and Scenarios
2.6. Identifying Acceptable Solutions
3. Results
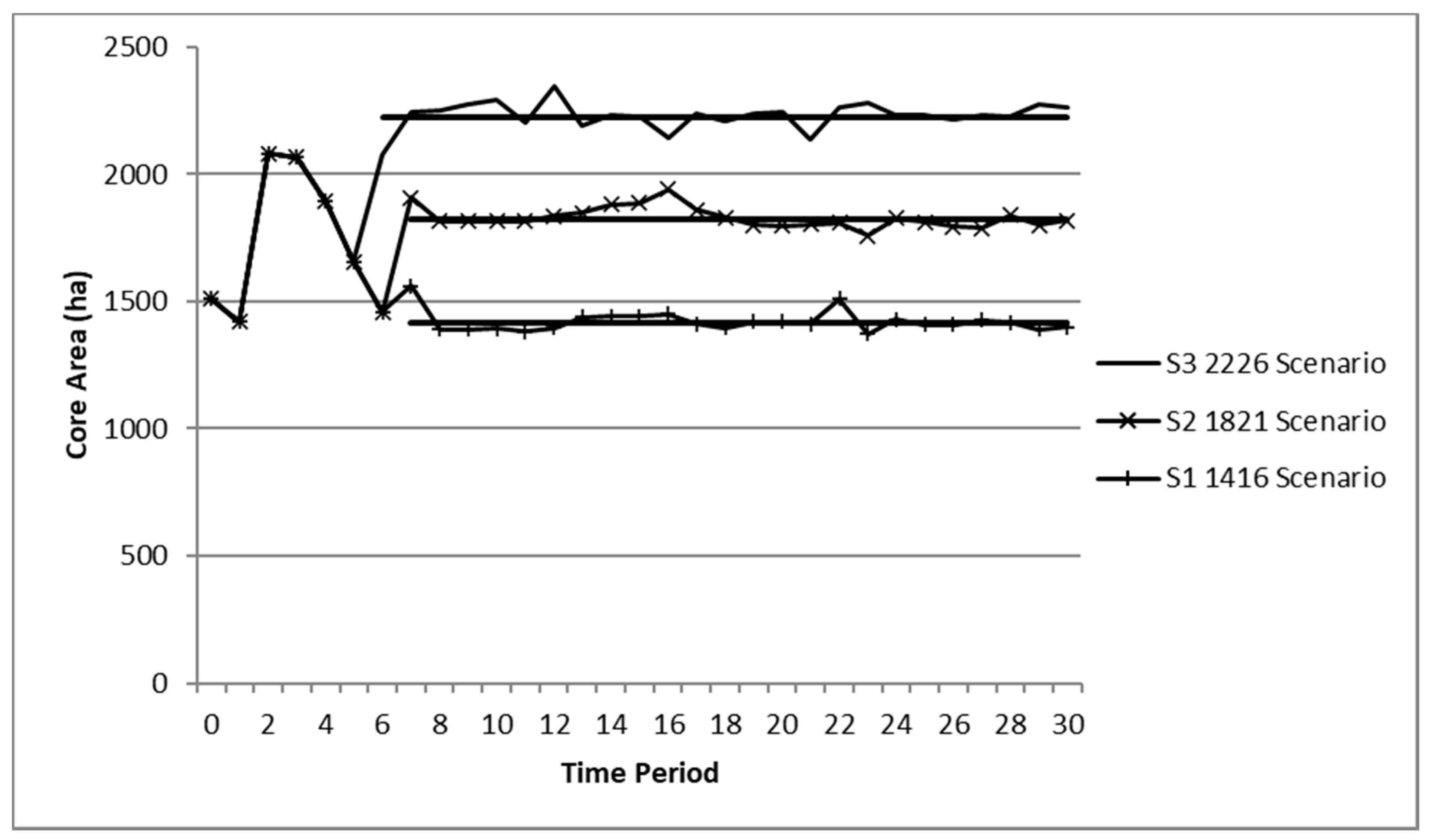
3.1. Kirtland’s Warbler Core Area by Scenario
3.2. Total Kirtland’s Warbler Habitat by Scenario
3.3. Habitat Location and Regeneration
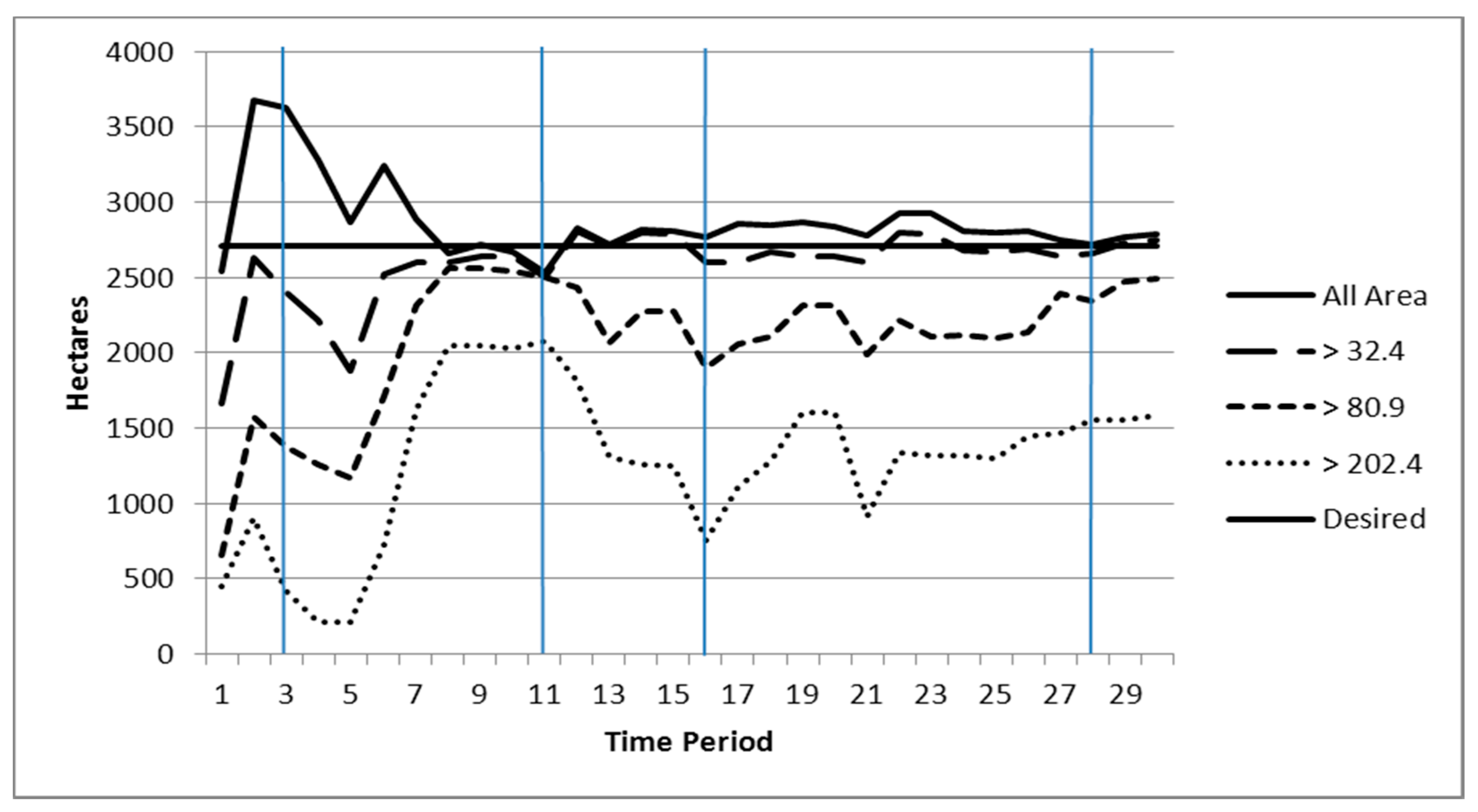
3.4. Patch and Core Area Efficiency Results
3.5. Financial Costs of Kirtland’s Warbler Habitat
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayfield, H. The Kirtland’s Warbler; Cranbrook Institute of Science: Bloomfield Hills, MI, USA, 1960. [Google Scholar]
- US Fish and Wildlife Service. Kirtland’s Warbler (Dendroica kirtlandii) 5 Year Review: Summary and Evaluation; US Fish and Wildlife Service East Lansing Field Office: East Lansing, MI, USA, 2012.
- Byelich, J.; DeCapita, M.E.; Irvine, G.W.; Radtke, R.E.; Johnson, N.I.; Jones, W.R.; Mayfield, H.; Mahalak, W.J. Kirtland’s Warbler Recovery Plan; US Fish and Wildlife Service: Mio, MI, USA, 1976.
- Michigan Department of Natural Resources; US Fish and Wildlife Service; US Forest Service. Kirtland’s Warbler Breeding Range Conservation Plan; Michigan Department of Natural Resources: Lansing, MI, USA, 2015.
- Scott, M.J.; Goble, D.D.; Haines, A.M.; Wiens, J.A.; Neel, M.C. Conservation-reliant species and the future of conservation. Conserv. Lett. 2010, 3, 91–97. [Google Scholar] [CrossRef]
- Brown, D.J.; Ribic, C.A.; Donner, D.M.; Nelson, M.D.; Bocetti, C.I.; Sheffield, C.M.D. Using a full annual cycle model to evaluate long-term population viability of the conservation-reliant Kirtland’s warbler after successful recovery. J. Appl. Ecol. 2016, 54, 439–449. [Google Scholar] [CrossRef]
- US Fish and Wildlife Service. Final Post-Delisting Monitoring Plan for the Kirtland’s Warbler (Setophaga Kirtlandii); Michigan Ecological Services Field Office: East Lansing, MI, USA, 2019.
- Probst, J.R.; Bocetti, C.; Sjogren, S. Population increase in Kirtland’s warbler and summer range expansion to Wisconsin and Michigan’s Upper Peninsula, USA. Oryx 2003, 37, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Richard, T. Confirmed occurrence and nesting of the Kirtland’s warbler at CFB Petawawa, Ontario: A first for Canada. Ont. Bird 2008, 26, 2–15. [Google Scholar]
- Trick, J.A.; Greveles, K.; Ditomasso, D.; Robaidek, J. The first Wisconsin nesting record of Kirtland’s warbler (Dendroica kirtlandii). Passeng. Pigeon 2008, 70, 93–102. [Google Scholar]
- Donner, D.M.; Ribic, C.A.; Probst, J.R. Male Kirtland’s Warblers’ patch-level response to landscape structure during periods of varying population size and habitat amounts. Forest Ecol. Manag. 2009, 258, 1093–1101. [Google Scholar] [CrossRef]
- Probst, J.R. Kirtland’s warbler breeding biology and habitat management. In Integrating Forest Management for Wildlife and Fish; North Central Forest Experiment Station: Minneapolis, MN, USA, 1988. [Google Scholar]
- Kashian, D.M.; Barnes, B.V.; Walker, W.S. Landscape ecosystems of northern Lower Michigan and the occurrence and management of the Kirtland’s warbler. For. Sci. 2003, 20, 140–159. [Google Scholar]
- Probst, J.R.; Weinrich, J. Relating Kirtland’s Warbler population to changing landscape composition and structure. Landsc. Ecol. 1993, 8, 257–271. [Google Scholar] [CrossRef]
- Donner, D.M.; Ribic, C.A.; Probst, J.R. Patch dynamics and the timing of colonization-abandonment events by male Kirtland’s Warblers in an early succession habitat. Biol. Conserv. 2010, 143, 1159–1167. [Google Scholar] [CrossRef]
- Kepler, C.B.; Irvine, G.W.; DeCapita, M.E.; Weinrich, J. The Conservation Management of Kirtland’s Warbler Dendroica kirtlandii. Bird Conserv. Intern. 1996, 6, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Marshall, E.; Haight, R.; Homans, F. Incorporating Environmental Uncertainty into Species Management Decisions: Kirtland’s Warbler Habitat Management as a Case Study. Conserv. Biol. 1998, 76, 975–985. [Google Scholar] [CrossRef]
- Marshall, E.; Homans, F.; Haight, R. Exploring Strategies for Improving the Cost Effectiveness of Endangered Species Management: The Kirtland’s Warbler as a Case Study. Land Econ. 2000, 76, 462–473. [Google Scholar] [CrossRef]
- Donner, D.M.; Probst, J.R.; Ribic, C.A. Influence of habitat amount, arrangement, and use on population trend estimates of male Kirtland’s warblers. Landsc. Ecol. 2008, 23, 467–480. [Google Scholar] [CrossRef]
- Kangas, M.; Kurtilla, T.; Eyvindson, H.; Kangas, J. Decision Support for Forest Management, 2nd ed.; Springer Ingernational Publishing: Berlin, Germany, 2015; Volume 30, pp. 1–307. [Google Scholar]
- Andersson, D.; Eriksson, L.O. Effects of temporal aggregation in integrated strategic/tactical forest planning. For. Policy Econ. 2007, 9, 965–981. [Google Scholar] [CrossRef]
- Rau, V.; Burkhardt, C.; Dorninger, C.; Hjort, L.; Ibe, L.; Kebler, J.; Kristensen, A.; McRobert, W.; Holm, H.S.; Zimmerman, D.; et al. Temporal patterns in ecosystem services research: A review and three recommendations. Ambio 2019, 49, 1377–1393. [Google Scholar] [CrossRef]
- Bellman, R. The theory of dynamic programming. Bull. Am. Math.Soc. 1954, 60, 503–515. [Google Scholar] [CrossRef] [Green Version]
- Hoganson, H.M.; Borges, J.G. Using dynamic programming and overlapping subproblems to address adjacency in large harvest scheduling problems. For. Sci. 1998, 44, 526–538. [Google Scholar]
- Borges, J.; Hoganson, H.; Rose, D. Combining a decomposition strategy with dynamic programming to solve spatially constrained forest management scheduling problems. For. Sci. 1999, 45, 201–212. [Google Scholar]
- Hoganson, H.M.; Bixby, J.; Bergmann, S.; Borges, J.G. Large scale planning to address interior space production studies from northern minnesota. Silva Lusit. 2004, 12, 35–47. [Google Scholar]
- USDA Forest Service. Final Environmental Impact Statement for Forest Plan Revision; Chippewa National Forest Superior National Forest: Duluth, MN, USA, 2004.
- Wei, Y.; Hoganson, H.M. Tests of a Dynamic Programming-Based Heuristic for Scheduling Forest Core Area Production over Large Landscapes. For. Sci. 2008, 54, 367–380. [Google Scholar]
- Henderson, E.B.; Hoganson, H.M. A learning heuristic for integrating spatial and temporal detail in forest planning. Nat. Resour. Model. 2021, 34, e12299. [Google Scholar] [CrossRef]
- Hoganson, H.M.; Rose, D.W. A Simulation Approach for Optimal Timber Management Scheduling. For. Sci. 1984, 30, 220–238. [Google Scholar]
- Hoganson, H.M.; Bixby, J.; Bergmann, S. Scheduling Old Forest Interior Space and Timber Production: Three Large-Scale Test Cases Using the DPSpace Model to Integrate Economic and Ecological Objectives; Minnesota Forest Resources Council: St. Paul, MN, USA, 2003.
- Hoganson, H.M.; Wei, Y.; Hokans, R.H. Integrating spatial objectives into forest plans for Minnesota’s National Forests. In Systems Analysis in Forest Resources: Proceedings of the 2003 Symposium; Pacific Northwest Research Station: Seattle, WA, USA, 2005. [Google Scholar]
- Wei, Y.; Hoganson, H. Landscape impacts from valuing core area in national forest planning. For. Ecol. Manag. 2005, 218, 89–106. [Google Scholar] [CrossRef]
- Hoganson, H.; Borges, J.G.; Wei, Y. Coordinating Management Decisions of Neighboring Stands with Dynamic Programming. In Designing Green Landscapes; Springer: Berlin/Heidelberg, Germany, 2008; pp. 187–213. [Google Scholar]
- USDA Forest Service. Hiawatha National Forest 2006 Forest Plan; USDA Forest Service: Escanaba, MI, USA, 2006.
- Baskent, E.Z.; Jordan, G.A. Characterizing spatial structure of forest landscapes. Can. J. For. Res. 1995, 25, 1830–1849. [Google Scholar] [CrossRef]
- Heinonen, T.; Kurttila, M.; Pukkala, T. Possibilities to aggregate raster cells through spatial optimization in forest planning. Silve Fenn. 2007, 41, 89–103. [Google Scholar] [CrossRef] [Green Version]
- USDA Forest Service. Hiawatha National Forest Final Environmental Impact Statement Appendix I; Forest Service: Escanaba, MI, USA, 2006.
- McGarigal, K.; Marks, B.J. FRAGSTATS: Spatial Pattern Analysis Program for Quantifying Landscape Structure; Gen. Tech. Rep. PNW-GTR-351; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1995; 122p.
- Nelson, M.D.; Buech, R.R. A test of 3 models of Kirtland’s warbler habitat suitability. Wildl. Soc. Bull. 1996, 24, 89–97. [Google Scholar]
- Llorente, D.P.; Hoganson, H.; Campione, M.W.; Miller, S. Using a marginal value approach to integrate ecological and economic objectives across the Minnesota landscape. Forest 2018, 9, 434. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constraint Set | Constraint Type | Constraint Level (ha) | Starting Condition (ha) |
|---|---|---|---|
| Red Pine all ages | Lower | 16,593 | 21,272 |
| Mature red pine | Lower | 11,129 | 11,162 |
| Openings | Lower | 4027 | 3937 |
| Openings | Upper | 4512 | 3937 |
| Regeneration < 10 | Upper | 8033 | 8107 * |
| Non-KW Age 0–2 | Upper | 809 | 1177 * |
| Benchmark/ Scenario | Financial Value of Solution ($MM) | Forest-Wide Constraints | |||
|---|---|---|---|---|---|
| Non-KW Type and Size by Period | Timber Even-Flow | Total KW Age 6–16, 2711 ha by Period | KW Age 6–16, Core Area by Period | ||
| B1: No Habitat | 14.92 | X | X | ||
| B2: Total Habitat | 8.10 | X | X | X | |
| B3: Core Only | 16.33 | X | 2226 | ||
| S1: 1416 Core | 10.34 | X | X | 1416 | |
| S2: 1821 Core | 8.69 | X | X | 1821 | |
| S3: 2226 Core | 6.92 | X | X | 2226 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henderson, E.; Hoganson, H. Spatially Explicit Kirtland’s Warbler Habitat Management Scheduling in Michigan’s Upper Peninsula. Forests 2021, 12, 1065. https://doi.org/10.3390/f12081065
Henderson E, Hoganson H. Spatially Explicit Kirtland’s Warbler Habitat Management Scheduling in Michigan’s Upper Peninsula. Forests. 2021; 12(8):1065. https://doi.org/10.3390/f12081065
Chicago/Turabian StyleHenderson, Eric, and Howard Hoganson. 2021. "Spatially Explicit Kirtland’s Warbler Habitat Management Scheduling in Michigan’s Upper Peninsula" Forests 12, no. 8: 1065. https://doi.org/10.3390/f12081065
APA StyleHenderson, E., & Hoganson, H. (2021). Spatially Explicit Kirtland’s Warbler Habitat Management Scheduling in Michigan’s Upper Peninsula. Forests, 12(8), 1065. https://doi.org/10.3390/f12081065