
Tree-Ring Width Data of Tsuga longibracteata Reveal Growing Season Temperature Signals in the North-Central Pearl River Basin since 1824 AD


Abstract
:1. Introduction
2. Materials and Methods
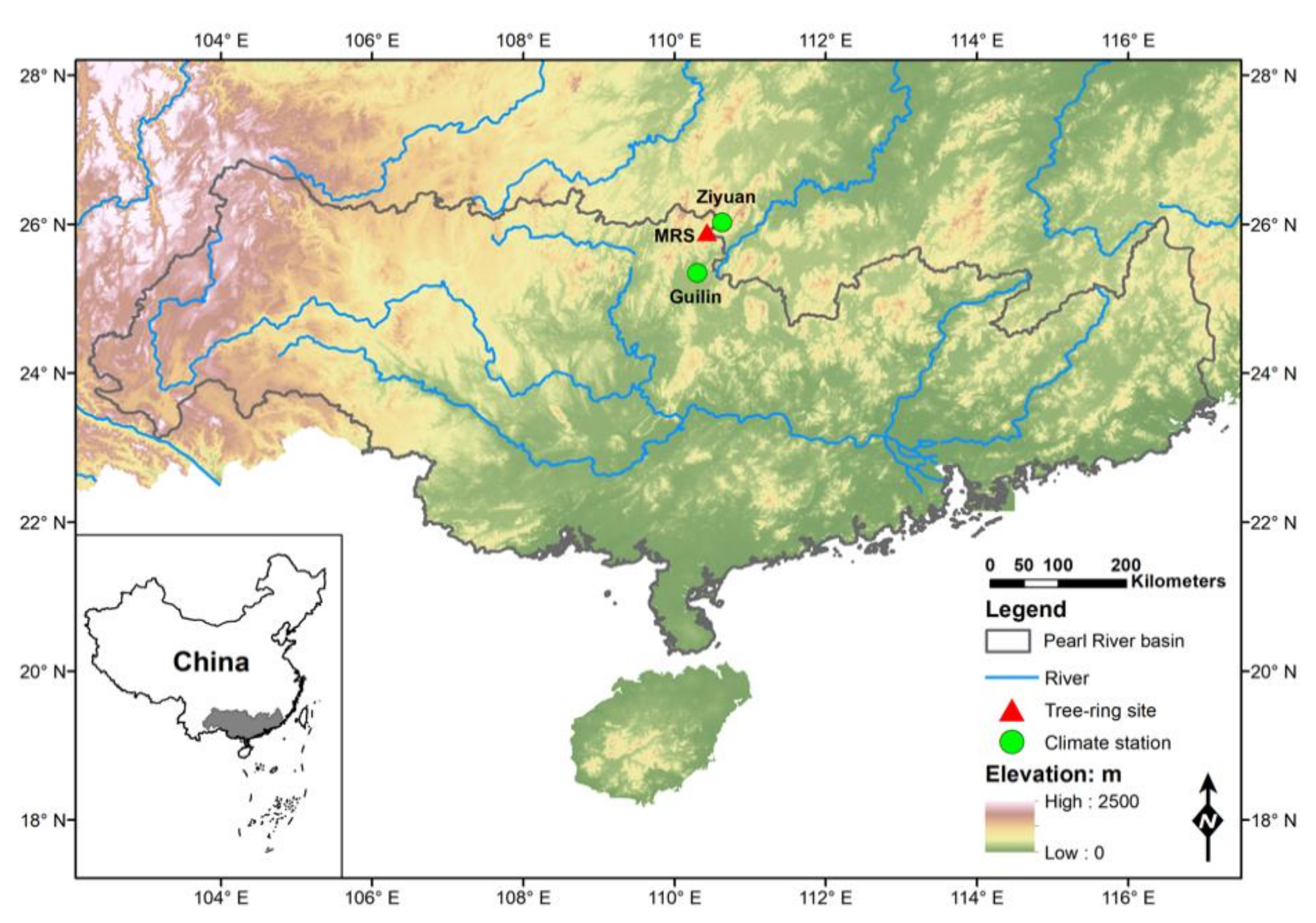
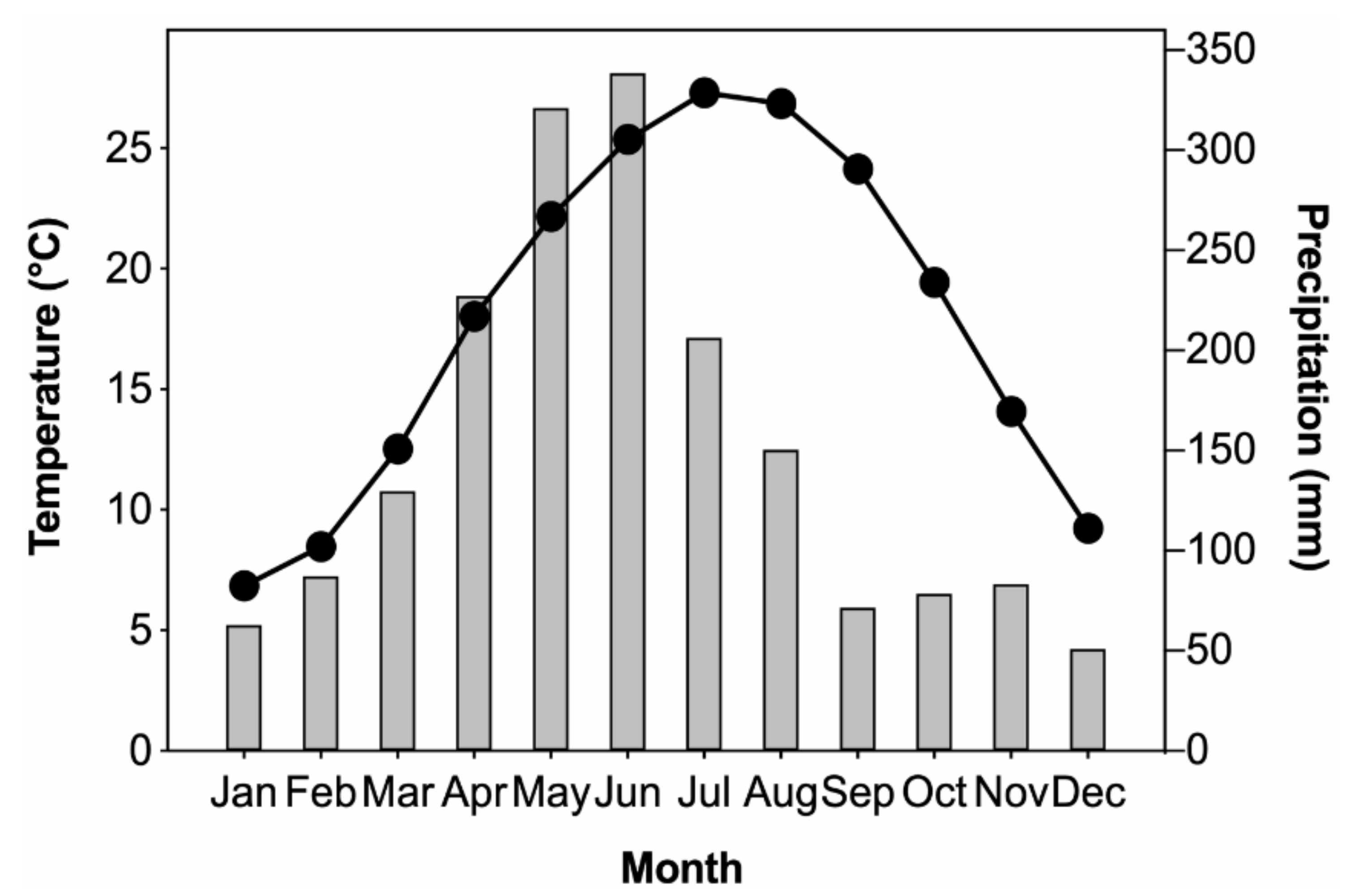
2.1. Characteristics of the Study Area
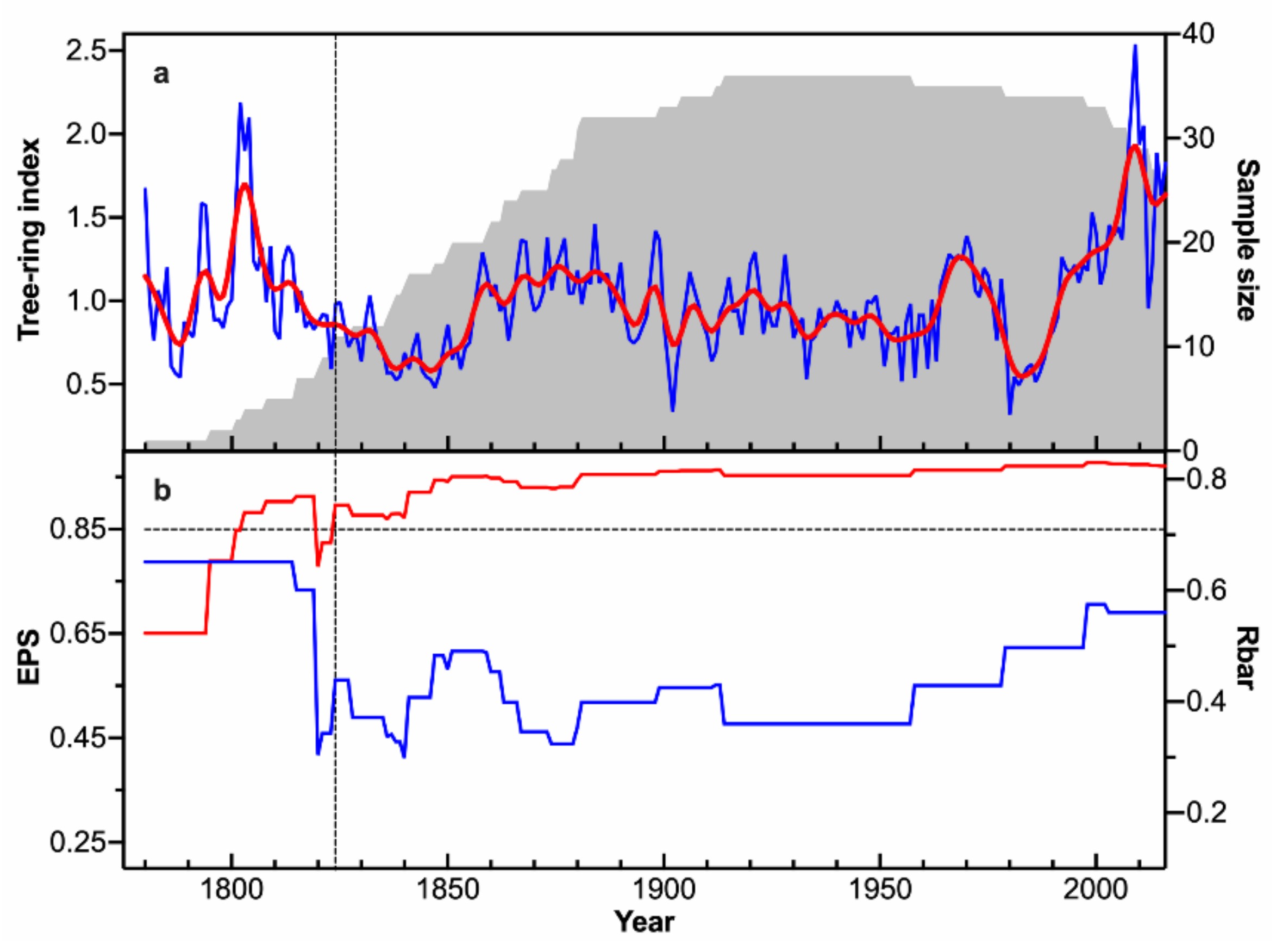
2.2. Tree-Ring Data
2.3. Climate Data
2.4. Methods
3. Results
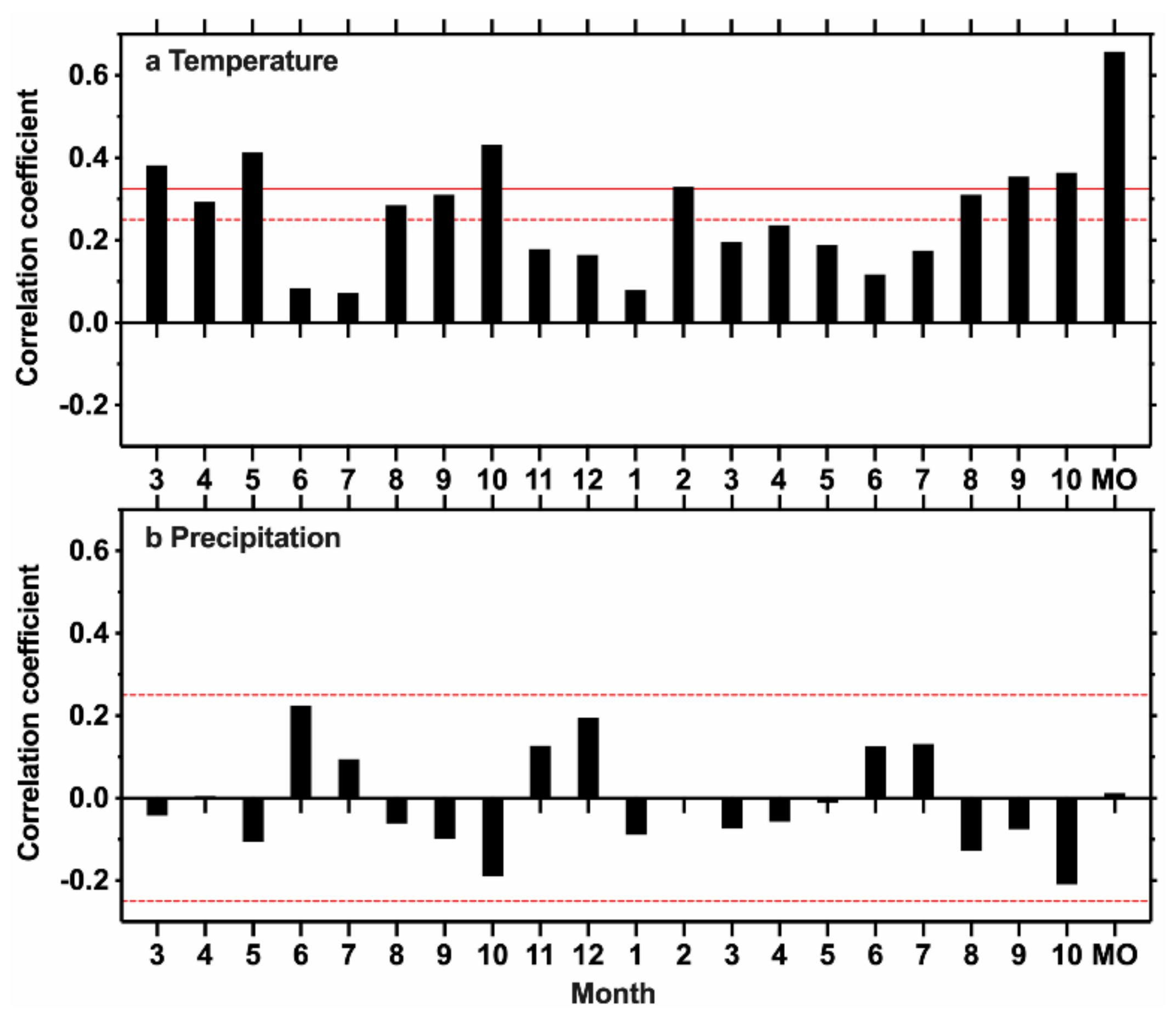
3.1. Climate–Growth Relationship
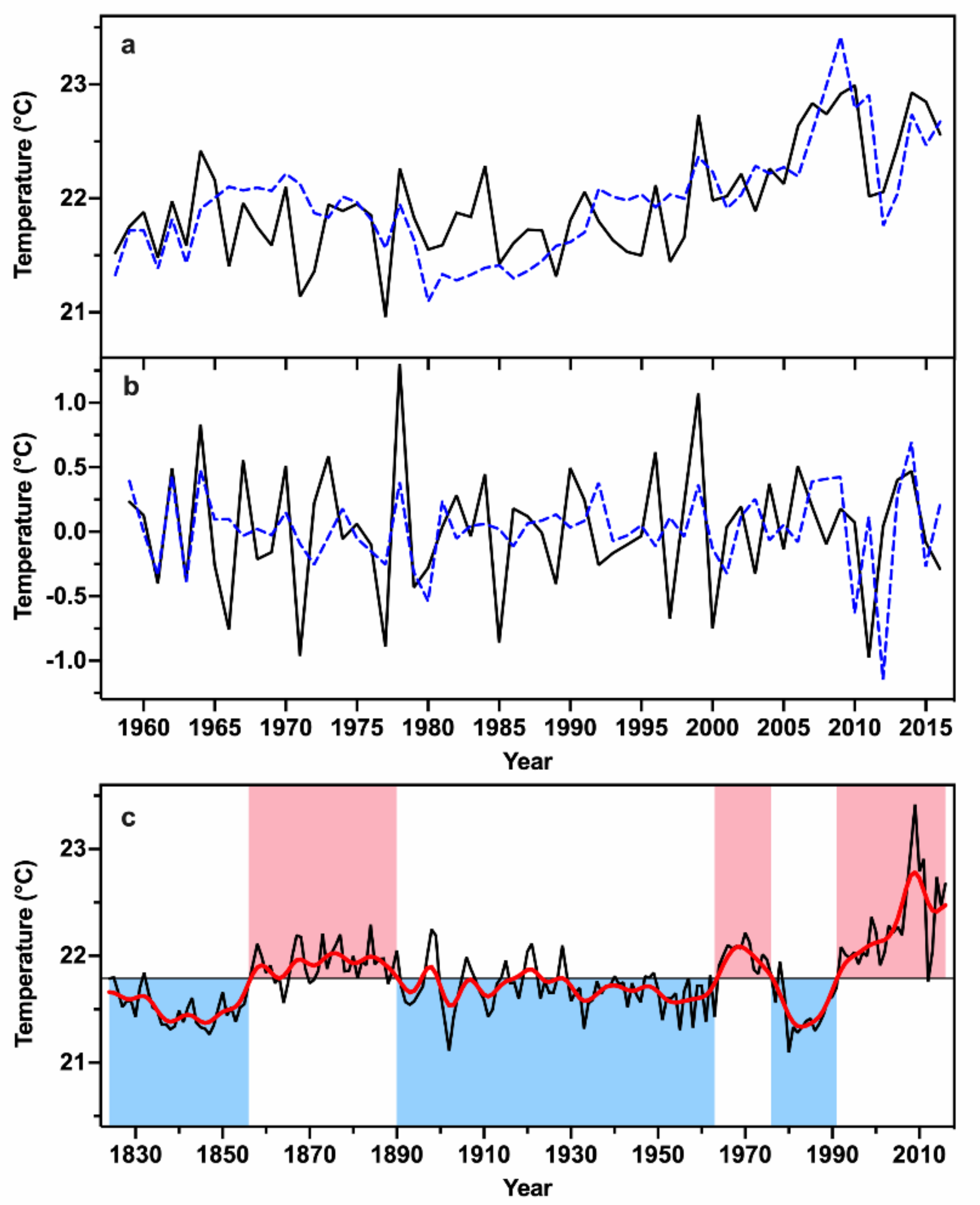
3.2. Previous Growing Season Temperature Reconstruction
4. Discussion
4.1. The Climate–Growth Relationship
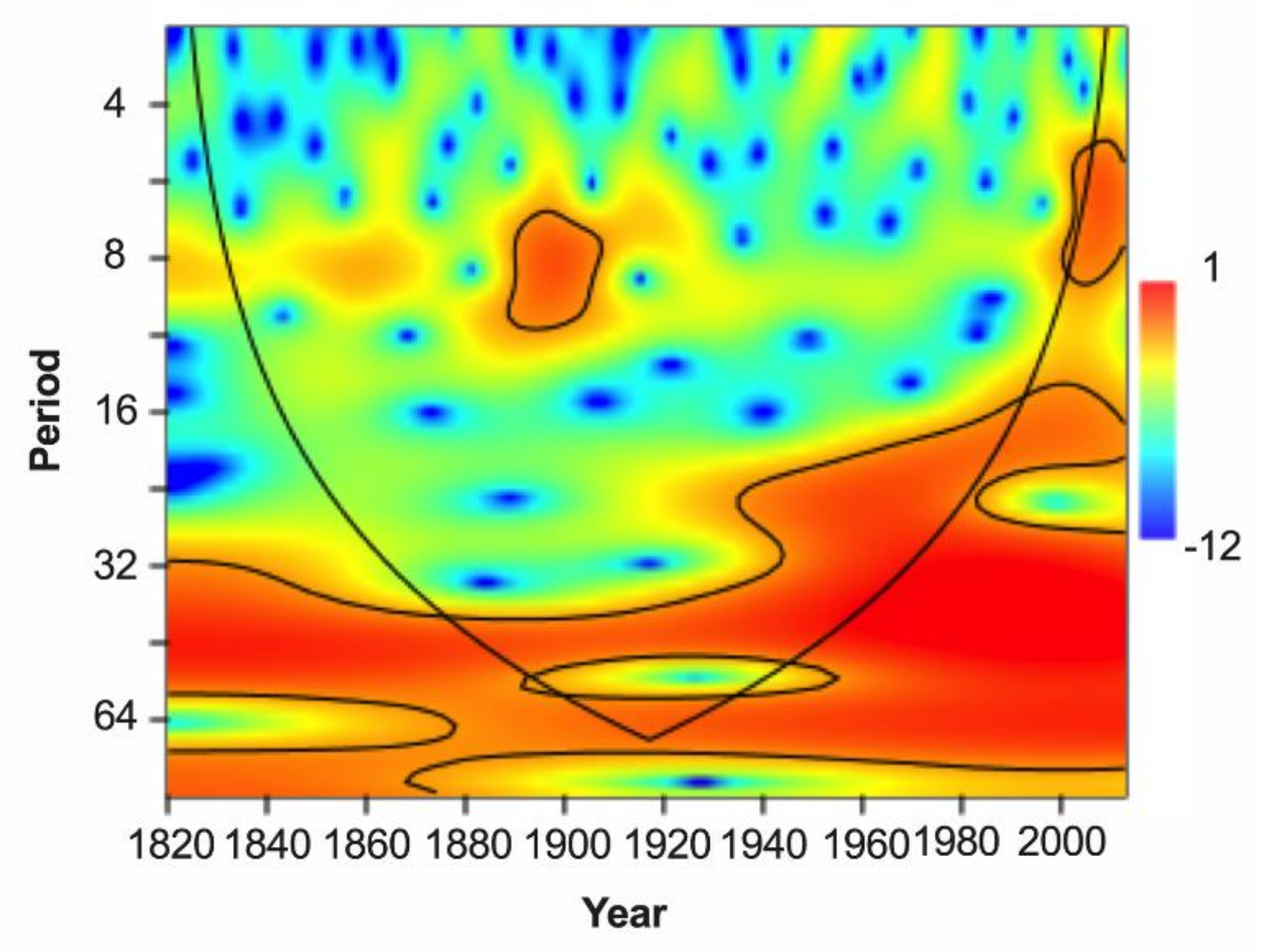
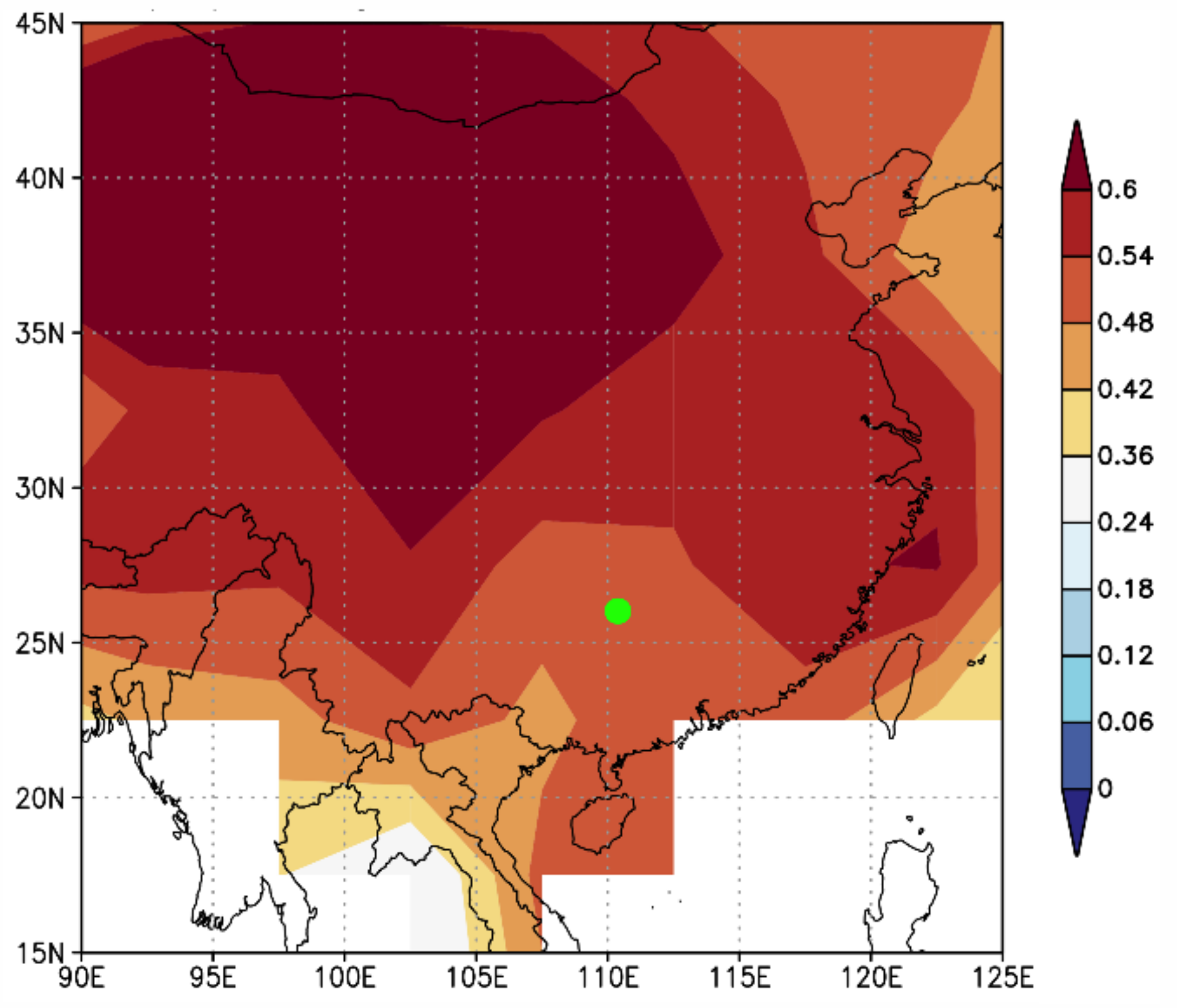
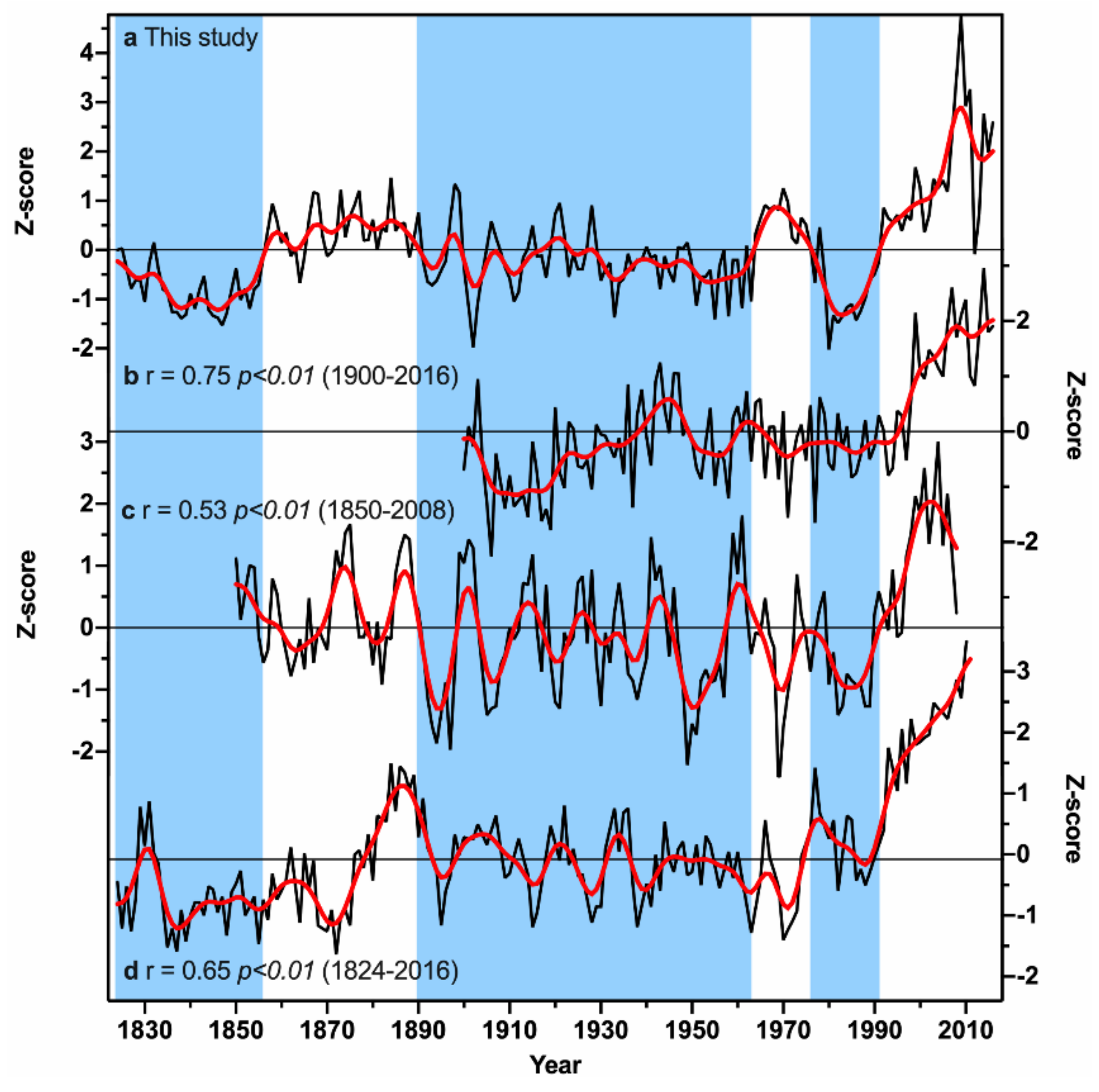
4.2. Tree-Ring Based Growing Season Temperature Variability
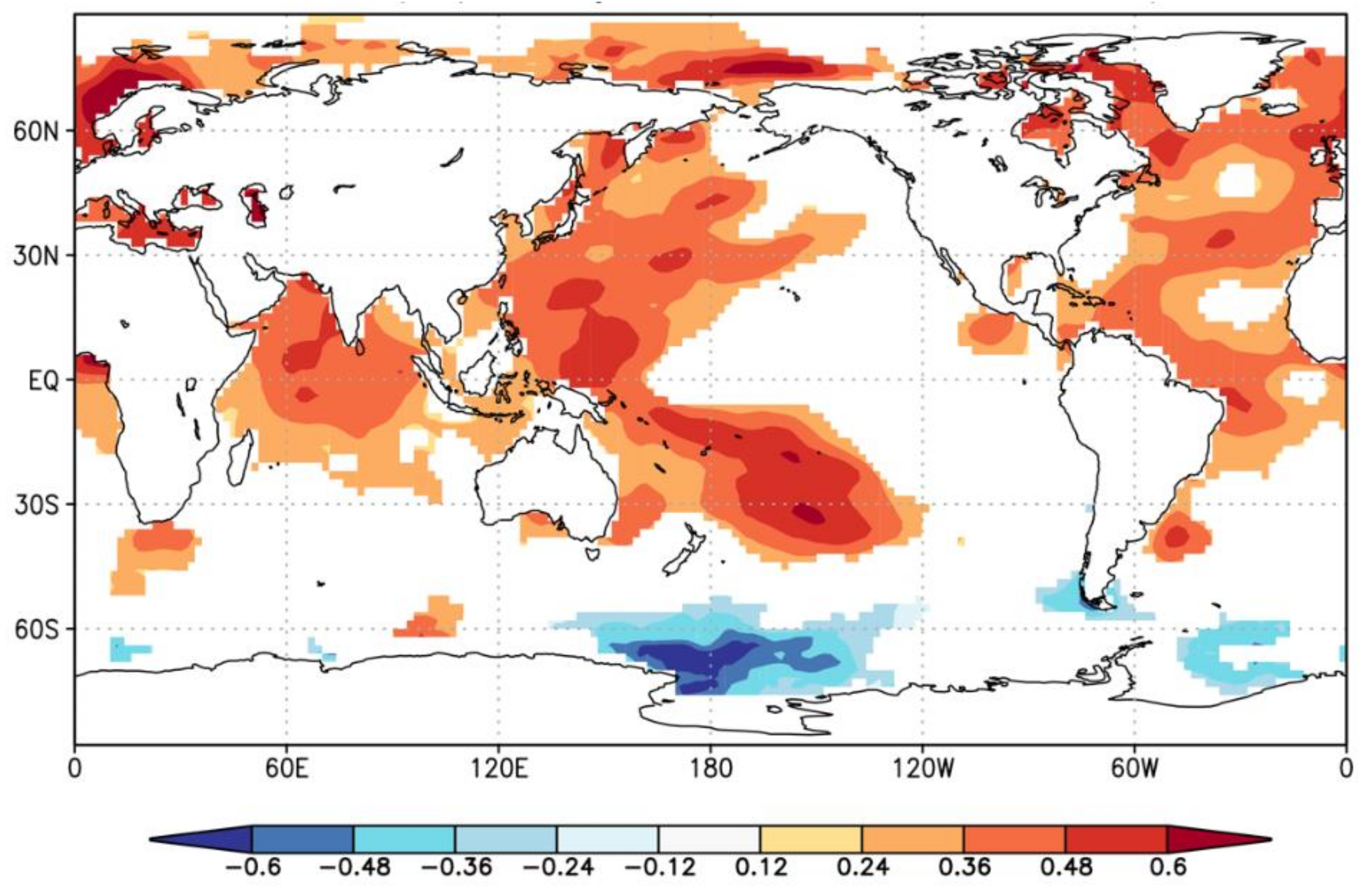
4.3. Linkages of the Growing Season Temperature Variability to Global SSTs
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Harris, N.L.; Gibbs, D.A.; Baccini, A.; Birdsey, R.A.; de Bruin, S.; Farina, M.; Fatoyinbo, L.; Hansen, M.C.; Herold, M.; Houghton, R.A.; et al. Global maps of twenty-first century forest carbon fluxes. Nat. Clim. Chang. 2021, 11, 234–240. [Google Scholar] [CrossRef]
- Jiang, M.; Medlyn, B.E.; Drake, J.E.; Duursma, R.A.; Anderson, I.C.; Barton, C.V.M.; Boer, M.M.; Carrillo, Y.; Castañeda-Gómez, L.; Collins, L.; et al. The fate of carbon in a mature forest under carbon dioxide enrichment. Nature 2020, 580, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Domke, G.M.; Oswalt, S.N.; Walters, B.F.; Morin, R.S. Tree planting has the potential to increase carbon sequestration capacity of forests in the United States. Proc. Natl. Acad. Sci. USA 2020, 117, 24649–24651. [Google Scholar] [CrossRef]
- Begović, K.; Rydval, M.; Mikac, S.; Čupić, S.; Svobodova, K.; Mikoláš, M.; Kozák, D.; Kameniar, O.; Frankovič, M.; Pavlin, J.; et al. Climate-growth relationships of Norway Spruce and silver fir in primary forests of the Croatian Dinaric mountains. Agric. For. Meteorol. 2020, 288–289, 108000. [Google Scholar] [CrossRef]
- Gong, Y.; Staudhammer, C.L.; Wiesner, S.; Starr, G.; Zhang, Y. Characterizing growing season length of subtropical coniferous forests with a phenological model. Forests 2021, 12, 95. [Google Scholar] [CrossRef]
- Poungparn, S.; Komiyama, A.; Umnouysin, S.; Rodtassana, C.; Sangtiean, T.; Maknual, C.; Pravinvongvuthi, T.; Suchewaboripont, V.; Kato, S. Ten-year estimation of net primary productivity in a mangrove forest under a tropical monsoon climate in eastern Thailand: Significance of the temperature Environment in the Dry Season. Forests 2020, 11, 987. [Google Scholar] [CrossRef]
- Au, T.F.; Maxwell, J.; Novick, K.; Robeson, S.; Warner, S.; Lockwood, B.; Phillips, R.; Harley, G.; Telewski, F.; Therrell, M.; et al. Demographic shifts in eastern US forests increase the impact of late-season drought on forest growth. Ecography 2020, 43, 1475–1486. [Google Scholar] [CrossRef]
- Novick, K.; Ficklin, D.; Stoy, P.; Williams, C.; Bohrer, G.; Oishi, A.; Papuga, S.; Blanken, P.; Noormets, A.; Sulman, B.; et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef] [Green Version]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Chang. 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Farahat, E.; Linderholm, H.W. Growth–climate relationship of European beech at its northern distribution limit. Eur. J. Forest Res. 2018, 137, 619–629. [Google Scholar] [CrossRef]
- Depardieu, C.; Girardin, M.; Nadeau, S.; Lenz, P.; Bousquet, J.; Isabel, N. Adaptive genetic variation to drought in a widely distributed conifer suggests a potential for increasing forest resilience in a drying climate. New Phytol. 2020, 227. [Google Scholar] [CrossRef] [Green Version]
- Szymański, N.; Wilczyński, S. Radial growth response of European larch provenances to interannual climate variation in Poland. Forests 2021, 12, 334. [Google Scholar] [CrossRef]
- Di Filippo, A.; Baliva, M.; Brunetti, M.; Fiore, L. Long-term tree-ting tesponse to drought and frost in two Pinus halepensis populations growing under contrasting environmental conditions in peninsular Italy. Forests 2021, 12, 305. [Google Scholar] [CrossRef]
- Šimůnek, V.; Vacek, Z.; Vacek, S.; Ripullone, F.; Hájek, V.; D’Andrea, G. Tree rings of European beech (Fagus Sylvatica L.) indicate the relationship with solar cycles during climate change in central and southern Europe. Forests 2021, 12, 259. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Grau, J.M.; De la Cruz, A.C.; Gil, P.M.; Minaya, M.; Fernández-Cancio, Á. Analysing atmospheric processes and climatic drivers of tree defoliation to determine forest vulnerability to climate warming. Forests 2017, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Chen, D.; Du, J.; Yang, G.; Bai, M.; Zhou, F.; Zheng, Z.; Ruan, C.; Fang, K. A 241-year Cryptomeria fortune tree-ring chronology in humid subtropical China and its linkages with the Pacific Decadal Oscillation. Atmosphere 2020, 11, 247. [Google Scholar] [CrossRef] [Green Version]
- Cabral-Alemán, C.; Pompa-García, M.; Acosta-Hernández, A.C.; Zúñiga-Vásquez, J.M.; Camarero, J.J. Earlywood and latewood widths of Picea chihuahuana show contrasting sensitivity to seasonal climate. Forests 2017, 8, 173. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Shi, J.; Zhang, D.; Yang, B.; Fang, K.; Yue, P.H. Moisture increase in response to high-altitude warming evidenced by tree-rings on the southeastern Tibetan Plateau. Clim. Dyn. 2017, 48, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Deng, Y.; Yan, X.; Li, Q.; Zhang, Y.; Gou, X. The unusual recent streamflow declines in the Bailong River, north-central China, from a multi-century perspective. Quat. Sci. Rev. 2021, 260, 106927. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhang, Y.X.; Shao, X.M. A tree-ring-based drought reconstruction from 1466 to 2013 CE for the Aksu area, western China. Clim. Chang. 2021, 165, 16. [Google Scholar] [CrossRef]
- Jiang, Y.A.; Cao, Y.T.; Han, S.J.; Zhang, J.H.; Hao, L. Spatial variation and temporal instability in the growth/climate relationship of white birch (Betula platyphylla Suk) in the Changbai mountain, China. Forests 2021, 12, 16. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Li, T.; Au, T.F. Tree growth divergence from winter temperature in the Gongga Mountains, southeastern Tibetan Plateau. Asian Geogr. 2020, 37, 1–15. [Google Scholar] [CrossRef]
- Yang, B.; Qin, C.; Bräuning, A.; Osborn, T.J.; Trouet, V.; Ljungqvist, F.C.; Esper, J.; Schneider, L.; Grießinger, J.; Büntgen, U.; et al. Long-term decrease in Asian monsoon rainfall and abrupt climate change events over the past 6,700 years. Proc. Natl. Acad. Sci. USA 2021, 118, e2102007118. [Google Scholar] [CrossRef]
- Zhao, S.; Pederson, N.; D’Orangeville, L.; HilleRisLambers, J.; Boose, E.; Penone, C.; Bauer, B.; Jiang, Y.; Manzanedo, R. The International Tree-Ring Data Bank (ITRDB) revisited: Data availability and global ecological representativity. J. Biogeogr. 2018, 46, 355–368. [Google Scholar] [CrossRef]
- Li, T.; Li, J.; Au, T.F.; Zhang, D.D. Moisture variability in the east Pearl River basin since 1894 CE inferred from tree ring records. Atmosphere 2020, 11, 13. [Google Scholar] [CrossRef]
- Cai, Q.; Liu, Y.; Liu, H.; Sun, C.; Wang, Y. Growing-season precipitation since 1872 in the coastal area of subtropical southeast China reconstructed from tree rings and its relationship with the East Asian summer monsoon system. Ecol. Indic. 2017, 82, 441–450. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.; Wei, W.; Yu, S.; Wang, H. Tree-ring response of subtropical tree species in southeast China on regional climate and sea-surface temperature variations. Trees 2015, 29, 17–24. [Google Scholar] [CrossRef]
- Lin, J.; Hu, Y.; Wang, F. Wood and Bark Anatomy of Nothotsuga (Pinaceae). Ann. Mo. Bot. Gard. 1995, 82, 603–609. [Google Scholar] [CrossRef]
- Tan, X.; Zhang, L.; Zhang, A.; Wang, Y.; Huang, D.; Wu, X.; Sun, X.; Xiong, Q.; Pan, K. The suitable distribution area of Tsuga longibracteata revealed by a climate and spatial constraint model under future climate change scenarios. Acta Ecol. Sin. 2018, 38, 8934–8945. (In Chinese) [Google Scholar] [CrossRef]
- Zhu, X.; Feng, D. Natural regeneration of Tsuga longibracteata Ⅱ. Seedling establishment of different type communities and environmental impacts. J. Fujian Coll. For. 2011, 31, 315–319. (In Chinese) [Google Scholar] [CrossRef]
- Zhong, X.; Huang, H. A study on the water conservation function of Tsuga longibracteata nature forest. J. Fujian For. Sci. Technol. 2004, 31, 54–57. (In Chinese) [Google Scholar] [CrossRef]
- Tian, Q.; Yang, S. Regional climatic response to global warming: Trends in temperature and precipitation in the Yellow, Yangtze and Pearl River basins since the 1950s. Quat. Int. 2017, 440, 1–11. [Google Scholar] [CrossRef]
- Ren, G.; Guan, Z.; Shao, X.; Gong, D. Changes in climatic extremes over mainland China. Clim. Res. 2011, 50, 105–111. [Google Scholar] [CrossRef]
- Wang, Z.; Zhong, R.; Lai, C.; Zeng, Z.; Lian, Y.; Bai, X. Climate change enhances the severity and variability of drought in the Pearl River Basin in South China in the 21st century. Agric. For. Meteorol. 2018, 249, 149–162. [Google Scholar] [CrossRef]
- Huang, C.; Luo, Y.; Zhang, J.; Jiang, D.; Yu, W.; Li, B. The Vertical Distribution Characters of Forest Soil Chemical Properties in Guangxi Maoer Mountain Nature Reserve. Chin. J. Anhui Agric. Sci. 2009, 37, 245–247. (In Chinese) [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Fritts, H.C. Tree rings and climate; Academic Press: London, UK, 1976. [Google Scholar]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Springer Science & Business Media: Dordrecht, The Netherlands, 1990. [Google Scholar]
- Melvin, T.M.; Briffa, K.R.; Nicolussi, K.; Grabner, M. Time-varying-response smoothing. Dendrochronologia 2007, 25, 65–69. [Google Scholar] [CrossRef]
- Cook, E.R.; Peters, K. Calculating unbiased tree-ring indices for the study of climatic and environmental change. Holocene 1997, 7, 361–370. [Google Scholar] [CrossRef]
- Frank, D.; Esper, J.; Cook, E.R. Adjustment for proxy number and coherence in a large-scale temperature reconstruction. Geophys. Res. Lett. 2007, 34, L16709. [Google Scholar] [CrossRef] [Green Version]
- Osborn, T.; Briffa, K.; Jones, P. Adjusting variance for sample size in tree-ring chronologies and other regional mean timeseries. Dendrochronologia 1997, 15, 89–99. [Google Scholar]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteor. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Zhu, Q.; He, H.; Zhou, X.; Tang, Y. Analysis on the characteristics and climatic causes of the persistent low temperature and snow freezing process in Guangxi. Chin. J. Meteorol. Res. Appl. 2019, 40, 38–41. (In Chinese) [Google Scholar] [CrossRef]
- Köppen, W. Das Geographische System der Klimate, in Handbuch der Klimatologie; Borntraeger Science Publishers: Berlin, Germany, 1936. [Google Scholar]
- Peel, M.; Finlayson, B.; McMahon, T. Updated World Map of the Koppen-Geiger Climate Classification. Hydrol. Earth Syst. Sci. Discuss. 2007, 4. [Google Scholar] [CrossRef] [Green Version]
- Michaelsen, J. Cross-validation in statistical climate forecast models. J. Clim. Appl. Meteor. 1987, 26, 1589–1600. [Google Scholar] [CrossRef] [Green Version]
- Torrence, C.; Compo, G.P. A practical guide to wavelet analysis. Bull. Am. Meteorol. Soc. 1998, 79, 61–78. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Thorne, P.W.; Banzon, V.F.; Boyer, T.; Chepurin, G.; Lawrimore, J.H.; Menne, M.J.; Smith, T.M.; Vose, R.S.; Zhang, H.-M. Extended reconstructed sea surface temperature, version 5 (ERSSTv5): Upgrades, validations, and intercomparisons. J. Clim. 2017, 30, 8179–8205. [Google Scholar] [CrossRef]
- Menne, M.J.; Williams, C.N.; Gleason, B.E.; Rennie, J.J.; Lawrimore, J.H. The global historical climatology network monthly temperature dataset, version 4. J. Clim. 2018, 31, 9835–9854. [Google Scholar] [CrossRef]
- Begum, S.; Kudo, K.; Rahman, M.H.; Nakaba, S.; Yamagishi, Y.; Nabeshima, E.; Nugroho, W.D.; Oribe, Y.; Kitin, P.; Jin, H.-O.; et al. Climate change and the regulation of wood formation in trees by temperature. Trees 2018, 32, 3–15. [Google Scholar] [CrossRef]
- Saderi, S.; Rathgeber, C.B.K.; Rozenberg, P.; Fournier, M. Phenology of wood formation in larch (Larix decidua Mill.) trees growing along a 1000-m elevation gradient in the French Southern Alps. Ann. For. Sci. 2019, 76, 89. [Google Scholar] [CrossRef]
- Meng, F.; Zhang, L.; Zhang, Z.; Jiang, L.; Wang, Y.; Duan, J.; Wang, Q.; Li, B.; Liu, P.; Hong, H.; et al. Enhanced spring temperature sensitivity of carbon emission links to earlier phenology. Sci. Total Environ. 2020, 745, 140999. [Google Scholar] [CrossRef]
- Yan, T.; Song, H.; Zeng, H. Spring phenophases of larch are more sensitive to spring warming than to year-round warming: Results of a seasonally asymmetric warming experiment. For. Ecol. Manag. 2020, 474, 118368. [Google Scholar] [CrossRef]
- Zhao, Y.; Shi, J.; Shi, S.; Yu, J.; Lu, H. Tree-ring latewood width based July–August SPEI reconstruction in South China since 1888 and its possible connection with ENSO. J. Meteor. Res. 2017, 31, 39–48. [Google Scholar] [CrossRef]
- Salzer, M.W.; Hughes, M.K.; Bunn, A.G.; Kipfmueller, K.F. Recent unprecedented tree-ring growth in bristlecone pine at the highest elevations and possible causes. Proc. Natl. Acad. Sci. USA 2009, 106, 20348–20353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulamsuren, C.; Hauck, M.; Kopp, G.; Ruff, M.; Leuschner, C. European beech responds to climate change with growth decline at lower, and growth increase at higher elevations in the center of its distribution range (SW Germany). Trees 2017, 31, 673. [Google Scholar] [CrossRef]
- Kolar, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal changes in the climate sensitivity of Norway spruce and European beech along an elevation gradient in Central Europe. Agric. For. Meteorol. 2017, 239, 24–33. [Google Scholar] [CrossRef]
- Classen, A.; Sundqvist, M.; Henning, J.; Newman, G.; Moore, J.; Cregger, M.; Moorhead, L.; Patterson, C. Direct and indirect effects of climate change on soil microbial and soil microbial-plant interactions: What lies ahead? Ecosphere 2015, 6, 130. [Google Scholar] [CrossRef]
- Zhao, Y.; Shi, J.; Shi, S.; Wang, B.; Yu, J. Summer climate implications of tree-ring latewood width: A case study of Tsuga longibracteata in South China. Asian Geogr. 2017, 34, 131–146. [Google Scholar] [CrossRef]
- Čater, M. Microsites influence the light response of young Douglas-fir (Pseudotsuga menziesii (Mirb.) Franco). Forests 2021, 12, 687. [Google Scholar] [CrossRef]
- Zheng, J.; Hua, Z.; Liu, Y.; Hao, Z. Temperature changes derived from phenological and natural evidence in South Central China from 1850 to 2008. Clim. Past 2015, 11, 1553–1561. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Shao, X.; Lu, F.; Li, Y. February–May temperature reconstruction based on tree-ring widths of Abies fargesii from the Shennongjia area in central China. Int. J. Biometeorol. 2016, 60, 1175–1181. [Google Scholar] [CrossRef]
- Hu, K.; Huang, G.; Huang, R. The impact of tropical Indian Ocean variability on summer surface air temperature in China. J. Clim. 2011, 24, 5365–5377. [Google Scholar] [CrossRef]
- Si, D.; Ding, Y. Oceanic forcings of the interdecadal variability in East Asian summer rainfall. J. Clim. 2016, 29, 7633–7649. [Google Scholar] [CrossRef]
- Gray, S.T.; Graumlich, L.J.; Betancourt, J.L.; Pederson, G.T. A tree-ring based reconstruction of the Atlantic Multidecadal Oscillation since 1567 AD. Geophys. Res. Lett. 2004, 31, L12205. [Google Scholar] [CrossRef]
- Schmitt, D.; Gischler, E.; Birgel, D.; Peckmann, J.; Anselmetti, F.S.; Vogel, H. Great Blue Hole (Lighthouse Reef, Belize): A continuous, annually-resolved record of Common Era sea surface temperature, Atlantic Multidecadal Oscillation and cyclone-controlled run-off. Quat. Sci. Rev. 2020, 247, 106570. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Li, T.; Au, T.F. 351-year tree ring reconstruction of the Gongga Mountains winter minimum temperature and its relationship with the Atlantic Multidecadal Oscillation. Clim. Chang. 2021, 165, 49. [Google Scholar] [CrossRef]
- Li, T.; Li, J. A 564-year annual minimum temperature reconstruction for the east central Tibetan Plateau from tree rings. Glob. Planet. Chang. 2017, 157, 165–173. [Google Scholar] [CrossRef]
- Wang, J.; Yang, B.; Ljungqvist, F.C.; Zhao, Y. The relationship between the Atlantic Multidecadal Oscillation and temperature variability in China during the last millennium. J. Quat. Sci. 2013, 28, 653–658. [Google Scholar] [CrossRef]
- Gaire, N.P.; Fan, Z.-X.; Shah, S.K.; Thapa, U.K.; Rokaya, M.B. Tree-ring record of winter temperature from Humla, Karnali, in central Himalaya: A 229 years-long perspective for recent warming trend. Geogr. Ann. Ser. A Phys. Geogr. 2020, 102, 297–316. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site Code | Latitude, Longitude | Elevation (m) | Cores/ Trees | Time Span (AD) | SD | MS | AC1 | Rbar | EPS |
|---|---|---|---|---|---|---|---|---|---|
| MRS | 25.90° N 110.43° E | 1991 | 36/18 | 1780–2016 | 0.34 | 0.18 | 0.65 | 0.46 | 0.92 |
| Rank | Warm Year | Temperature (°C) | Cold Year | Temperature (°C) |
|---|---|---|---|---|
| 1 | 2009 | 23.42 | 1980 | 21.10 |
| 2 | 2008 | 22.99 | 1902 | 21.11 |
| 3 | 2011 | 22.91 | 1847 | 21.27 |
| 4 | 2010 | 22.79 | 1982 | 21.28 |
| 5 | 2014 | 22.74 | 1986 | 21.30 |
| 6 | 2016 | 22.67 | 1955 | 21.31 |
| 7 | 2007 | 22.58 | 1838 | 21.31 |
| 8 | 2015 | 22.47 | 1846 | 21.32 |
| 9 | 1999 | 22.36 | 1933 | 21.32 |
| 10 | 1884 | 22.29 | 1958 | 21.33 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Li, J.; Au, T.F.; Zhang, D.D. Tree-Ring Width Data of Tsuga longibracteata Reveal Growing Season Temperature Signals in the North-Central Pearl River Basin since 1824 AD. Forests 2021, 12, 1067. https://doi.org/10.3390/f12081067
Li T, Li J, Au TF, Zhang DD. Tree-Ring Width Data of Tsuga longibracteata Reveal Growing Season Temperature Signals in the North-Central Pearl River Basin since 1824 AD. Forests. 2021; 12(8):1067. https://doi.org/10.3390/f12081067
Chicago/Turabian StyleLi, Teng, Jinbao Li, Tsun Fung Au, and David Dian Zhang. 2021. "Tree-Ring Width Data of Tsuga longibracteata Reveal Growing Season Temperature Signals in the North-Central Pearl River Basin since 1824 AD" Forests 12, no. 8: 1067. https://doi.org/10.3390/f12081067
APA StyleLi, T., Li, J., Au, T. F., & Zhang, D. D. (2021). Tree-Ring Width Data of Tsuga longibracteata Reveal Growing Season Temperature Signals in the North-Central Pearl River Basin since 1824 AD. Forests, 12(8), 1067. https://doi.org/10.3390/f12081067