


Characterizing Global Patterns of Mangrove Canopy Height and Aboveground Biomass Derived from SRTM Data



Abstract
:1. Introduction
2. Materials and Methods
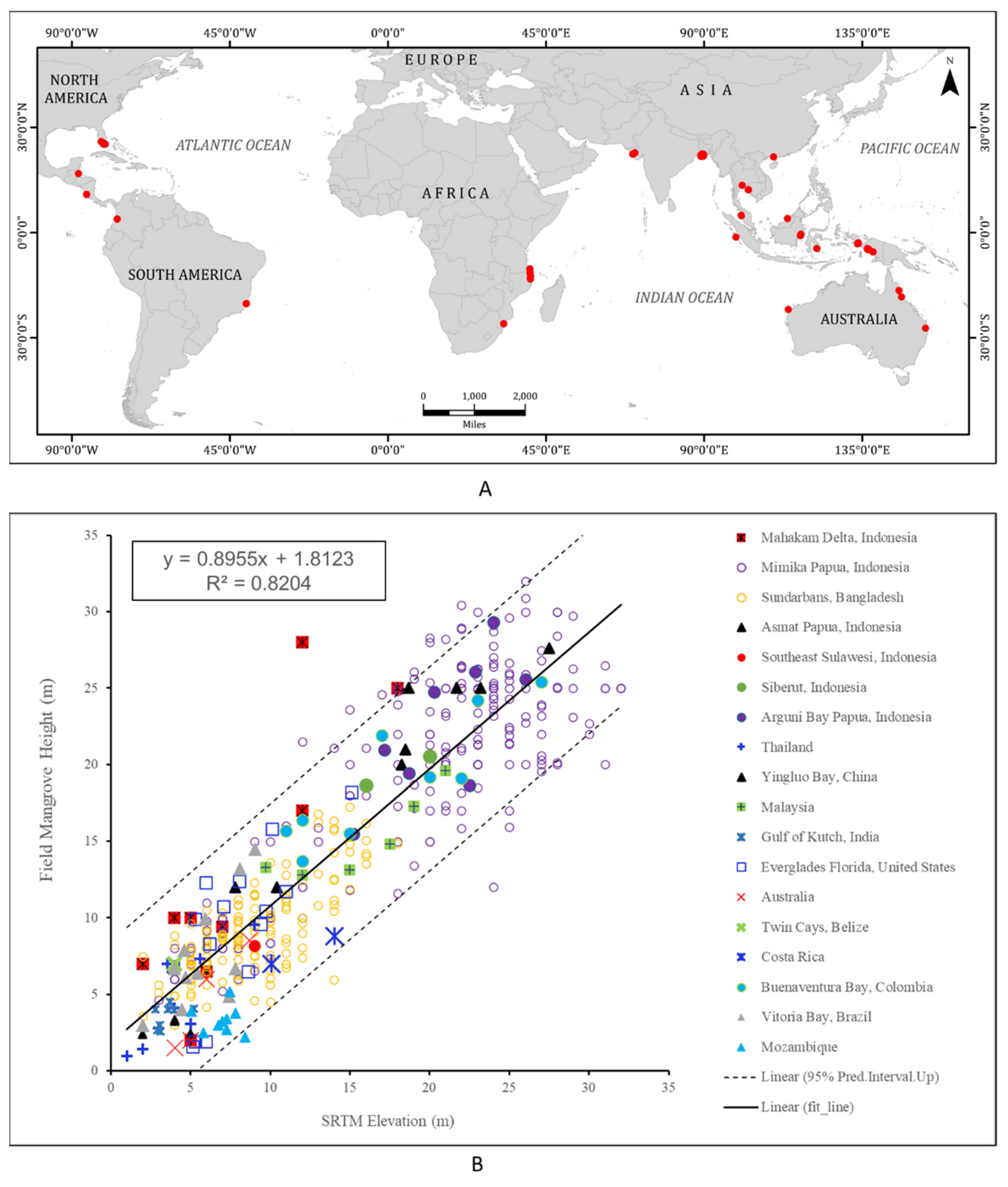
2.1. Field Data and SRTM Height
2.2. Empirical Models
3. Results
3.1. Mangrove Heights and Vertical Accuracy
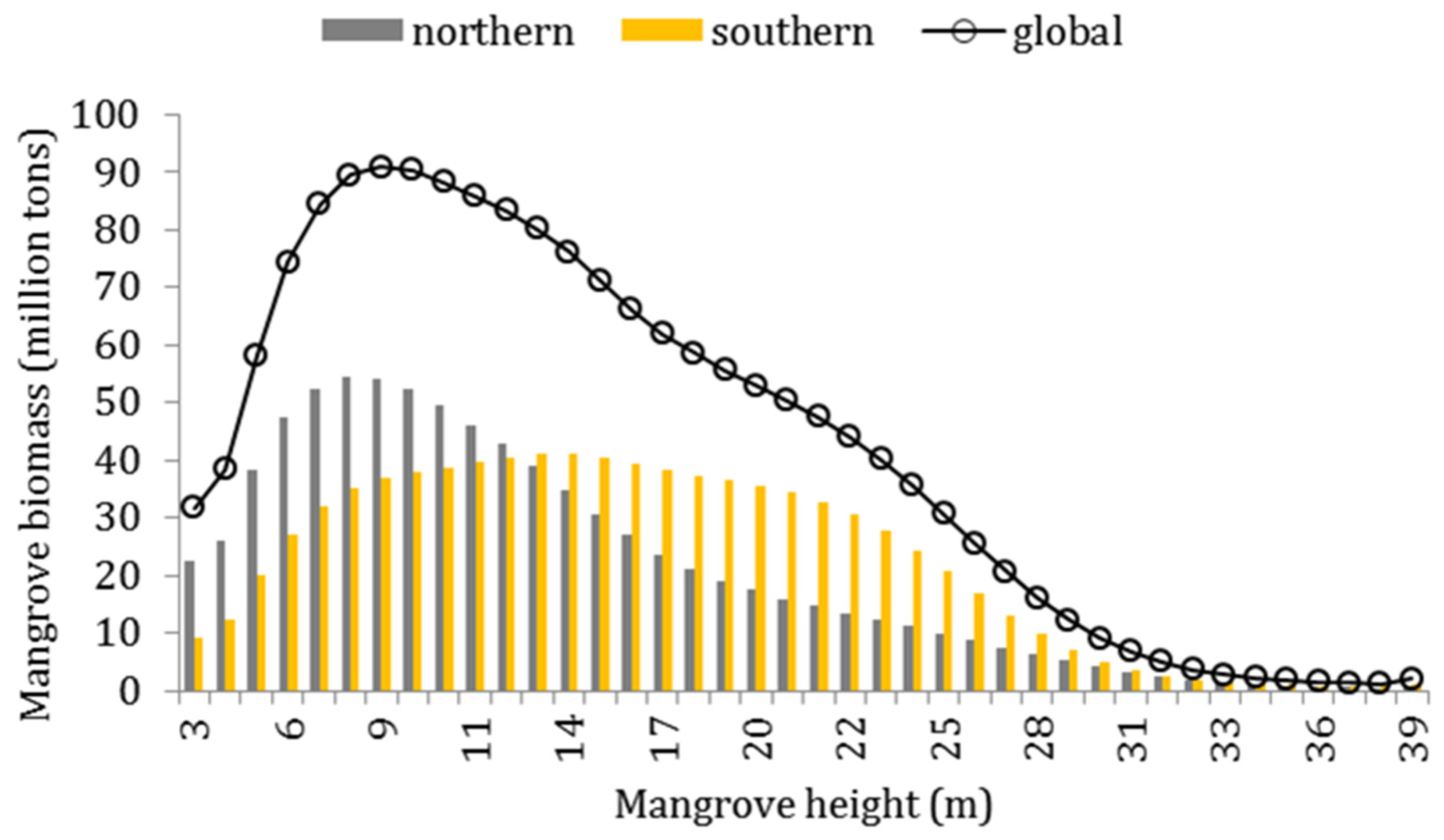
3.2. Mangrove Biomass
3.3. Mangrove-Rich Countries
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guannel, G.; Arkema, K.; Ruggiero, P.; Verutes, G. The Power of Three: Coral Reefs, Seagrasses and Mangroves Protect Coastal Regions and Increase Their Resilience. PLoS ONE 2016, 11, e0158094. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, J.; Spalding, M.; zu Ermgassen, P. The role of mangroves in fisheries enhancement. Nature Conserv. Wetl. Int. 2014, 54, 434. [Google Scholar]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the Most Carbon-Rich Forests in the Tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Pendleton, L.; Donato, D.C.; Murray, B.C.; Crooks, S.; Jenkins, W.A.; Sifleet, S.; Craft, C.; Fourqurean, J.W.; Kauffman, J.B.; Marbà, N.; et al. Estimating Global “Blue Carbon” Emissions from Conversion and Degradation of Vegetated Coastal Ecosystems. PLoS ONE 2012, 7, e43542. [Google Scholar] [CrossRef] [PubMed]
- Siikamäki, J.; Sanchirico, J.N.; Jardine, S.L. Global Economic Potential for Reducing Carbon Dioxide Emissions from Mangrove Loss. Proc. Natl. Acad. Sci. USA 2012, 109, 14369–14374. [Google Scholar] [CrossRef] [PubMed]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove Extinction Risk and Geographic Areas of Global Concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, S.E.; Casey, D. Creation of a High Spatio-Temporal Resolution Global Database of Continuous Mangrove Forest Cover for the 21st Century (CGMFC-21). Glob. Ecol. Biogeogr. 2016, 25, 729–738. [Google Scholar] [CrossRef]
- Osborne, D.J.; Spalding, M.D.; Blasco, F.; Field, C.D. (Eds.) World Mangrove Atlas. International Society for Mangrove Ecosystems, Okinawa 903-01, Japan. 178 pp. ISBN 4-906584-03-9. Price US$ 60.00 (Hardback). J. Trop. Ecol. 1998, 14, 723–724. [Google Scholar] [CrossRef]
- Giri, C.; Ochieng, E.; Tieszen, L.L.; Zhu, Z.; Singh, A.; Loveland, T.; Masek, J.; Duke, N. Status and Distribution of Mangrove Forests of the World Using Earth Observation Satellite Data. Glob. Ecol. Biogeogr. 2011, 20, 154–159. [Google Scholar] [CrossRef]
- Lagomasino, D.; Fatoyinbo, T.; Lee, S.K.; Feliciano, E.; Trettin, C.; Simard, M. A Comparison of Mangrove Canopy Height Using Multiple Independent Measurements from Land, Air, and Space. Remote Sens. 2016, 8, 327. [Google Scholar] [CrossRef]
- Persson, H.; Wallerman, J.; Olsson, H.; Fransson, J.E.S. Estimating Forest Biomass and Height Using Optical Stereo Satellite Data and a DTM from Laser Scanning Data. Can. J. Remote Sens. 2014, 39, 251–262. [Google Scholar] [CrossRef]
- Aljahdali, M.O.; Munawar, S.; Khan, W.R. Monitoring Mangrove Forest Degradation and Regeneration: Landsat Time Series Analysis of Moisture and Vegetation Indices at Rabigh Lagoon, Red Sea. Forests 2021, 12, 52. [Google Scholar] [CrossRef]
- Ellison, J.C. Factors Influencing Mangrove Ecosystems. In Mangroves: Ecology, Biodiversity and Management; Springer: Singapore, 2021; pp. 97–115. [Google Scholar] [CrossRef]
- Lugo, A.E.; Snedaker, S.C. The Ecology of Mangroves. Annu. Rev. Ecol. Syst. 2003, 5, 39–64. [Google Scholar] [CrossRef]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar] [CrossRef]
- Alongi, D.M. Present State and Future of the World’s Mangrove Forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Blasco, F.; Saenger, P.; Janodet, E. Mangroves as Indicators of Coastal Change. CATENA 1996, 27, 167–178. [Google Scholar] [CrossRef]
- Twilley, R.R.; Chen, R.H.; Hargis, T. Carbon Sinks in Mangroves and Their Implications to Carbon Budget of Tropical Coastal Ecosystems. Water Air Soil Pollut. 1992, 64, 265–288. [Google Scholar] [CrossRef]
- Duke, N.C.; Schmitt, K. Mangroves: Unusual Forests at the Seas Edge. In Tropical Forestry Handbook; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–24. [Google Scholar] [CrossRef]
- Simard, M.; Fatoyinbo, L.; Smetanka, C.; Rivera-Monroy, V.H.; Castañeda-Moya, E.; Thomas, N.; Van der Stocken, T. Mangrove Canopy Height Globally Related to Precipitation, Temperature and Cyclone Frequency. Nat. Geosci. 2019, 12, 40–45. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, Biomass, and Productivity of Mangrove Forests: A Review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Aslan, A.; Rahman, A.F.; Warren, M.W.; Robeson, S.M. Mapping Spatial Distribution and Biomass of Coastal Wetland Vegetation in Indonesian Papua by Combining Active and Passive Remotely Sensed Data. Remote Sens. Environ. 2016, 183, 65–81. [Google Scholar] [CrossRef]
- Simard, M.; Pinto, N.; Fisher, J.B.; Baccini, A. Mapping Forest Canopy Height Globally with Spaceborne Lidar. J. Geophys. Res. Biogeosci. 2011, 116, 4021. [Google Scholar] [CrossRef]
- Fatoyinbo, T.E.; Simard, M. Height and Biomass of Mangroves in Africa from ICESat/GLAS and SRTM. Int. J. Remote Sens. 2012, 34, 668–681. [Google Scholar] [CrossRef]
- Fatoyinbo, T.E.; Simard, M.; Washington-Allen, R.A.; Shugart, H.H. Landscape-Scale Extent, Height, Biomass, and Carbon Estimation of Mozambique’s Mangrove Forests with Landsat ETM+ and Shuttle Radar Topography Mission Elevation Data. J. Geophys. Res. Biogeosci. 2008, 113, G2. [Google Scholar] [CrossRef]
- Simard, M.; Rivera-Monroy, V.H.; Mancera-Pineda, J.E.; Castañeda-Moya, E.; Twilley, R.R. A Systematic Method for 3D Mapping of Mangrove Forests Based on Shuttle Radar Topography Mission Elevation Data, ICEsat/GLAS Waveforms and Field Data: Application to Ciénaga Grande de Santa Marta, Colombia. Remote Sens. Environ. 2008, 112, 2131–2144. [Google Scholar] [CrossRef]
- Lee, S.K.; Fatoyinbo, T.; Lagomasino, D.; Osmanoglu, B.; Simard, M.; Trettin, C.; Rahman, M.; Ahmed, I. Large-Scale Mangrove Canopy Height Map Generation from TanDEM-X Data by Means of Pol-InSAR Techniques. Int. Geosci. Remote Sens. Symp. 2015, 2015, 2895–2898. [Google Scholar] [CrossRef]
- Aslan, A.; Rahman, A.F.; Robeson, S.M. Investigating the Use of Alos Prism Data in Detecting Mangrove Succession through Canopy Height Estimation. Ecol. Indic. 2017, 87, 136–143. [Google Scholar] [CrossRef]
- Ximenes, A.C.; Ponsoni, L.; Lira, C.F.; Koedam, N.; Dahdouh-Guebas, F. Does Sea Surface Temperature Contribute to Determining Range Limits and Expansion of Mangroves in Eastern South America (Brazil)? Remote Sens. 2018, 10, 1787. [Google Scholar] [CrossRef]
- Woodroffe, C.D.; Grindrod, J. Mangrove Biogeography: The Role of Quaternary Environmental and Sea-Level Change. J. Biogeogr. 1991, 18, 479. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Donato, D.C. Protocols for the Measurement, Monitoring and Reporting of Structure, Biomass and Carbon Stocks in Mangrove Forests; CIFOR: Bogor, Indonesia, 2012. [Google Scholar]
- Mizanur Rahman, M.; Nabiul Islam Khan, M.; Fazlul Hoque, A.K.; Ahmed, I. Carbon Stock in the Sundarbans Mangrove Forest: Spatial Variations in Vegetation Types and Salinity Zones. Wetl. Ecol. Manag. 2015, 23, 269–283. [Google Scholar] [CrossRef]
- Kangkuso, A.; Jamili, J.; Septiana, A.; Raya, R.; Sahidin, I.; Rianse, U.; Rahim, S.; Alfirman, A.; Sharma, S.; Nadaoka, K. Allometric models and aboveground biomass of Lumnitzera racemosa Willd. Forest in Rawa Aopa Watumohai National Park, Southeast Sulawesi, Indonesia. For. Sci. Technol. 2016, 12, 43–50. [Google Scholar]
- Bismark, M.; Subiandono, E.; Heriyanto, N.M. Keragaman dan potensi jenis serta kandungan karbon hutan mangrove di Sungai Subelen Siberut, Sumatera Barat. J. Penelit. Hutan Konserv. Alam 2008, 5, 297–306. [Google Scholar] [CrossRef]
- Wannasiri, W.; Nagai, M.; Honda, K.; Santitamnont, P.; Miphokasap, P. Extraction of mangrove biophysical parameters using airborne LiDAR. Remote Sens. 2013, 5, 1787–1808. [Google Scholar] [CrossRef]
- Wang, G.; Guan, D.M.; Peart, R.; Chen, Y.; Peng, Y. Ecosystem carbon stocks of mangrove forest in Yingluo Bay, Guangdong Province of South China. For. Ecol. Manag. 2013, 310, 539–546. [Google Scholar] [CrossRef]
- Goessens, A.; Satyanarayana, B.; van der Stocken, T.; Zuniga, M.Q.; Mohd-Lokman, H.; Sulong, I.; Dahdouh-Guebas, F. Is Matang Mangrove Forest in Malaysia sustainably rejuvenating after more than a century of conservation and harvesting management? PLoS ONE 2014, 9, e105069. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.; Kamal, A.H.M.; Rosli, Z.; Hakeem, K.R.; Hoque, M.M. Composition and diversity of plants in Sibuti mangrove forest, Sarawak, Malaysia. For. Sci. Technol. 2016, 12, 70–76. [Google Scholar] [CrossRef]
- Rajkumar, S.Y.; Ketan, M.; Salvi, H.; Kamboj, R.D. Age and Growth relation of mangrove Avicennia marina (Forssk.) Vierh. In Gulf of Kachchh (GoK), India. Gulf of Kachchh (Gok), India (22 June 2018). Appl. Sci. Rep. 2017, 17. [Google Scholar] [CrossRef]
- Krauss, K.W.; Thomas, W.D.; Twilley, R.R.; Smith, T.J., III; Whelan, K.R.T.; Sullivan, K.J. Woody Debris in the Mangrove Forests of South Florida 1. Biotrop. J. Biol. Conserv. 2005, 37, 9–15. [Google Scholar]
- Lovelock, C.E.; Marilyn, C.B.; Martin, K.C.; Feller, I.C. Nutrient enrichment increases mortality of mangroves. PLoS ONE 2009, 4, e5600. [Google Scholar] [CrossRef]
- Loría-Naranjo, M.; Samper-Villarreal, J.; Cortés, J. Structural complexity and species composition of Potrero Grande and Santa Elena mangrove forests in Santa Rosa National Park, North Pacific of Costa Rica. Rev. Biol. Trop. 2014, 62, 33–41. [Google Scholar] [CrossRef]
- Blanco, J.F.; Bejarano, A.C.; Lasso, J.; Cantera, J.R. A new look at computation of the complexity index in mangroves: Do disturbed forests have clues to analyze canopy height patchiness? Wetl. Ecol. Manag. 2011, 9, 91–101. [Google Scholar] [CrossRef]
- Zamprogno, G.C.; Tognella, M.M.P.; Quaresma, V.D.S.; da Costa, M.B.; Pascoalini, S.S.; Couto, G.F.D. The structural heterogeneity of an urbanised mangrove forest area in southeastern Brazil: Influence of environmental factors and anthropogenic stressors. Braz. J. Oceanogr. 2016, 64, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Bandeira, S.O.; Macamo, C.C.F.; Kairo, J.G.; Amade, F.; Jiddawi, N.; Paula, J. Evaluation of mangrove structure and condition in two trans-boundary areas in the Western Indian Ocean. Aquatic Conserv. Mar. Freshwater Ecosyst. 2009, 19, S46–S55. [Google Scholar] [CrossRef]
- Simard, M.; Zhang, K.; Rivera-Monroy, V.; Ross, M.; Ruiz, P.; Castañeda-Moya, E.; Twilley, R.; Rodriguez, E. Mapping Height and Biomass of Mangrove Forests in Everglades National Park with SRTM Elevation Data. Photogramm. Eng. Remote Sens. 2006, 72, 299–311. [Google Scholar] [CrossRef]
- Ramsey, E.; Lu, Z.; Rangoonwala, A.; Rykhus, R. Multiple Baseline Radar Interferometry Applied to Coastal Land Cover Classification and Change Analyses. GISci. Remote Sens. 2006, 43, 283–309. [Google Scholar] [CrossRef]
- Pohjankukka, J.; Pahikkala, T.; Nevalainen, P.; Heikkonen, J. Estimating the Prediction Performance of Spatial Models via Spatial K-Fold Cross Validation. Int. J. Geogr. Inf. Sci. 2017, 31, 2001–2019. [Google Scholar] [CrossRef]
- Steyerberg, E.W.; Vickers, A.J.; Cook, N.R.; Gerds, T.; Gonen, M.; Obuchowski, N.; Pencina, M.J.; Kattan, M.W. Assessing the Performance of Prediction Models: A Framework for Some Traditional and Novel Measures. Epidemiology 2010, 21, 128. [Google Scholar] [CrossRef]
- Saenger, P.; Snedaker, S.C. Pantropical Trends in Mangrove Above-Ground Biomass and Annual Litterfall. Oecologia 1993, 96, 293–299. [Google Scholar] [CrossRef]
- Cintrón, G.; Schaeffer-Novelli, Y. Características y desarrollo estructural de los manglares de Norte y Sur América. Ciencia Interamericana 1985, 25, 4–15. [Google Scholar]
- Luther, D.A.; Greenberg, R. Mangroves: A Global Perspective on the Evolution and Conservation of Their Terrestrial Vertebrates. Bioscience 2009, 59, 602–612. [Google Scholar] [CrossRef]
- Rovai, A.S.; Twilley, R.R.; Castañeda-Moya, E.; Midway, S.R.; Friess, D.A.; Trettin, C.C.; Bukoski, J.J.; Stovall, A.E.L.; Pagliosa, P.R.; Fonseca, A.L.; et al. Macroecological Patterns of Forest Structure and Allometric Scaling in Mangrove Forests. Glob. Ecol. Biogeogr. 2021, 30, 1000–1013. [Google Scholar] [CrossRef]
- Boone Kauffman, J.; Arifanti, V.B.; Hernández Trejo, H.; del Carmen Jesús García, M.; Norfolk, J.; Cifuentes, M.; Hadriyanto, D.; Murdiyarso, D. The Jumbo Carbon Footprint of a Shrimp: Carbon Losses from Mangrove Deforestation. Front. Ecol. Environ. 2017, 15, 183–188. [Google Scholar] [CrossRef]
- Barbier, E.B. Progress and Challenges in Valuing Coastal and Marine Ecosystem Services. Rev. Environ. Econ. Policy 2020, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Rossi, C.; Gernhardt, S. Urban DEM Generation, Analysis and Enhancements Using TanDEM-X. ISPRS J. Photogramm. Remote Sens. 2013, 85, 120–131. [Google Scholar] [CrossRef]
- Jardine, S.L.; Siikamäki, J.V. A Global Predictive Model of Carbon in Mangrove Soils. Environ. Res. Lett. 2014, 9, 104013. [Google Scholar] [CrossRef]
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Location | SRTM Elevation | Field Mangrove Height | Longitude | Latitude | Mangrove Species | References |
|---|---|---|---|---|---|---|---|
| 1 | Mahakam Delta, Indonesia | 5.0 | 10.0 | 117.5296327 | −0.614998372 | R. mucronata, A. germinans, Nypa fruticans, S. alba, R. apiculata, Bruguiera sp. | Unpublished field data |
| 2 | Mimika Papua, Indonesia | 7.0 | 8.0 | 136.7123396 | −4.797423469 | R. mucronata, R. apiculata, R. stylosa, Avicennia sp., Sonneratia sp., Nypa fruticans, S. alba, R. apiculata, B. exaristat, B. gymnorrhiza, B. hainesii, B. parviflora, B. sexangula, B. cylindrica, Ceriops, Camptostemon, Lumnitzera | Unpublished field data |
| 3 | Sundarbans, Bangladesh | 10.0 | 9.2 | 89.49642047 | 22.07774034 | Heritiera fomes, Nypa fruticans, Bruguiera gymnorrhiza, R. apiculata, R. mucronata, Xylocarpus granatum, X. mekongensis | Unpublished field data |
| 4 | Asmat Papua, Indonesia | 21.6 | 25.0 | 138.0509175 | −5.47876875 | R. mucronata, R. apiculata, R. stylosa, Avicennia sp., Sonneratia sp., Nypa fruticans, S. alba, R. apiculata, B. exaristat, B. gymnorrhiza, B. hainesii, B. parviflora, B. sexangula, B. cylindrica, Ceriops, Camptostemon | Unpublished field data |
| 5 | Arguni Bay Papua, Indonesia | 18.7 | 19.4 | 133.7809701 | −3.07832475 | R. mucronata, R. apiculata, R. stylosa, Avicennia sp., Sonneratia sp., Nypa fruticans, S. alba, R. apiculata, Bruguiera sp. | Unpublished field data |
| 6 | Southeast Sulawesi-Indonesia | 9.0 | 8.2 | 122.056104 | −4.553076 | L. racemosa | Kangkuso et al. [33] |
| 7 | Siberut, Indonesia | 20.0 | 20.5 | 99.0538105 | −1.3072485 | R. apiculata, R. mucronata, B. cylindrica, B. gymnorrhiza, Xylocarpus granatum, Barringtonia racemosa, Ceriops tagal, Aegyceras corniculatum, Lumnitzera littorea, Avicennia alba | Bismark et al. [34] |
| 8 | Thailand | 3.0 | 2.8 | 102.1529199 | 12.52941738 | Avicennia alba, Avicennia officinalis | Wannasiri et al. [35] |
| 9 | Yingluo Bay, China | 2.0 | 2.4 | 109.759312 | 21.56500025 | Avicennia marina, Sonneratia apetala, A. corniculatum, K. obavata, B. gymnorrhiza, R. stylosa | Wang et al. [36] |
| 10 | Matang, Malaysia | 17.5 | 14.8 | 100.6003847 | 4.853099667 | R. apiculata, B. parvilora, B. sexangula, R. mucronata, Avicennia alba | Goessens et al. [37] |
| 11 | Sibuti Serawak, Malaysia | 21.0 | 19.6 | 113.736945 | 3.987122667 | R. apiculata, X. granatum, X. mekongensis, Nypa fruticans, Intsia bijuga, Thespesia populnea, Excoecaria agallocha, Acrostichum speciosum, Phoenix paludosa | Shah et al. [38] |
| 12 | Gulf of Kutch, India | 4.0 | 4.0 | 69.87755175 | 22.48217463 | A. marina | Rajkumar et al. [39] |
| 13 | Everglades Florida, United States | 5.3 | 9.9 | −81.06102613 | 25.4852244 | R. mangle, A. germinans, L. racemosa | Krauss et al. [40] |
| 14 | Brisbane, Australia | 8.7 | 8.5 | 153.033468 | −27.282594 | A. marina | Lovelock et al. [41] |
| 15 | Exmouth, Australia | 4.0 | 1.5 | 113.94707 | −21.961995 | A. marina | Lovelock et al. [41] |
| 16 | Hinchinbrook, Australia | 6.0 | 6.0 | 146.166667 | −18.333333 | R. mangle, A. germinans, R. lamarckii | Lovelock et al. [41] |
| 17 | Port Douglas, Australia | 5.0 | 2.0 | 145.44973 | −16.499527 | R. mangle, A. marina | Lovelock et al. [41] |
| 18 | Twin Cay, Belize | 4.0 | 7.0 | −88.100419 | 16.832535 | R. mangle | Lovelock et al. [41] |
| 19 | Potrero Grande, Costa Rica | 14.0 | 8.8 | −85.786436 | 10.851285 | A. germinans, L. racemosa, R. mangle, R. racemosa, P. rhizophorae | Loría-Naranjo et al. [42] |
| 20 | Santa Elena, Costa Rica | 10.1 | 7.0 | −85.78448 | 10.91266 | A. germinans, A. bicolor, L. racemosa, R. mangle, R. racemosa, P. rhizophorae | Loría-Naranjo et al. [42] |
| 21 | Buenaventura Bay, Colombia | 23.0 | 24.2 | −77.091394 | 3.830060111 | R. mangle, R. racemosa, A. germinans, L. racemosa, Pelliciera rhizophorae | Blanco et al. [43] |
| 22 | Vitoria Bay, Brazil | 4.6 | 7.9 | −40.33386427 | −20.27102091 | Avicennia schaueriana, L. racemosa, R. mangle | Zamprogno et al. [44] |
| 23 | Mngoji2, Mozambique | 7.8 | 3.8 | 40.36687 | −10.361206 | A. marina, C. tagal, R. mucronata, S. alba | Bandeira et al. [45] |
| 24 | Mngoji1, Mozambique | 7.5 | 5.2 | 40.414096 | −10.346828 | A. marina, R. mucronata, S. alba | Bandeira et al. [45] |
| 25 | Ulo, Mozambique | 7.3 | 2.7 | 40.447664 | −11.415561 | A. marina, C. tagal, R. mucronata, S. alba | Bandeira et al. [45] |
| 26 | Luchete, Mozambique | 8.4 | 2.2 | 40.425148 | −11.585771 | A. marina, C. tagal, R. mucronata, S. alba | Bandeira et al. [45] |
| 27 | Ibo, Mozambique | 6.8 | 3.0 | 40.57073 | −12.384053 | A. marina, C. tagal, R. mucronata, S. alba | Bandeira et al. [45] |
| 28 | Pemba, Mozambique | 7.3 | 3.4 | 40.485629 | −13.050629 | A. marina, B. gymnorhiza, C. tagal, R. mucronata, S. alba | Bandeira et al. [45] |
| 29 | Mecufi, Mozambique | 7.0 | 3.2 | 40.54862 | −13.296666 | A. marina, B. gymnorhiza, C. tagal, R. mucronata, S. alba | Bandeira et al. [45] |
| 30 | Saco, Mozambique | 5.0 | 3.9 | 32.914154 | −26.035665 | A. marina, B. gymnorhiza, C. tagal, R. mucronata | Bandeira et al. [45] |
| 31 | Sangala, Mozambique | 5.8 | 2.5 | 32.944879 | −25.991996 | A. marina, B. gymnorhiza, C. tagal, R. mucronata | Bandeira et al. [45] |
| No | Country | MFW (ha) | CGMFC-21 (ha) | This Study (ha) | Tall ≥20 m (ha) | Short to Medium <20 m (ha) | Aboveground Biomass (tons) |
|---|---|---|---|---|---|---|---|
| 1 | Indonesia | 3,112,989 | 2,407,313 | 2,393,244 | 562,596 | 1,830,648 | 472,150,415 |
| 2 | Brazil | 962,683 | 772,131 | 853,902 | 112,345 | 741,556 | 150,725,476 |
| 3 | Australia | 977,975 | 332,651 | 770,167 | 7806 | 762,361 | 98,882,395 |
| 4 | Nigeria | 653,669 | 265,704 | 568,235 | 25,147 | 543,088 | 74,975,813 |
| 5 | Malaysia | 505,386 | 496,868 | 497,116 | 20,937 | 476,178 | 83,931,536 |
| 6 | Papua New Guinea | 480,121 | 418,992 | 426,987 | 122,093 | 304,894 | 93,626,647 |
| 7 | Mexico | 741,917 | 302,103 | 390,922 | 9803 | 381,120 | 54,288,866 |
| 8 | Bangladesh | 436,570 | 177,390 | 366,710 | 853 | 365,856 | 52,675,523 |
| 9 | Myanmar | 494,584 | 279,260 | 323,398 | 12,893 | 310,505 | 47,050,344 |
| 10 | Mozambique | 318,851 | 122,620 | 276,396 | 556 | 275,840 | 32,788,751 |
| 11 | Guinea Bissau | 338,652 | 74,518 | 238,901 | 891 | 238,010 | 29,593,836 |
| 12 | Cuba | 421,538 | 166,036 | 227,513 | 240 | 227,273 | 28,682,781 |
| 13 | Philippines | 263,137 | 209,105 | 218,883 | 2029 | 216,854 | 25,492,129 |
| 14 | Madagascar | 278,078 | 85,222 | 214,914 | 1057 | 213,857 | 30,091,223 |
| 15 | India | 368,276 | 82,506 | 159,770 | 2940 | 156,830 | 20,015,239 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aslan, A.; Aljahdali, M.O. Characterizing Global Patterns of Mangrove Canopy Height and Aboveground Biomass Derived from SRTM Data. Forests 2022, 13, 1545. https://doi.org/10.3390/f13101545
Aslan A, Aljahdali MO. Characterizing Global Patterns of Mangrove Canopy Height and Aboveground Biomass Derived from SRTM Data. Forests. 2022; 13(10):1545. https://doi.org/10.3390/f13101545
Chicago/Turabian StyleAslan, Aslan, and Mohammed Othman Aljahdali. 2022. "Characterizing Global Patterns of Mangrove Canopy Height and Aboveground Biomass Derived from SRTM Data" Forests 13, no. 10: 1545. https://doi.org/10.3390/f13101545
APA StyleAslan, A., & Aljahdali, M. O. (2022). Characterizing Global Patterns of Mangrove Canopy Height and Aboveground Biomass Derived from SRTM Data. Forests, 13(10), 1545. https://doi.org/10.3390/f13101545