
Developing Tree Mortality Models Using Bayesian Modeling Approach



Abstract
:1. Introduction
2. Materials and Methods
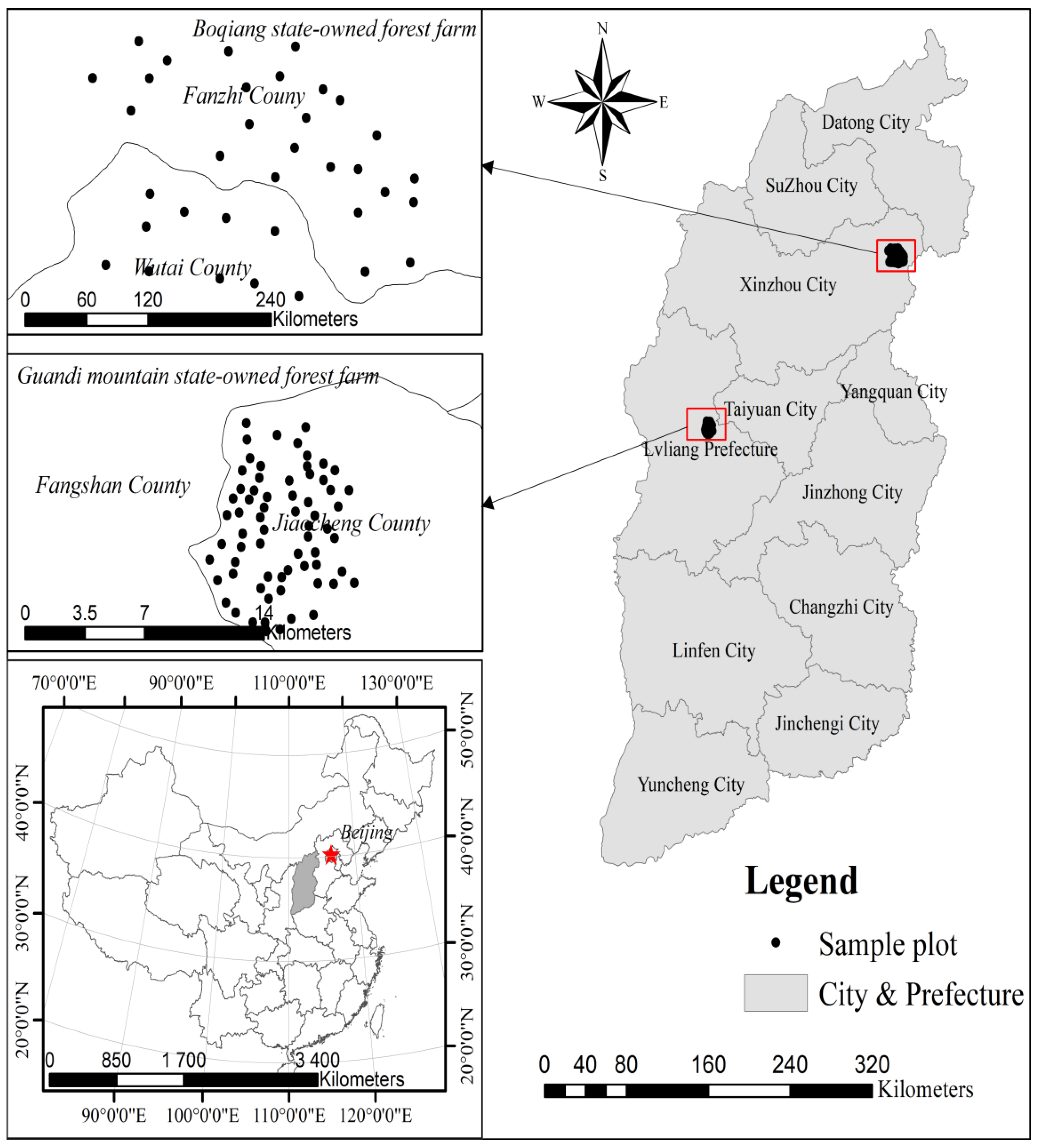
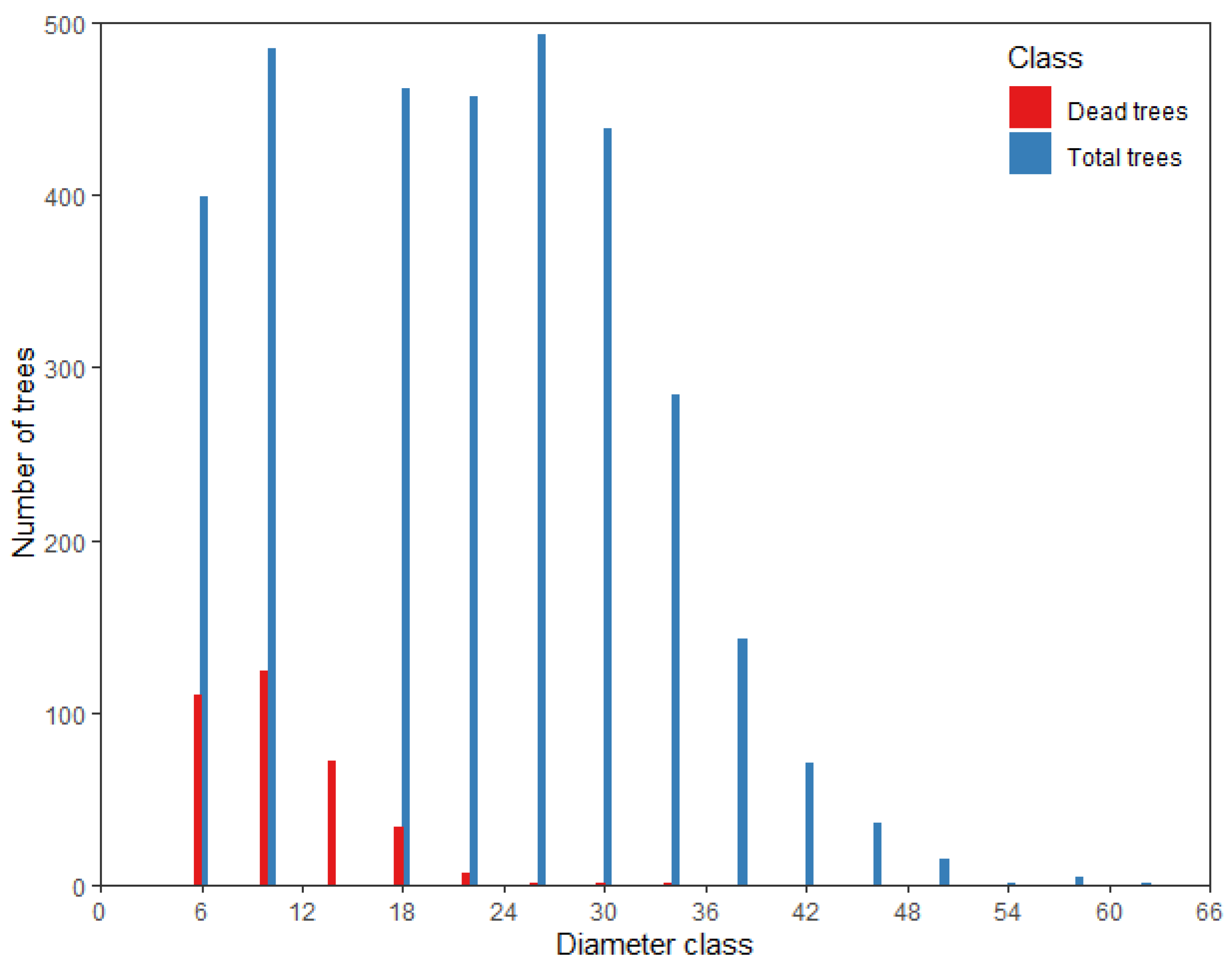
2.1. Data
2.2. Selection of Predictor Variables
3. Model Development
3.1. Individual Tree Variables
3.2. Stand Variables
3.3. Climate Variables
3.4. Two-Level Models
3.5. Model Evaluation
4. Results
4.1. Model Evaluation
4.2. Two-Level Mortality Model
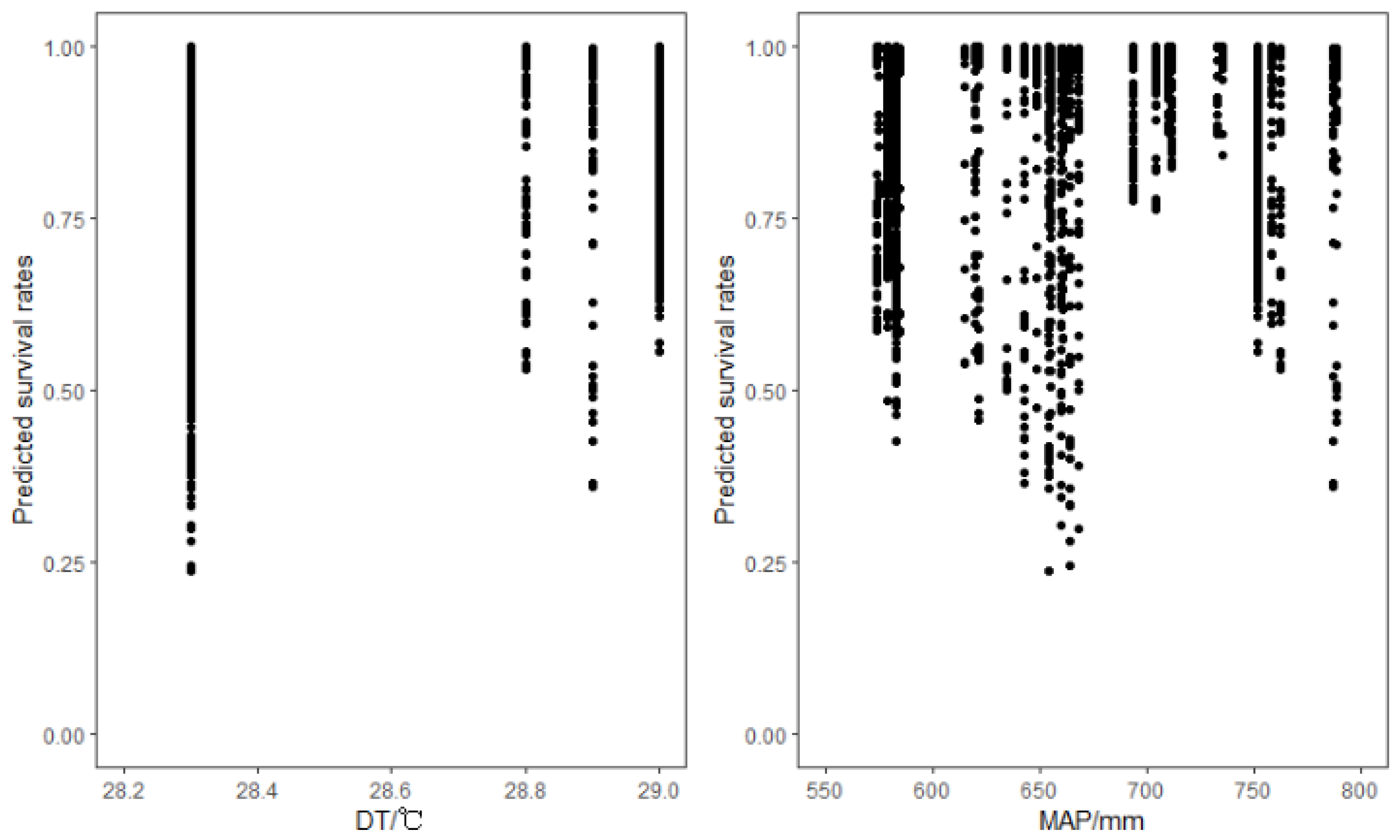
4.3. Climate Effects on Tree Mortality
5. Discussion
6. Conclusions
- (1)
- The best model included the predictor variables at three levels: Individual tree- and stand-, and environmental (climate)-levels in the Bayesian logistic models.
- (2)
- The Bayesian two-level model, which includes tree-level, stand-level, and climatic predictor variables, outperformed all the other forms of the models, describing larger variations of tree mortality and accounting for multiple sources of the unobserved heterogeneities.
- (3)
- Tree mortality significantly positively correlated with the sum of squares of tree diameters larger than the estimated diameter, and mean annual precipitation, but negatively correlated to the ratio of the diameter to the average square diameter of stand, the stand arithmetic mean diameter, and the mean of difference in temperature.
- (4)
- Presented mortality models will have significant implications for identifying different factors affecting tree mortality and precise prediction of the mortality.
- (5)
- With the mortality data collected from a wider distribution of the tree species of interest and advanced modeling techniques, the prediction performance of the tree mortality models may be improved, which we aim for in the future.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, X.; Lei, Y.; Cao, Q.V.; Chen, X.; Liu, X. Improving tree survival prediction with forecast combination and disaggregation. Can. J. For. Res. 2011, 41, 1928–1935. [Google Scholar] [CrossRef] [Green Version]
- Monserud, R.A.; Sterba, H. Modeling individual tree mortality for Austrian forest species. For. Ecol. Manag. 1999, 113, 109–123. [Google Scholar] [CrossRef]
- Eid, T.; Tuhus, E. Models for individual tree mortality in Norway. For. Ecol. Manag. 2001, 154, 69–84. [Google Scholar] [CrossRef]
- Yao, X.H.; Titus, S.J.; MacDonald, S.E. A generalized logistic model of individual tree mortality for aspen, white spruce, and lodgepole pine in Alberta mixedwood forests. Can. J. For. Res. 2001, 31, 283–291. [Google Scholar] [CrossRef]
- Das, A.J.; Stephenson, N.L. Improving estimates of tree mortality probability using potential growth rate. Can. J. For. Res. 2015, 45, 920–928. [Google Scholar] [CrossRef]
- Wyckoff, P.H.; Clark, J.S. Predicting tree mortality from diameter growth: A comparison of maximum likelihood and Bayesian approaches. Can. J. For. Res. 2000, 30, 156–167. [Google Scholar] [CrossRef]
- Adame, P.; Miren, D.R.; Caellas, I. Modeling individual-tree mortality in Pyrenean oak (Quercus pyrenaica Willd.) stands. Ann. For. Sci. 2010, 67, 1–10. [Google Scholar] [CrossRef]
- Chao, K.J.; Phillips, O.L.; Gloor, E.; Monteagudo, A.; Torres-Lezama, A.; Martinez, R.V. Growth and wood density predict tree mortality in Amazon forests. J. Ecol. 2008, 96, 281–292. [Google Scholar] [CrossRef]
- Crecente-Campo, F.; Marshall, P.; Rodríguez-Soalleiro, R. Modeling non-catastrophic individual-tree mortality for Pinus radiata plantations in northwestern Spain. For. Ecol. Manag. 2009, 257, 1542–1550. [Google Scholar] [CrossRef]
- Ruiz-Benito, P.; Lines, E.R.; Gómez-Aparicio, L.; Zavala, M.A.; Coomes, D.A. Patterns and drivers of tree mortality in Iberian forests: Climatic effects are modified by competition. PLoS ONE 2013, 8, e56843. [Google Scholar] [CrossRef] [Green Version]
- Adams, H.D.; Guardiola-Claramonte, M.; Barron-Gafford, G.A.; Villegas, J. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. USA 2009, 106, 7063–7066. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Lei, X.; Zhu, Q.; Cheng, H.; Peng, C. A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nat. Clim. Chang. 2011, 1, 467–471. [Google Scholar]
- Brando, P.M.; Balch, J.K.; Nepstad, D.C.; Morton, D.C.; Putz, F.E.; Coe, M.T.; Silverio, D.; Macedo, M.N.; Davidson, E.A.; Nobrega, C.C. Abrupt increases in Amazonian tree mortality due to drought–fire interactions. Proc. Natl. Acad. Sci. USA 2014, 111, 6347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.Q.; Cao, Q.V.; Duan, A.G.; Zhang, A.G. Modeling tree mortality in relation to climate, initial planting density and competition in Chinese fir plantations using a Bayesian logistic multilevel method. Can. J. For. Res. 2017, 49, 1278–1285. [Google Scholar] [CrossRef]
- Qiu, S.; Xu, M.; Xu, R.Q.; Li, Y.P.; Zheng, Y.; Daniel, C.; Cui, X.W.; Cui, L.X.; Liu, C.H.; Zhang, L.W.; et al. Climatic information improves statistical individual-tree mortality models for three key species of Sichuan Province, China. Ann. For. Sci. 2015, 72, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Q.; Lei, Y.C.; Liu, X.Z. Modeling stand mortality using Poisson mixture models with mixed-effects. Ifor.-Biogeosci. For. 2015, 8, 333–338. [Google Scholar] [CrossRef]
- Lutz, J.A.; Halpern, C.B. Tree mortality during early forest development: A long-term study of rates, causes, and consequences. Ecol. Monogr. 2006, 76, 257–275. [Google Scholar] [CrossRef]
- Larson, A.J.; Franklin, J.F. The tree mortality regime in temperate old-growth coniferous forests: The role of physical damage. Can. J. For. Res. 2010, 40, 2091–2103. [Google Scholar] [CrossRef] [Green Version]
- Van Mantgem, P.J.; Stephenson, N.L.; Byrne, J.C.; Daniels, L.D.; Franklin, J.F.; Fule, P.Z.; Harmon, M.E.; Larson, A.J.; Smith, J.M.; Taylor, A.H.; et al. Widespread increase of tree mortality rates in the western United States. Science 2009, 323, 521–524. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, D.A. Event Probabilities Estimated by Regression; Intermountain Forest & Range Experiment Station, Forest Service, US Department of Agriculture: Fort Collins, CO, USA, 1974. [Google Scholar]
- Weingartner, M.M. Estimating tree mortality of Norway spruce stands with neural networks. Adv. Environ. Res. 2001, 5, 405–414. [Google Scholar]
- Rose, C.E., Jr.; Hall, D.B.; Shiver, B.D.; Clutter, M.L.; Borders, B. A multilevel approach to individual tree survival prediction. For. Sci. 2006, 52, 31–43. [Google Scholar]
- Yang, Y.; Titus, S.J.; Huang, S. Modeling individual tree mortality for white spruce in Alberta. Ecol. Model. 2003, 163, 209–222. [Google Scholar] [CrossRef]
- Boeck, A.; Dieler, J.; Biber, P.; Pretzsch, H.; Ankerst, D.P. Predicting tree mortality for European beech in southern Germany using spatially explicit competition indices. For. Sci. 2014, 60, 613–622. [Google Scholar] [CrossRef]
- Coble, D.W.; Cao, Q.V.; Jordan, L. An Annual Tree Survival and Diameter Growth Model for Loblolly and Slash Pine Plantations in East Texas. South. J. Appl. For. 2012, 36, 79–84. [Google Scholar] [CrossRef]
- Moser, J.W. Dynamics of an Uneven-Aged Forest Stand. For. Sci. 1972, 18, 184–191. [Google Scholar]
- Hamilton, D.A., Jr. Extending the range of applicability of an individual tree mortality model. Can. J. For. Res. 1990, 20, 1212–1218. [Google Scholar] [CrossRef]
- Adams, H.D.; Williams, A.P.; Xu, C.; Rauscher, S.A.; Jiang, X.; McDowell, N.G. Empirical and process-based approaches to climate-induced forest mortality models. Front. Plant Sci. 2013, 4, 438. [Google Scholar] [CrossRef] [Green Version]
- Somers, G.L.; Oderwald, R.G.; Harms, W.R.; Langdon, O.G. Predicting Mortality with a Weibull Distribution. For. Sci. 1980, 26, 291–300. [Google Scholar]
- Holzwarth, F.; Kahl, A.; Bauhus, J.; Wirth, C. Many ways to die—Partitioning tree mortality dynamics in a near-natural mixed deciduous forest. J. Ecol. 2013, 101, 220–230. [Google Scholar] [CrossRef]
- Buford, M.A.; Hafley, W.L. Probability distributions as models for mortality. For. Sci. 1985, 31, 331–341. [Google Scholar]
- Kobe, R.K.; Coates, K.D. Models of sapling mortality as a function of growth to characterize interspecific variation in shade tolerance of eight tree species of northwestern British Columbia. Can. J. For. Res. 1997, 27, 227–236. [Google Scholar] [CrossRef]
- Travis, W.; Shaw, D.C.; Ganio, L.M.; Fitzgerald, S. A review of logistic regression models used to predict post-fire tree mortality of western North American conifers. Int. J. Wildland Fire 2012, 21, 1–35. [Google Scholar]
- Zeng, Z.; Yin, G.; Zhang, Y.; Sun, Y.; Wang, T.; Piao, S. MODIS based estimation of forest aboveground biomass in China. PLoS ONE 2015, 10, e0130143. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Zhang, H.; Sharma, R.P.; Pang, L.; Wang, G. A generalized nonlinear mixed-effects height to crown base model for Mongolian oak in northeast China. For. Ecol. Manag. 2017, 384, 34–43. [Google Scholar] [CrossRef]
- Fu, L.; Lei, Y.; Affleck, D.L.; Nelson, A.S.; Shen, C.; Wag, M.; Zheng, J.; Ye, Q.; Yang, G. Additivity of nonlinear tree crown width models: Aggregated and disaggregated model structures using nonlinear simultaneous equations. For. Ecol. Manag. 2018, 427, 372–382. [Google Scholar]
- West, P.W.; Ratkowsky, D.A.; Davis, A.W. Problems of hypothesis testing of regressions with multiple measurements from individual sampling units. For. Ecol. Manag. 1984, 7, 207–224. [Google Scholar] [CrossRef]
- Chen, D.; Huang, X.; Sun, X.; Ma, W.; Zhang, S. A Comparison of Hierarchical and Non-Hierarchical Bayesian Approaches for Fitting Allometric Larch (Larix. spp.) Biomass Equations. Forests 2016, 7, 18. [Google Scholar] [CrossRef] [Green Version]
- Mauricio, Z.C.; Carlos, A.S.; Lauren, A. Probability distribution of allometric coefficients and Bayesian estimation of aboveground tree biomass. For. Ecol. Manag. 2012, 277, 173–179. [Google Scholar]
- Zhang, X.; Zhang, J.; Duan, A. A Hierarchical Bayesian Model to Predict Self-Thinning Line for Chinese Fir in Southern China. PLoS ONE 2015, 10, e0139788. [Google Scholar]
- Zhang, X.; Duan, A.; Zhang, J. Tree Biomass Estimation of Chinese fir (Cunninghamia lanceolata) Based on Bayesian Method. PLoS ONE 2013, 8, e79868. [Google Scholar] [CrossRef] [Green Version]
- Anholt, B.R.; Werner, E.; Skelly, D.K. Effect of Food and Predators on the Activity of Four Larval Ranid Frogs. Ecology 2000, 81, 3509–3521. [Google Scholar] [CrossRef]
- Leng, W.F.; He, H.S.; Liu, H.J. Response of larch species to climate changes. J. Plant Ecol. 2008, 1, 203–205. [Google Scholar] [CrossRef] [Green Version]
- Tao, B.; Cao, M.K.; Gui, R.; Liu, J.Y.; Wang, S.Q. Global Carbon Project (GCP) Beijing Office: A new bridge for understanding regional carbon cycles. J. Geogr. Inf. Syst. 2006, 016, 375–377. [Google Scholar]
- Chen, H.; Xu, Z.B. Preliminary study on the tree death of Korean pine deciduous mixed forest of Changbai Mountain. Chin. J. Appl. Ecol. 1991, 2, 89–91. [Google Scholar]
- Ban, Y.; Xu, H.C.; Li, Z.D. Mortality patterns of Larix gmelini and effect of fallen dead wood on regeneration of old Larixgmeliforest. Chin. J. Appl. Ecol. 1997, 8, 449. [Google Scholar]
- Chambers, J.Q.; Negron-Juarez, R.I.; Marra, D.M.; Di Vittorio, A.; Tews, J.; Roberts, D.; Ribeiro, G.H.; Trumbore, S.E.; Higuchi, N. The steady-state mosaic of disturbance and succession across an old-growth Central Amazon forest landscape. Proc. Natl. Acad. Sci. USA 2013, 110, 3949–3954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erb, K.H.; Fetzel, T.; Plutzar, C.; Kastner, T.; Lauk, C.; Mayer, C.; Niedertscheider, M.; KöRner, C.; Haberl, H. Biomass turnover time in terrestrial ecosystems halved by land use. Nat. Geosci. 2016, 9, 674–678. [Google Scholar] [CrossRef]
- Lewis, S.L.; Brando, P.M.; Phillips, O.L.; van der Heijden, G.M.; Nepstad, D. The 2010 Amazon drought. Science 2011, 331, 554. [Google Scholar] [CrossRef]
- Wang, T.; Hamann, A.; Spittlehouse, D.L.; Murdock, T.Q. Climate WNA—High-resolution spatial climate data for western north America. J. Appl. Meteorol. Climatol. 2012, 51, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Wyckoff, P.H.; Clark, J.S. The relationship between growth and mortality for seven co-occurring tree species in the southern Appalachian Mountains. J. Ecol. 2002, 90, 604–615. [Google Scholar] [CrossRef] [Green Version]
- Raulier, F.; Lambert, M.; Pothier, D.; Ung, C. Impact of dominant tree dynamics on site index curves. For. Ecol. Manag. 2003, 184, 65–78. [Google Scholar] [CrossRef]
- Akinwande, M.O.; Dikko, H.G.; Samson, A. Variance Inflation Factor: As a Condition for the Inclusion of Suppressor Variable(s) in Regression Analysis. Open J. Stat. 2015, 5, 754–767. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Lingsma, H.F.; Steyerberg, E.W.; Lesaffre, E. Logistic random effects regression models: A comparison of statistical packages for binary and ordinal outcomes. BMC Med. Res. Methodol. 2011, 11, 77. [Google Scholar] [CrossRef] [Green Version]
- Spiegelhalter, D.J.; Best, N.G.; Carlin, B.P.; van der Linde, A. Bayesian measures of model complexity and fit. J. R. Stat. Soc. Ser. B 2002, 64, 583–639. [Google Scholar] [CrossRef] [Green Version]
- Hurst, J.M.; Allen, R.B.; Coomes, D.A.; Duncan, R.P. Size-specific tree mortality varies with neighbourhood crowding and disturbance in a Montane Nothofagus forest. PLoS ONE 2011, 6, e26670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 31 March 2021).
- Zweig, M.H.; Campbell, G. Receiver-operating characteristic (ROC) plots: A fundamental evaluation tool in clinical medicine. Clin. Chem. 1993, 39, 561–577. [Google Scholar] [CrossRef]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer Science & Business Media: Berlin, Germany, 2006. [Google Scholar]
- Mueller, R.C.; Scudder, C.M.; Porter, M.E.; Talbot Trotter, R., III; Gehring, C.A.; Whitham, T.G. Differential tree mortality in response to severe drought: Evidence for long-term vegetation shifts. J. Ecol. 2005, 93, 1085–1093. [Google Scholar] [CrossRef]
- Mccarthy, M.A. Bayesian Methods for Ecology; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Wunder, J.; Reineking, B.; Matter, J.F.; Bigler, C.; Bugmann, H. Predicting tree death for Fagus sylvatica and Abies alba using permanent plot data. J. Veg. Sci. 2010, 18, 525–534. [Google Scholar] [CrossRef]
- Ma, Z.; Peng, C.; Li, W.; Zhu, Q.; Wang, W.; Song, X.; Liu, J. Modeling individual tree mortality rates using marginal and random effects regression models. Nat. Resour. Modeling 2013, 26, 131–153. [Google Scholar] [CrossRef]
- Lorimer, C.G.; Frelich, L.E. A Simulation of Equilibrium Diameter Distributions of Sugar Maple (Acer saccharum). Bull. Torrey Bot. Club 1984, 111, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Chen, Q.; Sharma, R.P.; Wang, Y.; He, P.; Guo, J.; Lei, Y.; Fu, L. A climate sensitive mixed-effects diameter class mortality model for Prince Rupprecht larch (Larix gmelinii var. principis-rupprechtii) in northern China. For. Ecol. Manag. 2021, 491, 119091. [Google Scholar] [CrossRef]
- Zhang, Z.X. Dendrology (The North), 2nd ed.; China Forestry Publishing House: Beijing, China, 2010. (In Chinese) [Google Scholar]
- Buchman, R.G.; Pederson, S.P.; Walters, N.R. A tree survival model with application to species of the Great Lakes region. Can. J. For. Res. 1983, 13, 601–608. [Google Scholar] [CrossRef]
- Martínezvilalta, J.; Pinol, J. Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. For. Ecol. Manag. 2002, 161, 247–256. [Google Scholar] [CrossRef]
- Juknys, R.; Vencloviene, J.; Jurkonist, N.; Bartkevicius, E.; Sepetiene, J. Relation between Individual Tree Mortality and Tree Characteristics in a Polluted and Non-Polluted environment. Environ. Monit. Assess. 2006, 121, 519–542. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Yepez, E.A. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Cobb, N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Sun, W.; Wang, G. A climate-sensitive aboveground biomass model for three larch species in northeastern and northern China. Trees 2017, 31, 557–573. [Google Scholar] [CrossRef]
- Ricker, M.; Gutiérrez-García, G.; Daly, D.C. Modeling long-term tree growth curves in response to warming climate: Test cases from a subtropical mountain forest and a tropical rainforest in Mexico. Can. J. For. Res. 2007, 37, 977–989. [Google Scholar] [CrossRef]
- Li, Y.; Sun, W.; Zhu, H.; Zhao, X.; Bai, Y.; Zhang, Y. Multi-time scale analysis on the variations of temperature and precipitation of main urban in Northeast of China. Sci. Tech. Eng. 2015, 15, 23–31. [Google Scholar]
- Wang, X.; Zhao, C.; Jia, Q. Impacts of climate change on forest ecosystems in Northeast China. Adv. Clim. Chang. Res. 2013, 4, 230–241. [Google Scholar]
- Chen, Y.; Song, X.; Zhang, Z.; Shi, P.; Tao, F. Simulating the impact of flooding events on non-point source pollution and the effects of fifilter strips in an intensive agricultural watershed in China. Limnology 2015, 16, 91–101. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Safranyik, L. Mountain pine beetle and forest carbon feedback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variables | Meaning |
|---|---|
| MAT (°C) | Mean annual temperature |
| MWMT (°C) | Mean warmest month temperature |
| MCMT (°C) | Mean coldest month temperature |
| DT (°C) | Temperature difference between MWMT and MCMT, or continentality (°C) |
| MAP (mm) | Mean annual precipitation |
| AHM (°C) | Annual heat (MAT + 10)/(MAP/1000) |
| DD (°C) | Degree-days below 0 °C, chilling degree-days |
| DD5 (°C) | Degree-days above 5 °C, growing degree-days |
| DD_18 (°C) | Degree-days below 18 °C, heating degree-days |
| DD18 (°C) | Degree-days above 18 °C, cooling degree-days |
| NFFD | The number of frost-free days |
| PAS | Precipitation as snow (mm) between August in previous year and July in current year |
| EMT | Extreme minimum temperature over 30 years |
| EXT | Extreme maximum temperature over 30 years |
| Eref | Hargreaves reference evaporation |
| CMD | Hargreaves climatic moisture deficit |
| Scales | Variables | Min | Max | Mean | Std |
|---|---|---|---|---|---|
| individual tree variables (I) | D (cm) | 5.00 | 67.00 | 20.92 | 10.08 |
| H (m) | 1.80 | 37.80 | 16.52 | 7.22 | |
| DL (cm) | 0 | 4.28 | 1.52 | 0.79 | |
| RD | 0.16 | 2.93 | 0.95 | 0.43 | |
| Stand variables (S) | N | 250.00 | 2875.00 | 1233.00 | 555.47 |
| DH (m) | 15.86 | 30.72 | 23.08 | 3.12 | |
| SMD (cm) | 12.69 | 33.44 | 23.03 | 3.97 | |
| RSI | 0.48 | 1.59 | 0.77 | 0.17 | |
| Climate variables (C) | MAT (°C) | 0.90 | 4.40 | 3.20 | 1.29 |
| MWMT (°C) | 14.60 | 17.60 | 16.52 | 0.98 | |
| MCMT (°C) | −14.20 | −10.70 | −12.02 | 1.33 | |
| DT (°C) | 28.30 | 29.00 | 28.52 | 0.33 | |
| MAP (mm) | 574.00 | 789.00 | 583.00 | 77.41 | |
| AHM (°C) | 14.20 | 25.10 | 20.97 | 4.17 | |
| DD (°C) | 1023.00 | 1515.00 | 1194.00 | 182.19 | |
| DD5 (°C) | 994.00 | 1575.00 | 1368.00 | 205.17 | |
| DD_18 (°C) | 4917.00 | 6123.00 | 5336.00 | 441.26 | |
| DD18 (°C) | 11.00 | 53.00 | 34.09 | 14.21 | |
| NFFD | 152.00 | 173.00 | 164.10 | 6.87 | |
| PAS | 41.00 | 140.00 | 69.74 | 38.71 | |
| EMT | −28.60 | −26.30 | −27.33 | 0.86 | |
| EXT | 25.20 | 28.80 | 27.64 | 1.26 | |
| Eref | 540.00 | 719.00 | 664.50 | 70.86 | |
| CMD | 44.00 | 199.00 | 139.40 | 62.70 |
| Model | Equation | AUC | Threshold |
|---|---|---|---|
| I | 1.7026 − 1.4113DL + 7.1936RD | 0.832 | 0.908 |
| S | 29.6056 + 0.4059SMD − 0.8952DH | 0.604 | 0.885 |
| C | −259.3 + 10.49DT − 0.0039MAP | 0.604 | 0.885 |
| I + S | −0.0514 − 0.9608DL + 4.4935RD + 0.0529SMD | 0.831 | 0.889 |
| I + C | 3.1002 − 1.2071DL + 6.1207RD − 0.0025MAP | 0.832 | 0.901 |
| S + C | −437.1 + 0.8300SMD − 1.009DH + 17.78DT − 0.0638MAP | 0.603 | 0.882 |
| I + S + C | −0.8265 − 1.104DL + 6.611RD + 0.1167SMD + 3.118DT − 0.0012MAP | 0.832 | 0.894 |
| Model | Form | I | S | C | Variance Component | Thres-Hold | AUC | DIC | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Intercept | DL | RD | SMD | D T | MAP | Block | Plot | |||||
| I | base | 1.703 *** | −1.411 *** | 7.194 *** | 0.908 | 0.828 | 1498.87 | |||||
| One-level | 1.409 ** | −1.119 *** | 5.214 *** | 1.003 | 0.832 | 1479.14 | ||||||
| Two-level | 1.727 * | −1.409 *** | −1.119 *** | 2.335 | 0.966 | 0.830 | 1445.76 | |||||
| I + S | base | −0.051 | −0.961 *** | 4.494 *** | 0.053 *** | 0.889 | 0.831 | 1598.87 | ||||
| One-level | 0.175 | −1.115 *** | 4.782 *** | 0.058 . | 0.843 | 0.832 | 1472.02 | |||||
| Two-level | −0.608 | −1.055 *** | 5.501 **** | 0.092 * | 1.368 | 0.577 | 0.830 | 1463.21 | ||||
| I + C | base | 3.100 *** | −1.207 *** | 6.121 *** | −0.002 * | 0.901 | 0.832 | 1580.79 | ||||
| One-level | 3.617 ** | −1.243 *** | 5.584 *** | −0.003 . | 1.126 | 0.830 | 1469.72 | |||||
| Two-level | 6.844 | −1.104 *** | 6.319 *** | −0.008 | 2.472 | 0.832 | 0.826 | 1439.77 | ||||
| I + S + C | base | −82.65 *** | −1.104 *** | 6.611 *** | 0.117 *** | 3.118 *** | 0.012 *** | 0.894 | 0.832 | 1544.29 | ||
| One-level | −63.36 | −1.224 *** | 6.178 *** | 0.102 . | 2.424 | −0.011 . | 1.215 | 0.830 | 1455.61 | |||
| Two-level | −16.81 . | −0.944 *** | 6.36 *** | 0.1187 * | 6.420 . | −0.026 * | 3.879 | 0.626 | 0.834 | 1441.67 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, L.; Chen, X.; Zhou, X.; Sharma, R.P.; Li, J. Developing Tree Mortality Models Using Bayesian Modeling Approach. Forests 2022, 13, 604. https://doi.org/10.3390/f13040604
Xie L, Chen X, Zhou X, Sharma RP, Li J. Developing Tree Mortality Models Using Bayesian Modeling Approach. Forests. 2022; 13(4):604. https://doi.org/10.3390/f13040604
Chicago/Turabian StyleXie, Lu, Xingjing Chen, Xiao Zhou, Ram P. Sharma, and Jianjun Li. 2022. "Developing Tree Mortality Models Using Bayesian Modeling Approach" Forests 13, no. 4: 604. https://doi.org/10.3390/f13040604
APA StyleXie, L., Chen, X., Zhou, X., Sharma, R. P., & Li, J. (2022). Developing Tree Mortality Models Using Bayesian Modeling Approach. Forests, 13(4), 604. https://doi.org/10.3390/f13040604