
Climatic and Anthropogenic Drivers of Forest Succession in the Iberian Pyrenees during the Last 500 Years: A Statistical Approach


Abstract
:1. Introduction
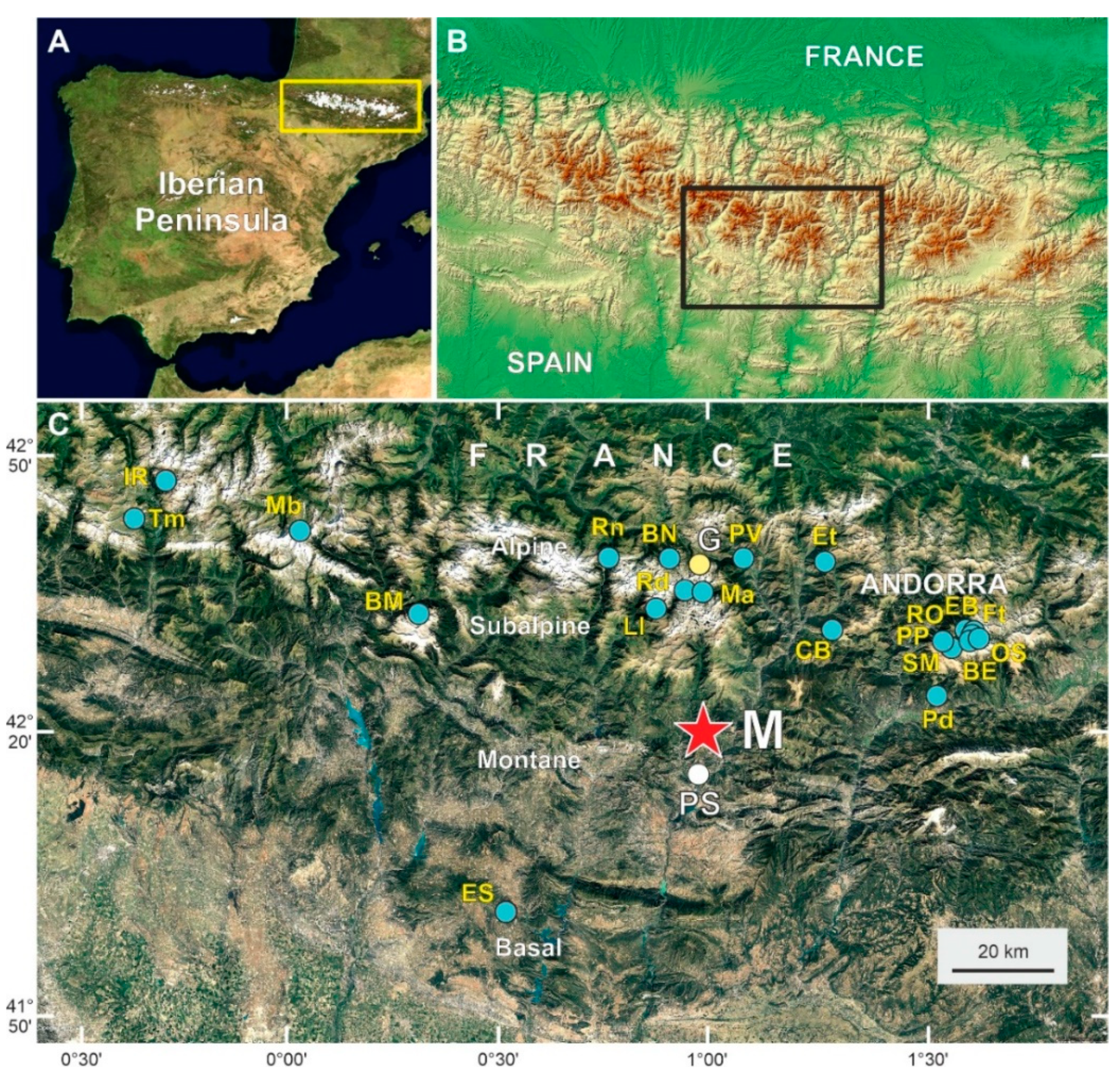
2. Study Site
2.1. Location and Climate
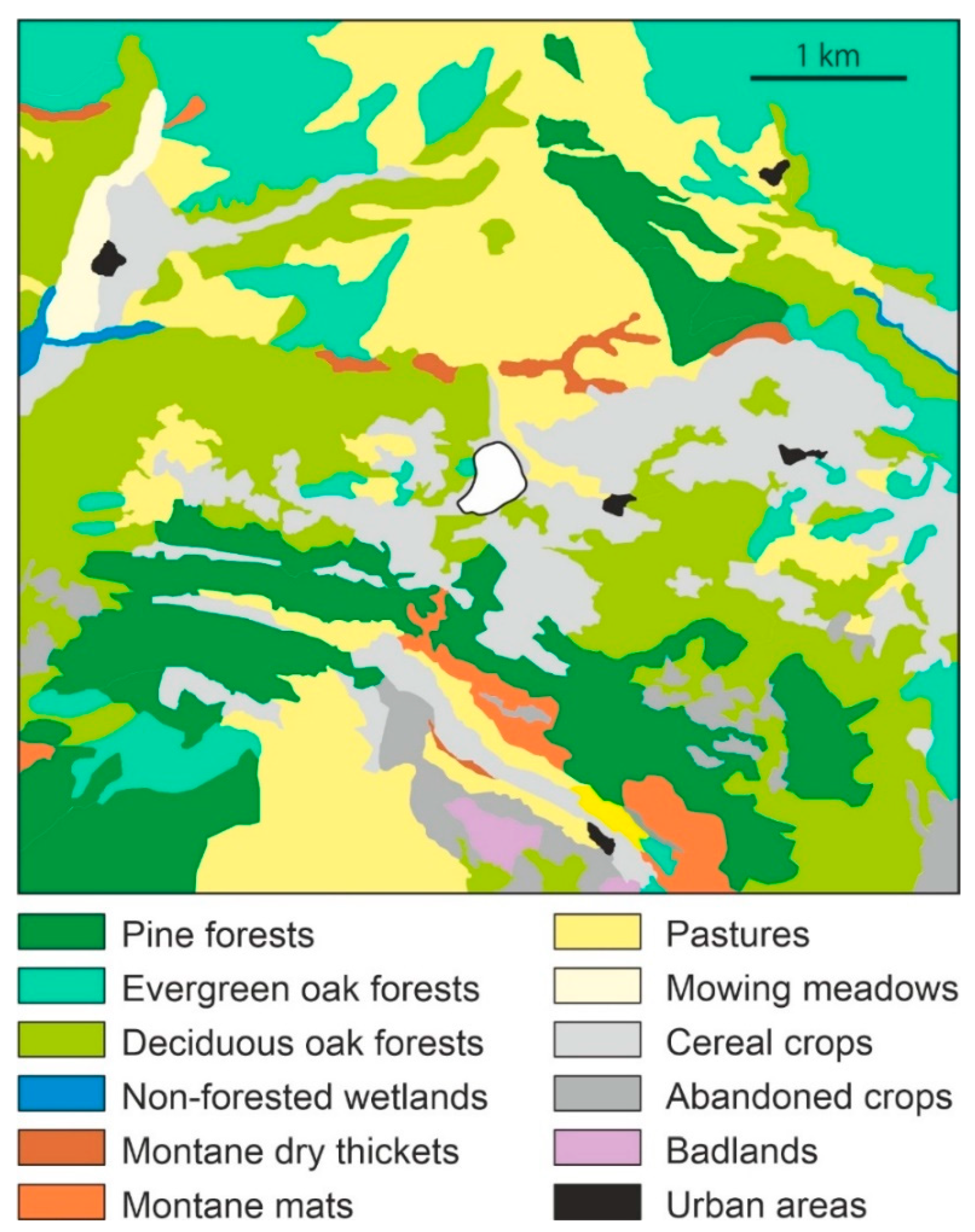
2.2. Regional Vegetation
3. Material and Methods
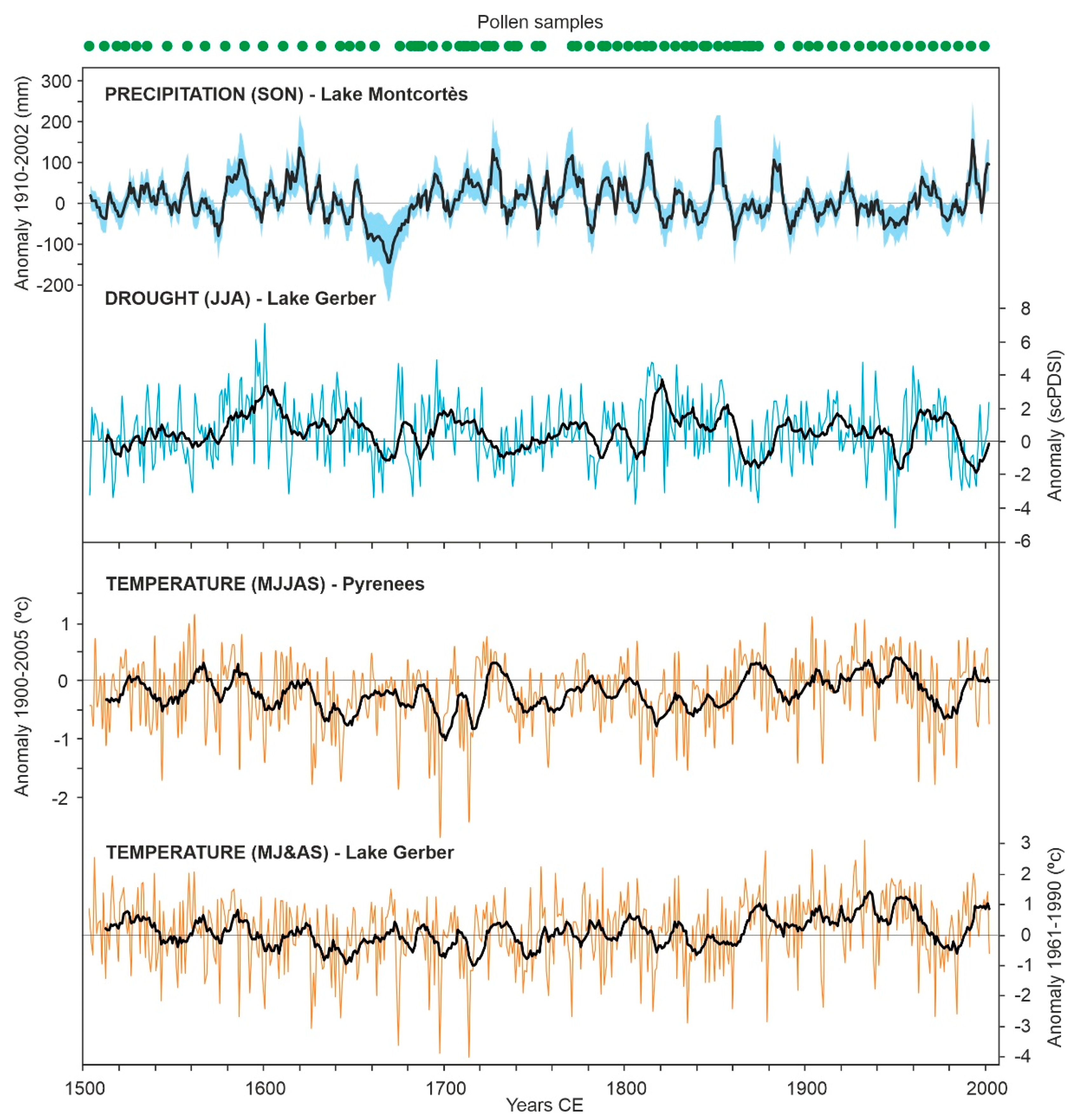
3.1. Raw Data
3.2. Statistical Methods
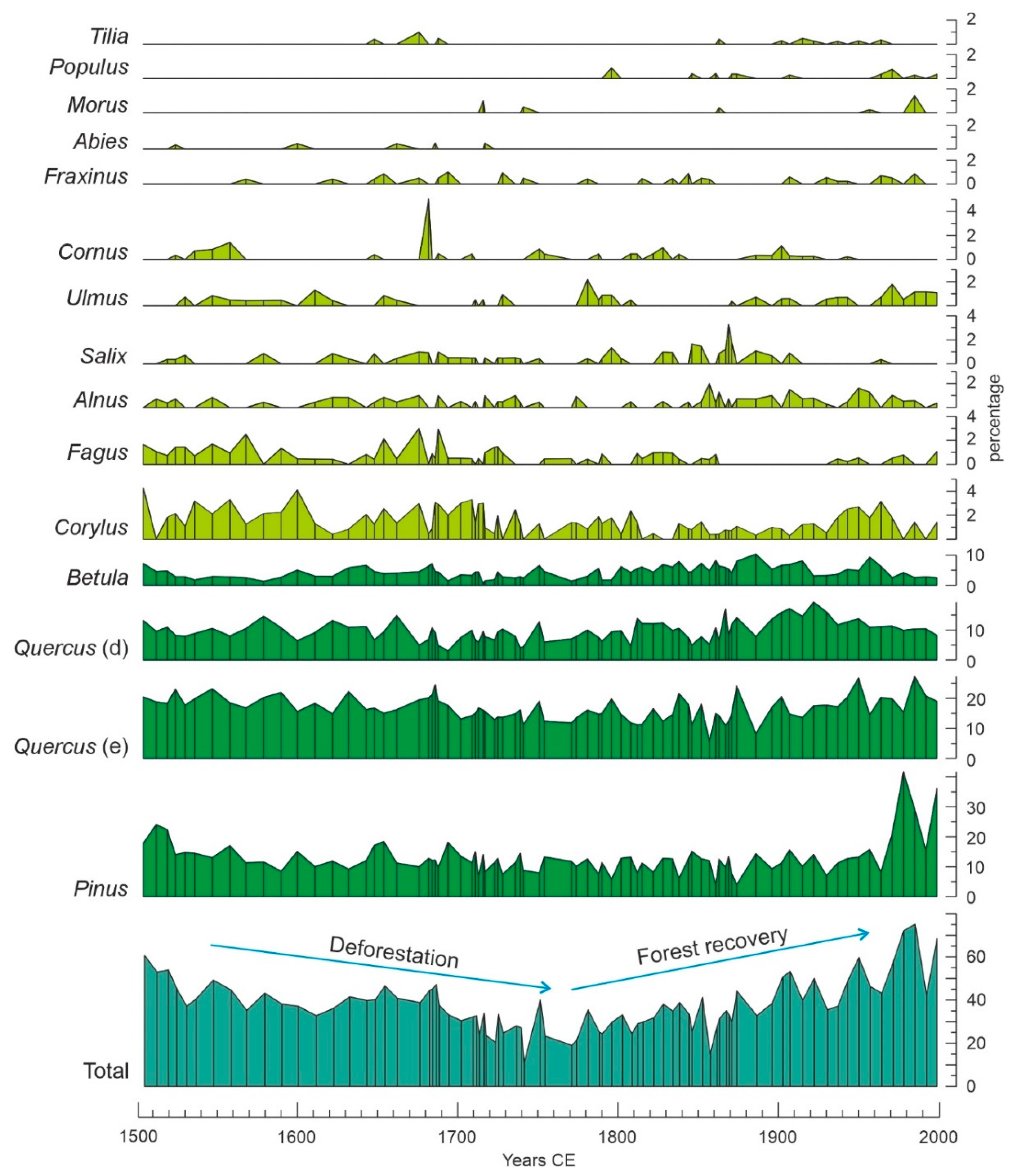
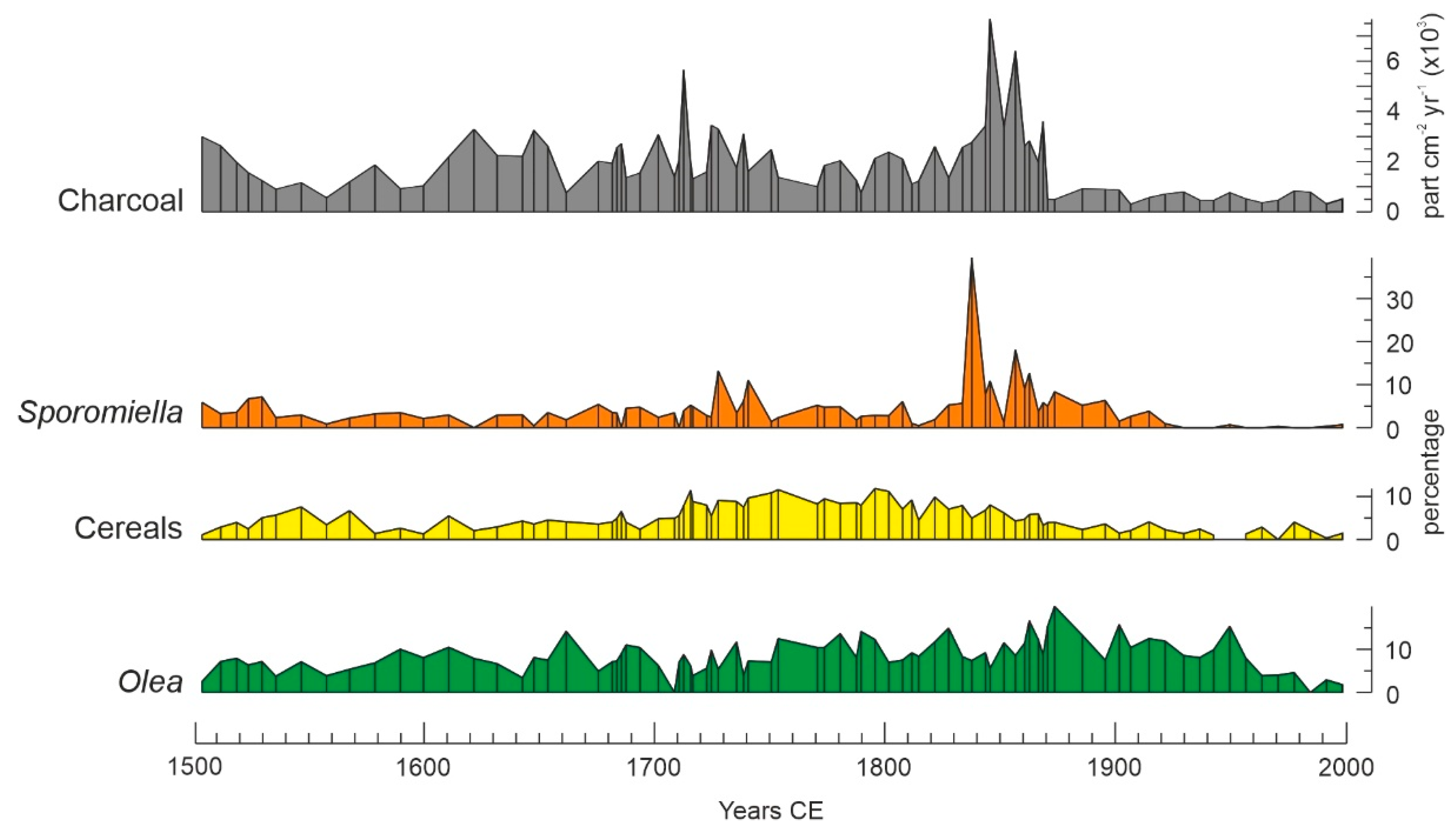
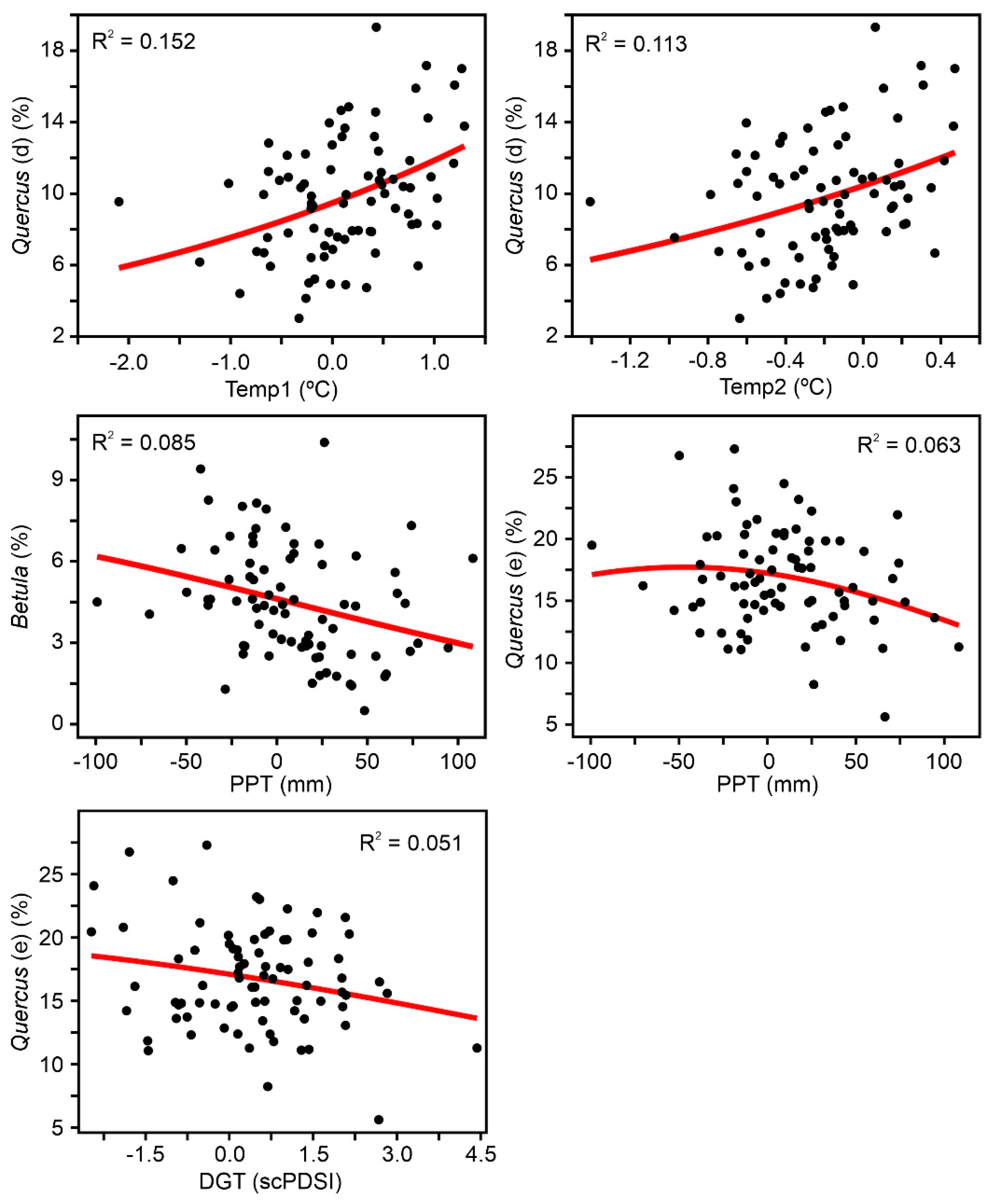
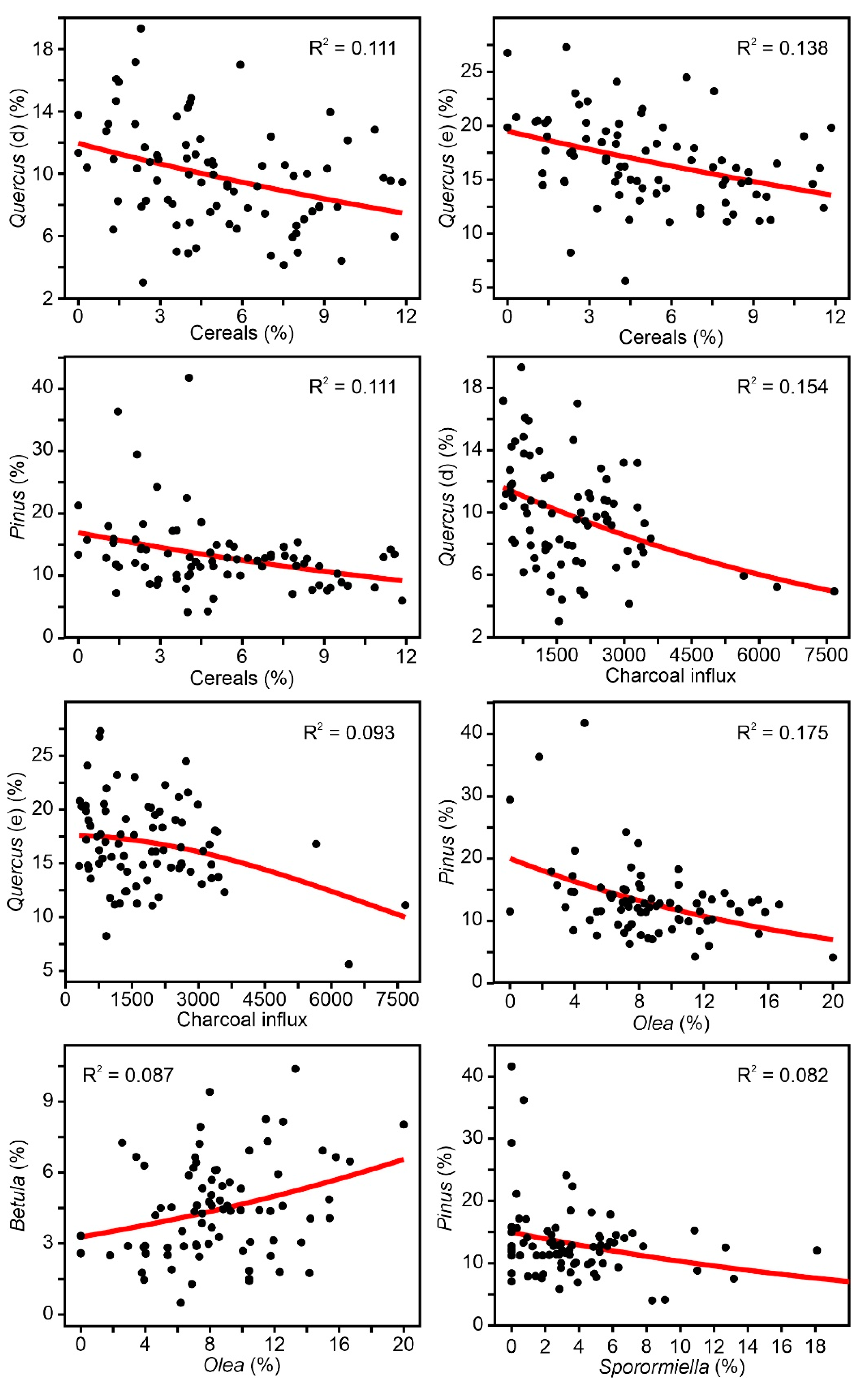
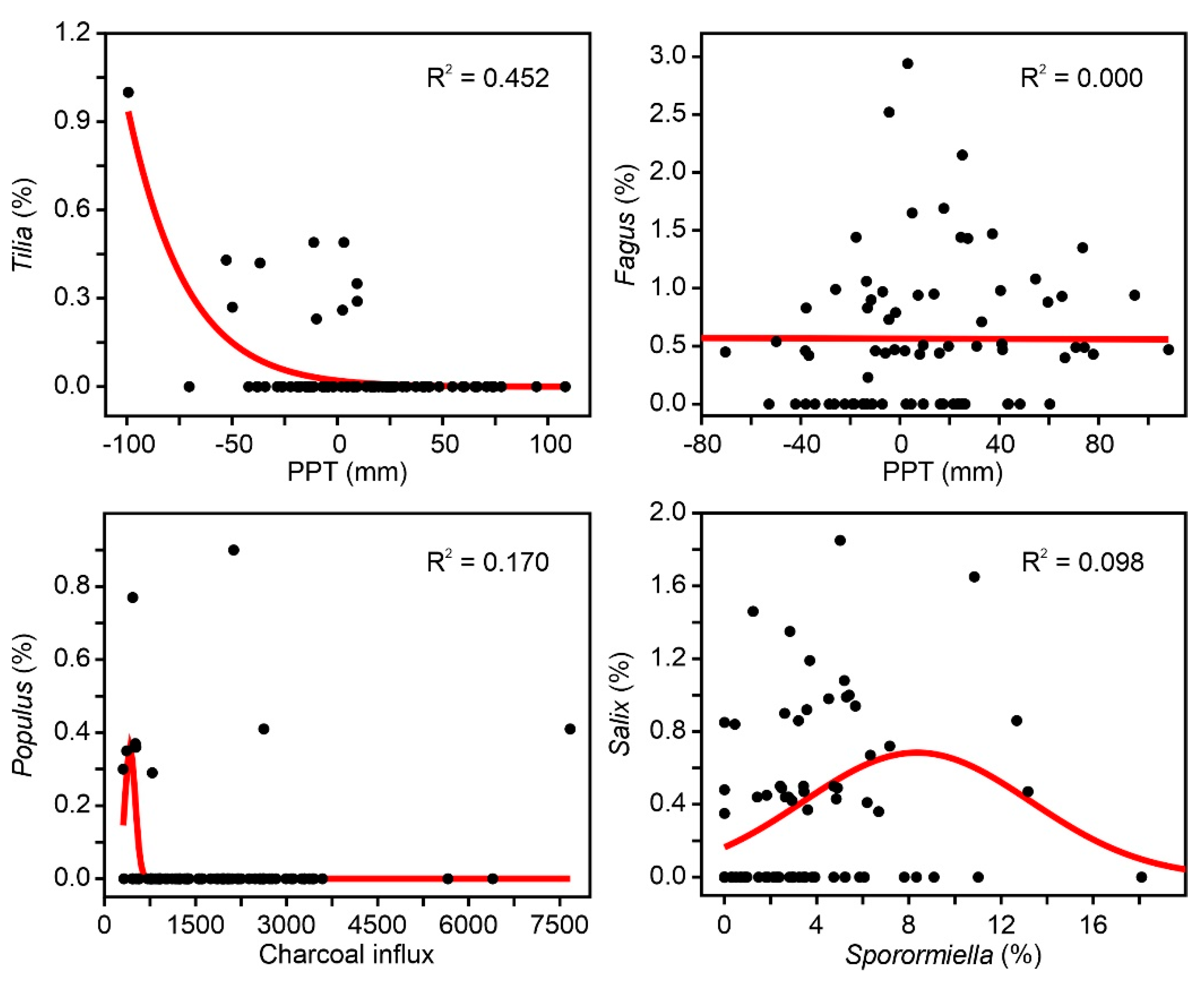
4. Results
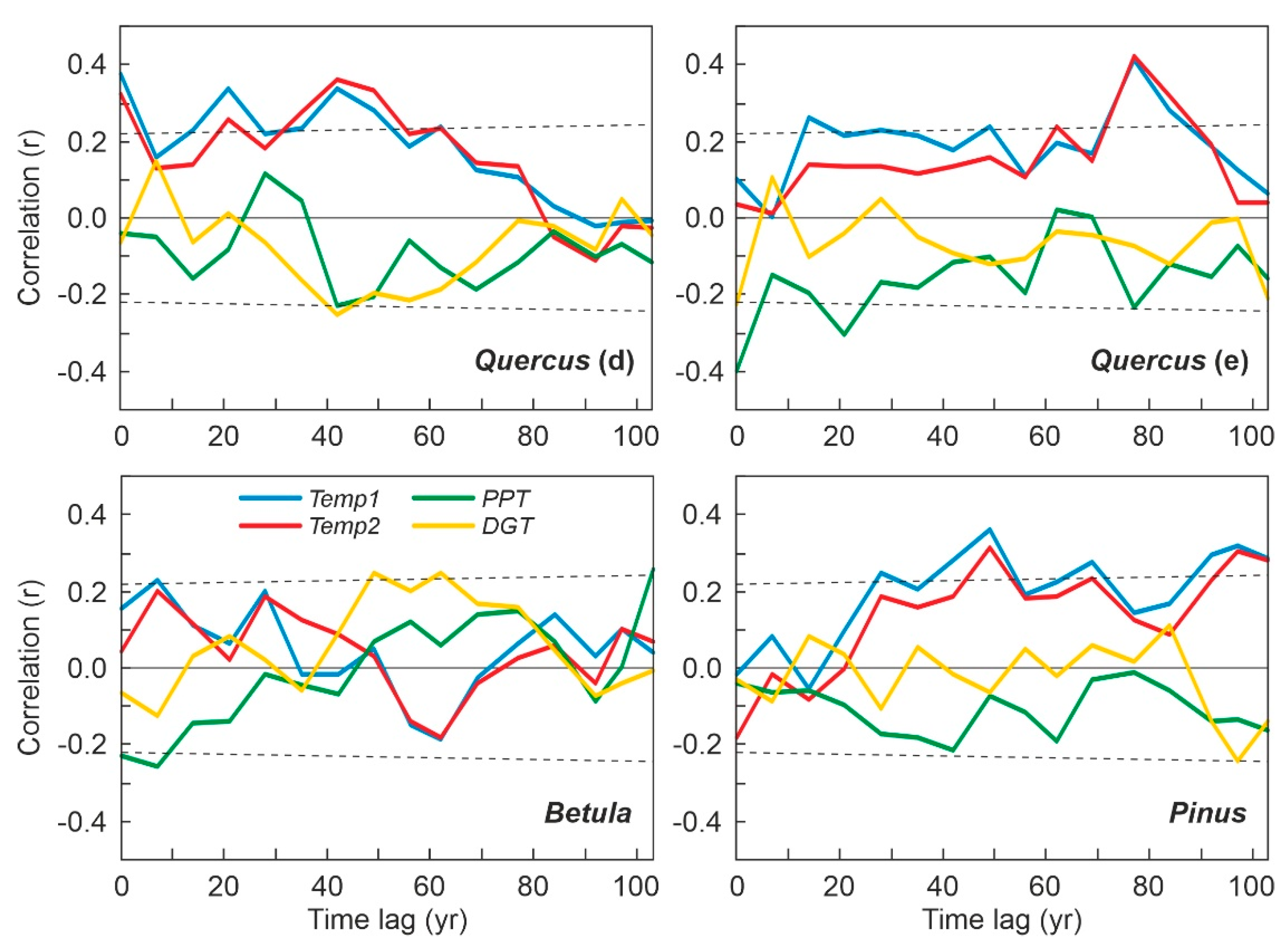
4.1. Individual Responses of Tree Taxa to External Drivers
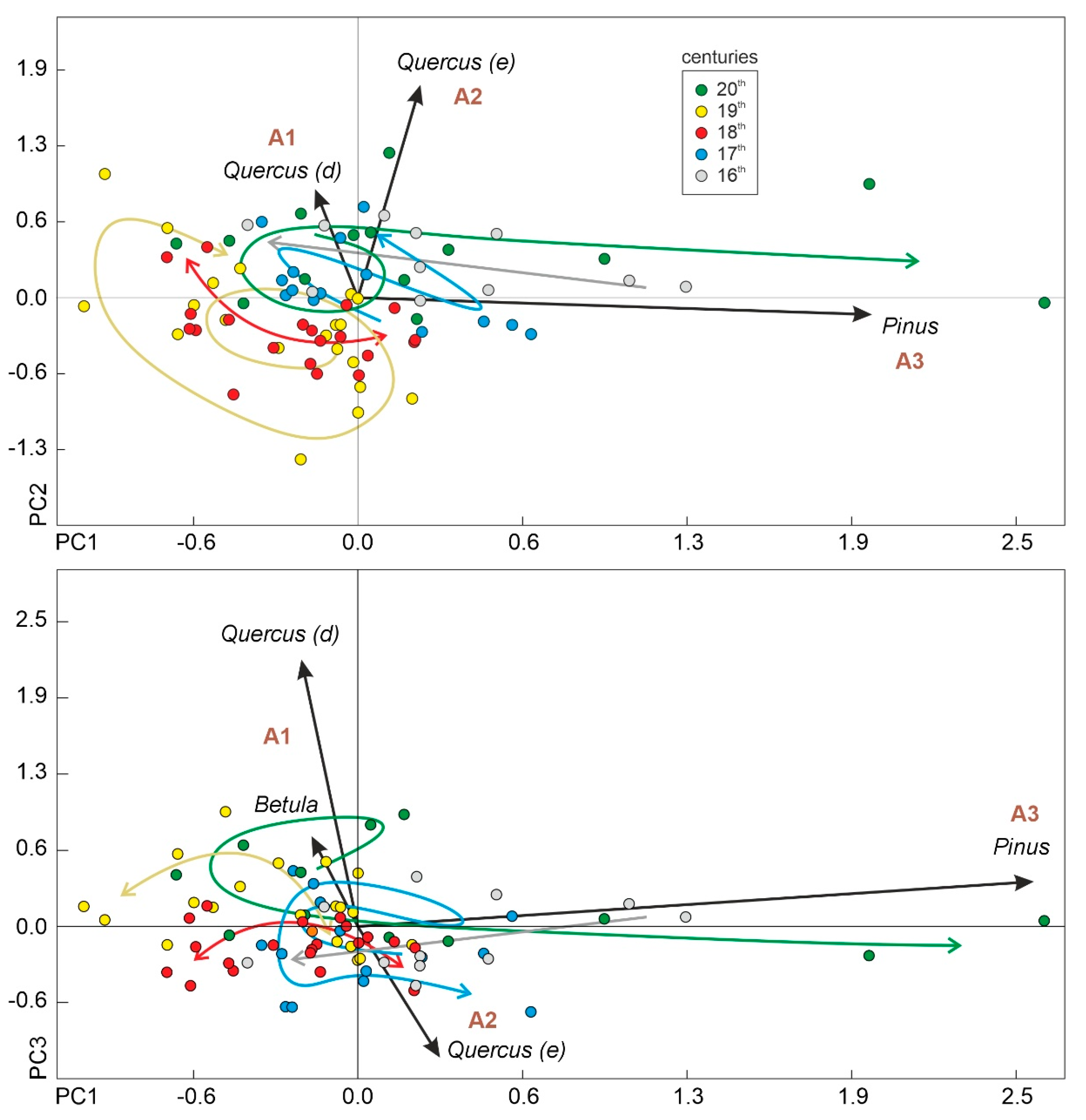
4.2. Forest Assemblages and Rate-of-Change Analysis
4.3. Successional Trends
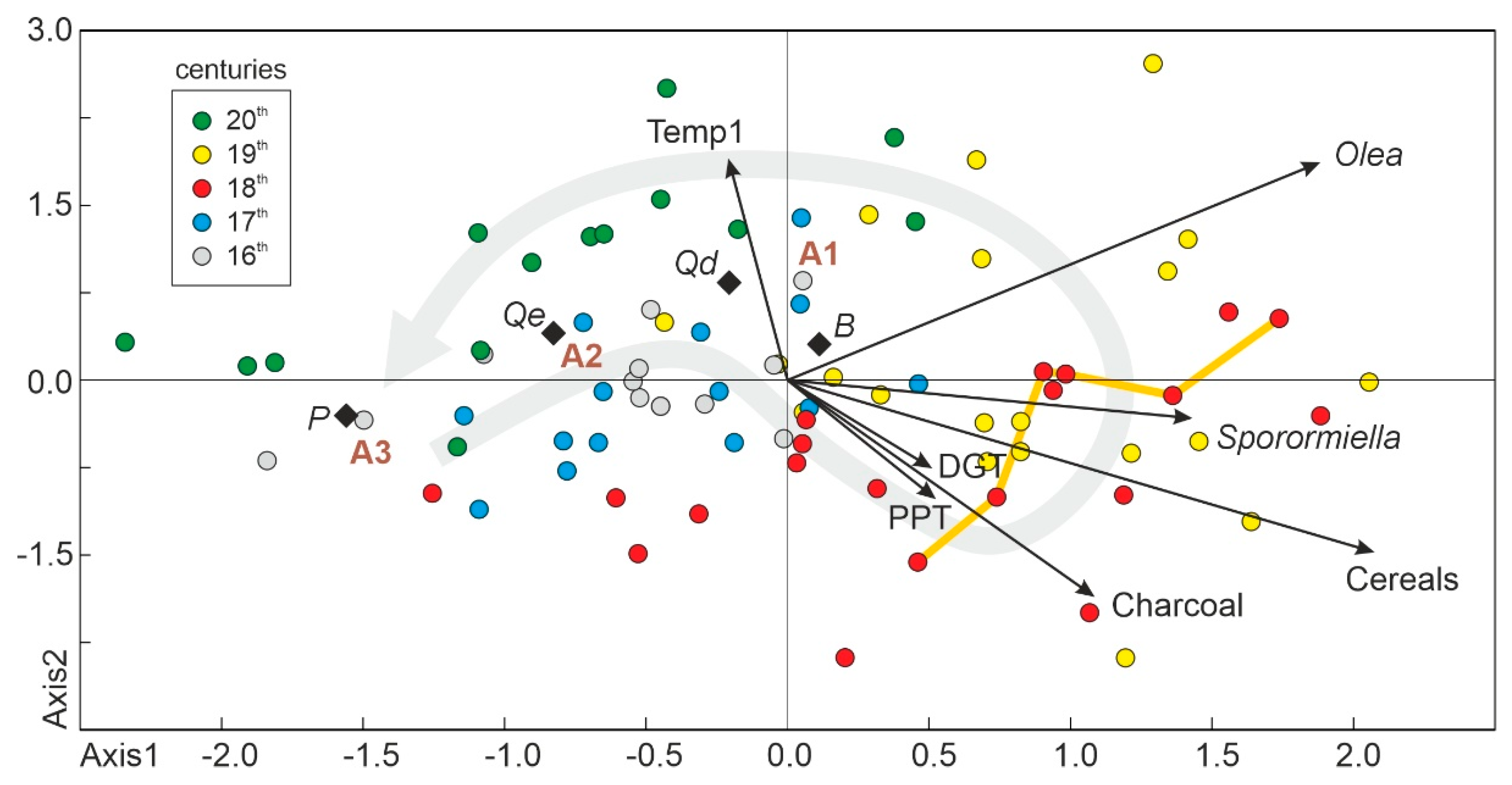
4.4. Synthetic Successional-Environmental Analysis
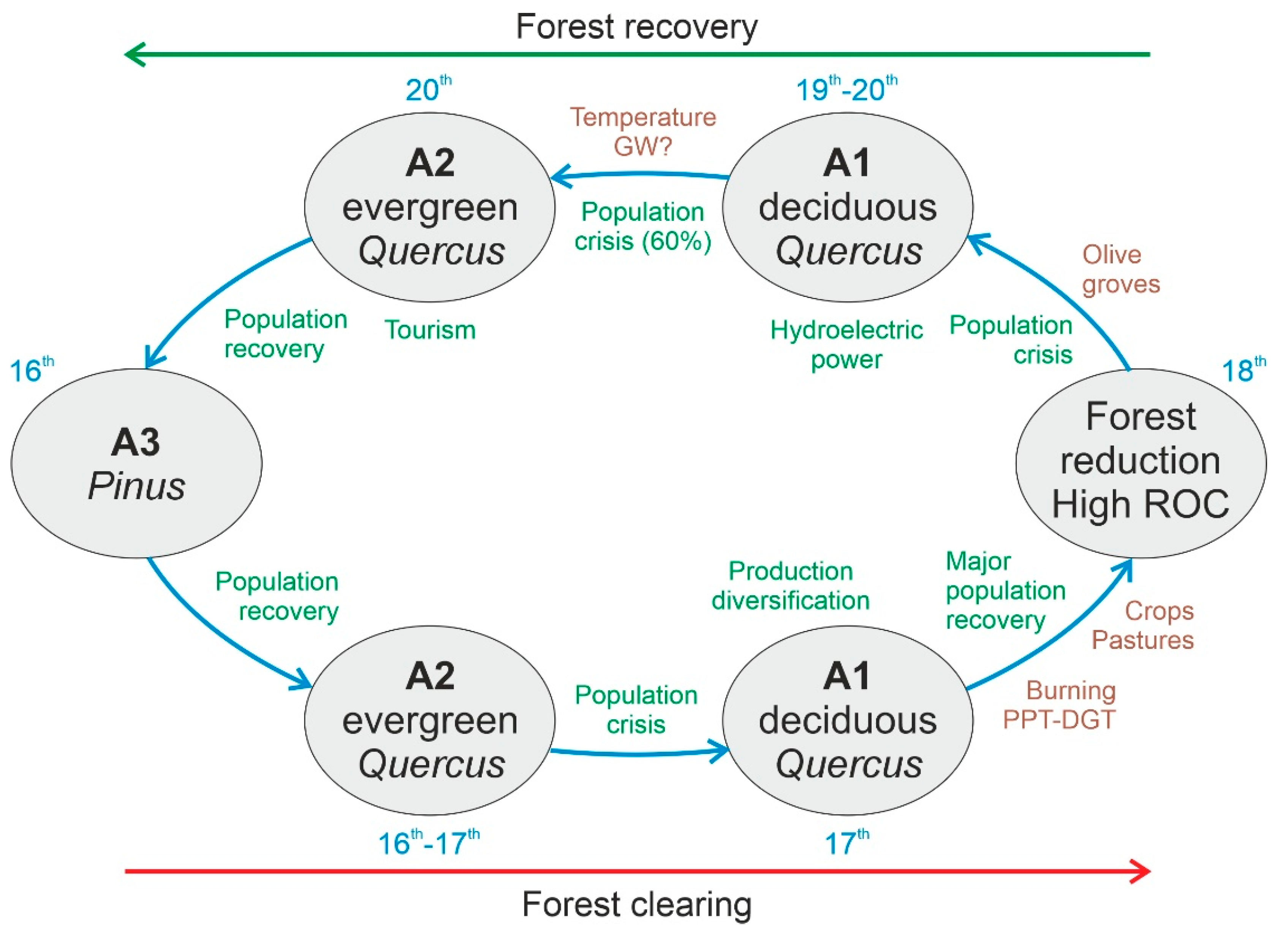
5. Discussion and Conclusions
6. Final Remarks
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falk, D.A.; van Mantgem, P.J.; Keeley, J.E.; Gregg, R.N.; Guiterman, C.H.; Tepley, A.J.; Young, D.; Marshall, L.A. Mechanisms of forest resilience. For. Ecol. Manag. 2022, 512, 120129. [Google Scholar] [CrossRef]
- Trugman, A. Integrating plant physiology and community ecology across scales through a trait-based models to predict drought mortality. New Phytol. 2022, 234, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Yi, C.; Hendrey, G.; Niu, S.; McDowell, N.; Allen, C.D. Tree mortality in a warming world: Causes, patterns, and implications. Environ. Res. Lett. 2022, 17, 030201. [Google Scholar] [CrossRef]
- Willis, K.J.; Bailey, R.M.; Bhagwat, S.A.; Birks, H.J.B. Biodiversity baselines, thresholds and resilience: Testing predictions and assumptions using palaeoecological data. Trends Ecol. Evol. 2010, 25, 583–591. [Google Scholar] [CrossRef]
- Willis, K.J.; Birks, H.J.B. What is natural? The need for a long-term perspective in biodiversity conservation. Science 2006, 314, 1261–1265. [Google Scholar] [CrossRef] [Green Version]
- Vegas-Vilarrúbia, T.; Rull, V.; Montoya, E.; Safont, E. Quaternary palaeoecology and nature conservation: A general review with emphasis on the Neotropics. Quat. Sci. Rev. 2011, 30, 2361–2388. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T. What is long term in ecology? Trends Ecol. Evol. 2011, 26, 3–4. [Google Scholar] [CrossRef]
- Smol, J.P.; Last, W.L. (Eds.) Tracking Environmental Change Using Lake Sediments; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Dubois, N.; Jacob, J. Molecular biomarkers of anthropogenic impacts in natural archives: A review. Front. Ecol. Evol. 2016, 4, 92. [Google Scholar] [CrossRef] [Green Version]
- Castaneda, I.S.; Schouten, S. A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quat. Sci. Rev. 2011, 30, 2851–2891. [Google Scholar] [CrossRef]
- Leunda, M.; González-Sampériz, P.; Gil-Romera, G.; Aranbarri, J.; Moreno, A.; Oliva-Urcia, B.; Sevilla-Callejo, M.; Valero-Gracés, B. The Late-Glacial and Holocene Marboré Lake sequence (2612 m a.s.l., Central Pyrenees, Spain): Testing high altitude sites sensitivity to millennial scale vegetation and climate variability. Glob. Planet. Chang. 2017, 157, 214–231. [Google Scholar] [CrossRef] [Green Version]
- Ejarque, A. Génesis y Configuración Microregional de un Paisaje Cultural Pirenaico de alta Montaña Durante el Holoceno: Estudio Polínico y de Otros Indicadores Paleoambientales en el Valle de Madriu-Perafita-Claror (Andorra). Ph.D. Thesis, University Rovira i Virgili, Tarragona, Spain, 2009. [Google Scholar]
- Miras, Y.; Ejarque, A.; Riera, S.; Orengo, H.A.; Palet, J.M. Andorran high Pyrenees (Perafiya Valley, Andorra): Serra Mitjana fen. Grana 2015, 54, 313–316. [Google Scholar] [CrossRef] [Green Version]
- Ejarque, A.; Miras, Y.; Riera, S.; Palet, J.M.; Orengo, H.A. Testing micro-regional variability in the Holocene shaping of high mountain cultural landscapes: A palaeoenvironmental case-study in the eastern Pyrenees. J. Archaeol. Sci. 2010, 37, 1468–1479. [Google Scholar] [CrossRef] [Green Version]
- Cunill, R.; Soriano, J.M.; Bal, M.C.; Pèlachs, A.; Rodríguez, J.M.; Pérez-Obiol, R. Holocene high-altitude vegetation dynamics in the Pyrenees: A pedoanthracology contribution to an interdisciplinary appraoch. Quat. Int. 2013, 289, 60–70. [Google Scholar] [CrossRef]
- Pla, S.; Catalan, J. Chrysophyte cysts from lake sediments reveal the submillennial winter/spring climate variability in the northwestern Mediterranean region throughout the Holocene. Clim. Dyn. 2005, 24, 263–278. [Google Scholar] [CrossRef]
- Catalan, J.; Pla-Rabés, S.; García, J.; Camarero, L. Air temperature-driven CO2 consumption by rock weathering at short timescales: Evidence from a Holocene lake sediment record. Geoch. Cosmoch. Acta 2014, 136, 67–79. [Google Scholar] [CrossRef]
- Miras, Y.; Ejarque, A.; Riera, S.; Palet, J.M.; Orengo, H.; Euba, I. Holocene vegetation changes and land-use history in the Andorran Pyrenees since the Early Neolithic: The pollen record of Bosc dels estanyons (2180 m a.s.l., Vall del Madriu, Andorra). Comptes Rendus Palevol 2007, 6, 291–300. [Google Scholar] [CrossRef] [Green Version]
- Montserrat Martí, J.M. Evolución Glaciar y Postglaciar del Clima y la Vegetación en la Vertiente sur del Pirineo: Estudio Palinológico; Monografías del Instituto Pirenaico de Ecología: Zaragoza, Spain, 1992. [Google Scholar]
- Ejarque, A.; Julià, R.; Riera, S.; Palet, J.M.; Orengo, H.A.; Miras, Y. Tracing the history of highland human management in the eastern Pre-Pyrenees: An interdisciplinary palaeoenvironmental study at the Pradell fen, Spain. Holocene 2009, 19, 1241–1255. [Google Scholar] [CrossRef] [Green Version]
- Rull, V.; Cañellas-Boltà, N.; Vegas-Vilarrúbia, T. Late-Holocene forest resilience in the central Pyrenean highlands as deduced from pollen analysis Lake Sant Maurici sediments. Holocene 2021, 31, 1797–1803. [Google Scholar] [CrossRef]
- Pérez-Sanz, A.; González-Sampériz, P.; Moreno, A.; Valero-Garcés, B.; Gil-Romera, G.; Rieradevall, M.; Tarrats, P.; Lasheras-Álvarez, L.; Morellón, M.; Belmonte, A.; et al. Holocene climatic variability, vegetation dynamics and fire regime in the central Pyrenees: The basa de la Mora sequence (NE Spain). Quat. Sci. Rev. 2013, 73, 149–169. [Google Scholar] [CrossRef] [Green Version]
- Garcés-Pastor, S.; Cañellas-Boltà, N.; Clavaguera, A.; Calero, M.A.; Vegas-Vilarrúbia, T. Vegetation shifts, human impact and peat bog development in Bassa Nera pond (Central Pyrenees) during the last millennium. Holocene 2016, 27, 553–565. [Google Scholar] [CrossRef]
- Garcés-Pastor, S.; Cañellas-Boltà, N.; Pèlachs, A.; Soriano, J.M.; Pérez-Obiol, R.; Pérez-Hasse, A.; Calero, M.A.; Andreu, O.; Escolà, N.; Vegas-Vilarrúbia, T. Environmental history and vegetation dynamics in response to climate variations and human pressure during the Holocene in Bassa Nera, Central Pyrenees. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 479, 48–60. [Google Scholar] [CrossRef]
- Pèlachs, A.; Soriano, J.M.; Nadal, J.; Esteban, A. Holocene environmental history and human impact in the Pyrenees. Contrib. Sci. 2007, 3, 421–429. [Google Scholar]
- Catalan, J.; Pèlachs, A.; Gassiot, E.; Antolín, F.; Ballesteros, A.; Batalla, M.; Burjachs, F.; Buchaca, T.; Camarero, L.; Clemente, I.; et al. Interacción entre clima y ocupación humana en la configuración del paisaje vegetal del Parque nacional de Aigüestortes i Estany de sant Maurici a lo largo de los últimos 15.000 años. Proy. Investig. Parq. Nac. 2013, 71–92. [Google Scholar]
- Pèlachs, A.; Pérez-Obiol, R.; Ninyerola, M.; Nadal, J. Landscape dynamics of Abies and fagus in the southern Pyrenees during the last 2200 years as a result of anthropogenic impacts. Rev. Palaeobot. Palynol. 2009, 156, 337–349. [Google Scholar] [CrossRef]
- Riera, S.; Wansard, G.; Julià, R. 2000-year environmental history of a karstic lake in the Mediterranean Pre-Pyrenees: The Estanya lakes (Spain). Catena 2004, 55, 293–324. [Google Scholar] [CrossRef]
- González-Sampériz, P.; Aranbarri, J.; Pérez-Sanz, A.; Gil-Romera, G.; Moreno, A.; Leunda, M.; SEvilla-Callejo, M.; Corella, J.P.; Morellón, M.; Oliva, B.; et al. Environmental and climate change in the southern Central Pyrenees since the Last Glacial Maximum: A review from lake records. Catena 2017, 149, 668–688. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T. Conifer Forest Dynamics in the Iberian Pyrenees during the Middle Ages. Forests 2021, 12, 1685. [Google Scholar] [CrossRef]
- González-Sampériz, P.; Montes, L.; Aranbarri, J.; Leunda, M.; Domingo, R.; Laborda, R.; Sanjuan, Y.; Gil-Romera, G.; Lasanta, T.; García-Ruiz, J.M. Escenarios, tiempo e indicadores paleoambientales para la identificación del Antropoceno en el paisaje vegetal del Pirineo central (NE Iberia). Cuad. Investig. Geogr. 2019, 45, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Rull, V.; Vegas-Vilarrúbia, T. A spatiotemporal gradient in the anthropization of Pyrenean landscapes. Preliminary report. Quat. Sci. Rev. 2021, 258, 106909. [Google Scholar] [CrossRef]
- Rull, V. Quaternary Ecology, Evolution and Biogeography; Elsevier: Amsterdam, The Netherlands; Academic Press: London, UK, 2020. [Google Scholar]
- Zolitschka, B.; Francus, P.; Ojala, A.E.K. Varves in lake sediments—A review. Quat. Sci. Rev. 2015, 1171, 1–41. [Google Scholar] [CrossRef]
- Corella, J.P.; Valero-Garcés, B.L.; Vicente-Serrano, S.M.; Brauer, A.; Benito, G. Three millennia of heavy rainfalls in Western Mediterranean: Frequency, seasonality and atmospheric drivers. Sci. Rep. 2016, 6, 38206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corella, J.P.; Benito, G.; Wilhelm, B.; Montoya, E.; Rull, V.; Vegas-Vilarrúbia, T.; Valero-Garcés, B.L. A millennium-long perspective of flood-related seasonal sediment yield in Mediterranean watersheds. Glob. Planet. Change 2019, 177, 127–140. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Trapote, M.C.; Montoya, E.; Valero-Garcés, B. A unique Pyrenean varved record provides a detailed reconstruction of Mediterranean vegetation and land-use dynamics over the last three millennia. Quat. Sci. Rev. 2021, 268, 107128. [Google Scholar] [CrossRef]
- Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Valero-Garcés, B. Bronze Age to Medieval vegetation dynamics and landscape anthropization in the southern-central Pyrenees. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 571, 110392. [Google Scholar] [CrossRef]
- Trapote, M.C.; Rull, V.; Giralt, S.; Corella, J.P.; Montoya, E.; Vegas-Vilarrúbia, T. High-resolution (sub-decadal) pollen analysis of varved sediments from Lake Montcortès (southern Pyrenean flank): A fine-tuned record of landscape dynamics and human impact during the last 500 years. Rev. Palaeobot. Palynol. 2018, 259, 207–222. [Google Scholar] [CrossRef] [Green Version]
- Rull, V.; González-Sampériz, P.; Corella, J.P.; Morellón, M.; Giralt, S. Vegetation changes in the southern Pyrenean flank during the last millennium in relation to climate and human activities: The Montcortès lacustrine record. J. Paleolimnol. 2011, 46, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Rull, V.; Vegas-Vilarrúbia, T. Crops and weeds from the Estany de Montcortès catchment, central Pyrenees, during the last millennium: A comparison of palynological and historical records. Veg. Hist. Archaeobot. 2015, 24, 699–710. [Google Scholar] [CrossRef]
- Montoya, E.; Rull, V.; Vegas-Vilarrúbia, T.; Corella, J.P.; Giralt, S.; Valero-Garcés, B. Grazing activities in the southern central Pyrenees during the last millennium as deduced from the non-pollen palynomorphs (NPP) record of Lake Montcortès. Rev. Palaeobot. Palynol. 2018, 254, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Dorado-Liñán, I.; Büntgen, U.; González-Rouco, F.; Zorita, E.; Montávez, J.P.; Gómez-Navarro, J.J.; Brunet, M.; Heinrich, I.; Helle, G.; Gutiérrez, E. Estimating 750 years of temperature variations and uncertainties in the Pyrenees by tree-ring reconstructions and climate simulations. Clim. Past 2012, 8, 919–933. [Google Scholar] [CrossRef] [Green Version]
- Büntgen, U.; Krusic, P.J.; Verstege, A.; Sangüesa-Barreda, G.; Wagner, S.; Camarero, J.J.; Ljungqvist, F.C.; Zorita, E.; Oppenheimer, C.; Konter, O.; et al. New tree-ring evidence from the Pyrenees reveals western Mediterranean climate variability since Medieval times. J. Clim. 2017, 30, 5295–5318. [Google Scholar] [CrossRef]
- Corella, J.P.; Benito, G.; Rodriguez-Lloveras, X.; Brauer, A.; Valero-Garcés, B.L. Annually-resolved lake record of extreme hydrometeorological events since AD 1347 in NE Iberian Peninsula. Quat. Sci. Rev. 2014, 93, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Vegas-Vilarrúbia, T.; Corella, J.P.; Sigró, J.; Rull, V.; Dorado-Liñán, I.; Valero-Garcés, B.L.; Gutiérrez, E. Regional precipitation trends since 1500 CE, as reconstructed from calcite sublayers of a varved Mediterranean lake record (central Pyrenees). Sci. Total Environ. 2022, 826, 153773. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.M.; Foster, B.L.; Ballanthyne, F.; Collins, C.D.; Antonovics, J.; Holt, R.D. Metapopulations and metacommunities: Combining spatial and temporal perspectives in plant ecology. J. Ecol. 2012, 100, 88–103. [Google Scholar] [CrossRef]
- Holyoak, M.; Leibold, M.A.; Holt, R.D. Metacommunities: Spatial Dynamics and Ecological Communities; Chicago University Press: Chicago, IL, USA, 2005. [Google Scholar]
- Rull, V.; Trapote, M.C.; Safont, E.; Cañellas-Boltà, N.; Pérez-Zanón, N.; Sigró, J.; Buchaca, T.; Vegas-Vilarrúbia, T. Seasonal patterns of pollen sedimentation in Lake Montcortès (Central Pyrenees) and potential applications to high-resolution paleoecology: A 2-year pilot study. J. Paleolimnol. 2017, 57, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, E.; Aniz, M. Guía del Parque Nacional de Aigüestortes i Estany de Sant Maurici; Organismo Autónomo de Parques Nacionales: Madrid, Spain, 2013. [Google Scholar]
- Mercadé, A.; Vigo, J.; Rull, V.; Vegas-Vilarrúbia, T.; Garcés, S.; Cañellas-Boltà, N. Vegetation and landscape around Lake Montcortès (Catalan pre-Pyrenees) as a tool for palaeoeological studies of lake sediments. Colect. Bot. 2013, 32, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Carreras, J.; Vigo, J.; Ferré, A. Manual dels Hàbitats de Catalunya; Departament de Medi Ambient i Habitatge, Generalitat de Catalunya: Barcelona, Spain, 2005.
- Corella, J.P.; Brauer, A.; Mangili, C.; Rull, V.; Vegas-Vilarrúbia, T.; Morellón, M.; Valero-Garcés, B. The 1.5-ka varved record of Lake Montcortès (southern Pyrenees, NE Spain). Quat. Res. 2012, 78, 323–332. [Google Scholar] [CrossRef]
- Davis, M.B. Climatic instability, time lags, and community disequilibrium. In Community Ecology; Diamond, J., Case, T.J., Eds.; Harper & Row: New York, NY, USA, 1984; pp. 269–284. [Google Scholar]
- Ter Braak, C.J.F.; Prentice, I.C. A theory of gradient analysis. Adv. Ecol. Res. 1988, 18, 271–317. [Google Scholar]
- Jongman, R.H.G.; Ter Braak, C.J.F.; Van Tongeren, O.F.R. Data Analysis in Community and Landscape Ecology; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Kendall, M.G. Rank Correlation Methods; Griffin: London, UK, 1970. [Google Scholar]
- Pearson, K. Notes on regression and inheritance in the case of two parents. Proc. R. Soc. Lond. 1895, 58, 240–242. [Google Scholar]
- Bennett, K.D.; Humphry, R.V. Analysis of Late-glacial and Holocene rates of vegetational change at two sites in the British Isles. Rev. Palaeobot. Palynol. 1995, 85, 263–287. [Google Scholar] [CrossRef]
- Rull, V. Successional patterns of the Gran Sabana (sotheastern Venezuela) vegetation during the last 5000 years, and its responses to climatic fluctuations and fire. J. Biogeogr. 1992, 19, 329–338. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley: New York, NY, USA, 1984. [Google Scholar]
- Legendre, L.; Legendre, L. Numerical Ecology; Elsevier: New York, NY, USA, 1983. [Google Scholar]
- Gareth, J.; Witten, D.; Hastie, T.; Tibshirani, R. An Introduction to Statistical Learning; Springer: New York, NY, USA, 2017. [Google Scholar]
- Kovach, W.L. Comparisons of multivariate analytical techniques for use in pre-Quaternary plant paleoecology. Rev. Palaeobot. Palynol. 1989, 60, 255–282. [Google Scholar] [CrossRef]
- Kovach, W.L. Multivariate techniques for biostratigraphical correlation. J. Geol. Soc. 1993, 150, 697–705. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T. Paleontological Data Analysis; Blackwell: London, UK, 2006. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontologic statistics software for education and data analysis. Palaeontol. Electr. 2001, 4, 9. [Google Scholar]
- Bennett, K.D. Determination of the number of zones in a biostratigraphical sequence. New Phytol. 1996, 132, 155–170. [Google Scholar] [CrossRef]
- Kaul, A.; Mandal, S.; Davidov, O.; Peddada, S.D. Analysis of microbiome data in the presence of excess zeros. Front. Micorbiol. 2017, 8, 2114. [Google Scholar] [CrossRef]
- Silverman, J.D.; Roche, K.; Mukherjee, S.; David, L.A. Naughty all zeros in sequence count data are the same. Comp. Struct. Biotech. J. 2020, 18, 2789–2798. [Google Scholar] [CrossRef]
- Marugan, C.M.; Rapalino, V. Història del Pallars. Dels Orígens als Nostres Dies; Pagès Editors: Lleida, Spain, 2005. [Google Scholar]
- Rull, V. Cultural development of the Pallars region (NE Iberian Peninsula) from the Bronze Age to the present. PaleorXiv 2021. [Google Scholar] [CrossRef]
- Bringué, J.M. L’edat moderna. In Història del Pallars. Dels Orígens als Nostres Dies; Marugan, C.M., Rapalino, V., Eds.; Pagès Editors: Lleida, Spain, 2005; pp. 87–119. [Google Scholar]
- Marugan, C.M.; Oliver, J. El Pallars medieval. In Història del Pallars. Dels Orígens als Nostres Dies; Marugan, C.M., Rapalino, V., Eds.; Pagès Editors: Lleida, Spain, 2005; pp. 45–86. [Google Scholar]
- Farràs, F. El Pallars contemporani. In Història del Pallars. Dels Orígens als Nostres Dies; Marugan, C.M., Rapalino, V., Eds.; Pagès Editors: Lleida, Spain, 2005; pp. 121–144. [Google Scholar]
- Majeed, H.T.; Periago, C.; Alarcón, M.; Belmonte, J. Airborne parameters and their relationship with meteorological variables in NE Iberian Peninsula. Aerobiologia 2018, 34, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Mathys, A.S.; Brang, P.; Stillhard, J.; Bugmann, H.; Hobi, M.L. Long-term tree species population dynamics in Swiss forest reserves influenced by forest structure and climate. For. Ecol. Manag. 2021, 481, 118666. [Google Scholar] [CrossRef]
- Wang, W.J.; Thompson, F.R.; He, H.S.; Fraser, J.S.; Dijack, W.D.; Spetich, M.A. Population dynamics has greater effects than climate change on tree species distribution in a temperate forest region. J. Biogeogr. 2018, 45, 2766–2778. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sites | A | E (m) | AR (yr) | NS | YS | Resolution | References |
|---|---|---|---|---|---|---|---|
| Marboré (L) | Mb | 2612 | 15,000 | 80 | 188 | Bicentennial | [11] |
| Forcat (L) | Ft | 2531 | 9500 | 105 | 90 | Centennial | [12] |
| Estany Blau (L) | EB | 2471 | 1200 | 40 | 30 | Multidecadal | [12] |
| Serra Mitjana (B) | SM | 2406 | 1500 | 15 | 100 | Centennial | [13] |
| Riu dels Orris (B) | RO | 2390 | 8000 | 60 | 133 | Centennial | [14] |
| Orris del Seut (B) | OS | 2300 | 3700 | 32 | 116 | Centennial | [14] |
| Estanilles (B) | Et | 2247 | 12,000 | 90 | 133 | Centennial | [15] |
| Redon (L) | Rn | 2240 | 10,000 | 130 | 77 | Subcentennial | [16,17] |
| Planells de Perafita (B) | PP | 2240 | 10,000 | 50 | 200 | Bicentennial | [12] |
| Bosc dels Estanyons (B) | BE | 2180 | 12,000 | 90 | 133 | Centennial | [14,18] |
| Ibón de las Ranas (L) | IR | 2092 | 10,000 | 40 | 250 | Bicentennial | [19] |
| Pradell (B) | Pd | 1975 | 1400 | 37 | 38 | Multidecadal | [20] |
| Sant Maurici (L) | Ma | 1914 | 2000 | 71 | 28 | Multidecadal | [21] |
| Basa de la Mora (L) | BM | 1914 | 15,000 | 140 | 107 | Centennial | [22] |
| Bassa Nera (B) | BN | 1891 | 7000 | 110 | 64 | Subcentennial | [23,24] |
| Coma de Burg (L) | CB | 1821 | 10,000 | 70 | 143 | Centennial | [25] |
| Tramacastilla (L) | Tm | 1668 | 15,000 | 150 | 100 | Centennial | [19] |
| Llebreta (L) | Ll | 1619 | 3600 | 240 | 15 | Bidecadal | [26] |
| Prats de Vila (B) | PV | 1150 | 2200 | 30 | 73 | Subcentennial | [27] |
| Estanya (L) | Es | 670 | 20,000 | 65 | 308 | Multicentennial | [28,29] |
| Quercus (d) | Betula | Quercus (e) | Pinus | |
|---|---|---|---|---|
| Temp1 | 0.374 | 0.251 | 0.104 | −0.014 |
| Temp2 | 0.322 | 0.133 | 0.118 | −0.180 |
| PPT | −0.089 | −0.288 | 0.231 | −0.039 |
| DGT | −0.036 | −0.064 | −0.225 | −0.032 |
| Olea | 0.220 | 0.292 | −0.195 | −0.408 |
| Cereals | −0.325 | −0.207 | −0.367 | −0.343 |
| Sporormiella | −0.330 | 0.170 | −0.295 | −0.334 |
| Charcoal | −0.402 | 0.064 | −0.293 | −0.114 |
| PC1 | PC2 | PC3 | |
|---|---|---|---|
| Quercus (d) | −0.082 | 0.452 | 0.841 |
| Betula | −0.068 | 0.019 | 0.285 |
| Cornus | −0.004 | 0.014 | −0.013 |
| Salix | −0.012 | −0.030 | 0.006 |
| Alnus | 0.008 | −0.001 | 0.021 |
| Tilia | −0.002 | 0.004 | −0.005 |
| Quercus (e) | 0.120 | 0.886 | −0.414 |
| Corylus | 0.006 | 0.068 | −0.123 |
| Fagus | 0.013 | 0.006 | −0.057 |
| Abies | 0.000 | 0.003 | −0.005 |
| Pinus | 0.987 | −0.070 | 0.142 |
| Morus | 0.009 | 0.004 | −0.007 |
| Ulmus | 0.020 | 0.019 | 0.001 |
| Populus | 0.002 | 0.006 | 0.000 |
| Fraxinus | 0.004 | −0.001 | −0.008 |
| Cumulative variance (%) | 52.329 | 76.639 | 91.075 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rull, V.; Vegas-Vilarrúbia, T. Climatic and Anthropogenic Drivers of Forest Succession in the Iberian Pyrenees during the Last 500 Years: A Statistical Approach. Forests 2022, 13, 622. https://doi.org/10.3390/f13040622
Rull V, Vegas-Vilarrúbia T. Climatic and Anthropogenic Drivers of Forest Succession in the Iberian Pyrenees during the Last 500 Years: A Statistical Approach. Forests. 2022; 13(4):622. https://doi.org/10.3390/f13040622
Chicago/Turabian StyleRull, Valentí, and Teresa Vegas-Vilarrúbia. 2022. "Climatic and Anthropogenic Drivers of Forest Succession in the Iberian Pyrenees during the Last 500 Years: A Statistical Approach" Forests 13, no. 4: 622. https://doi.org/10.3390/f13040622
APA StyleRull, V., & Vegas-Vilarrúbia, T. (2022). Climatic and Anthropogenic Drivers of Forest Succession in the Iberian Pyrenees during the Last 500 Years: A Statistical Approach. Forests, 13(4), 622. https://doi.org/10.3390/f13040622