
Identification and Analysis of JAZ Gene Family in Ginkgo biloba Reveals Candidate Genes for Biosynthesis of Terpene Trilactones

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of GbJAZs
2.2. Bioinformatics Analysis of GbJAZs
2.3. Expression Analysis of GbJAZs
2.4. Determination of TTL Content in G. biloba Leaves
3. Results
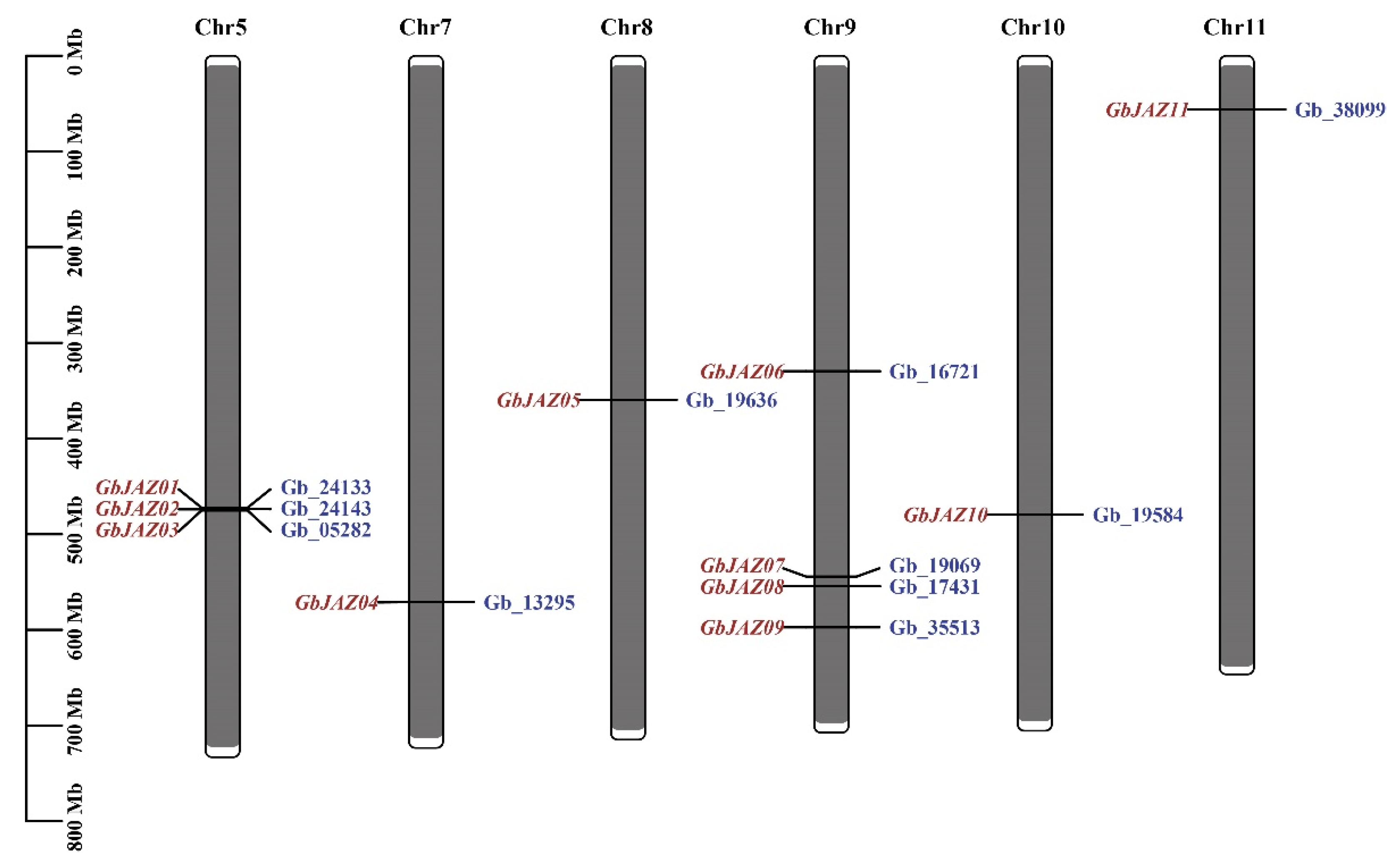
3.1. Identification and Physicochemical Properties Analysis of GbJAZs
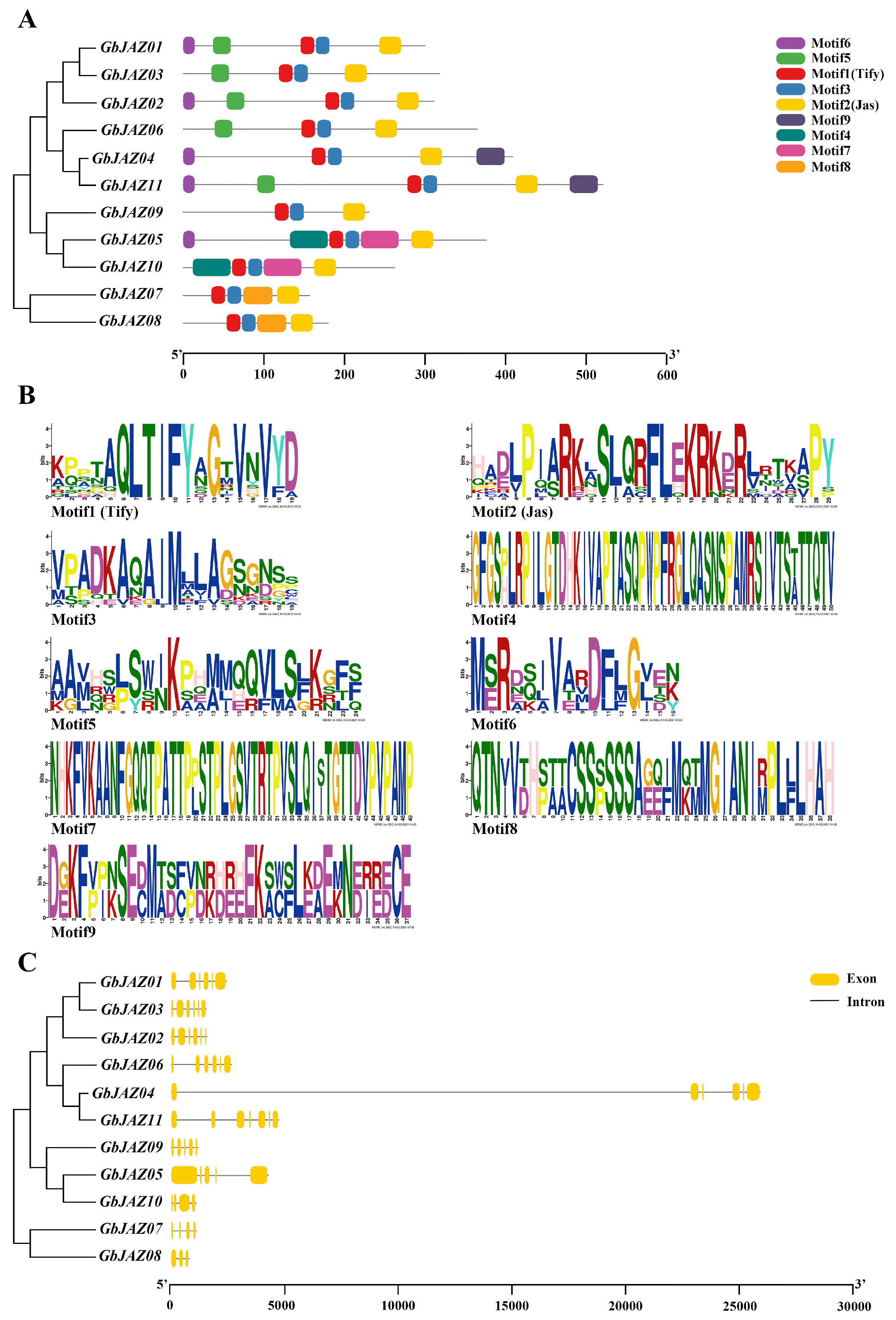
3.2. Gene Structure Analysis of GbJAZs
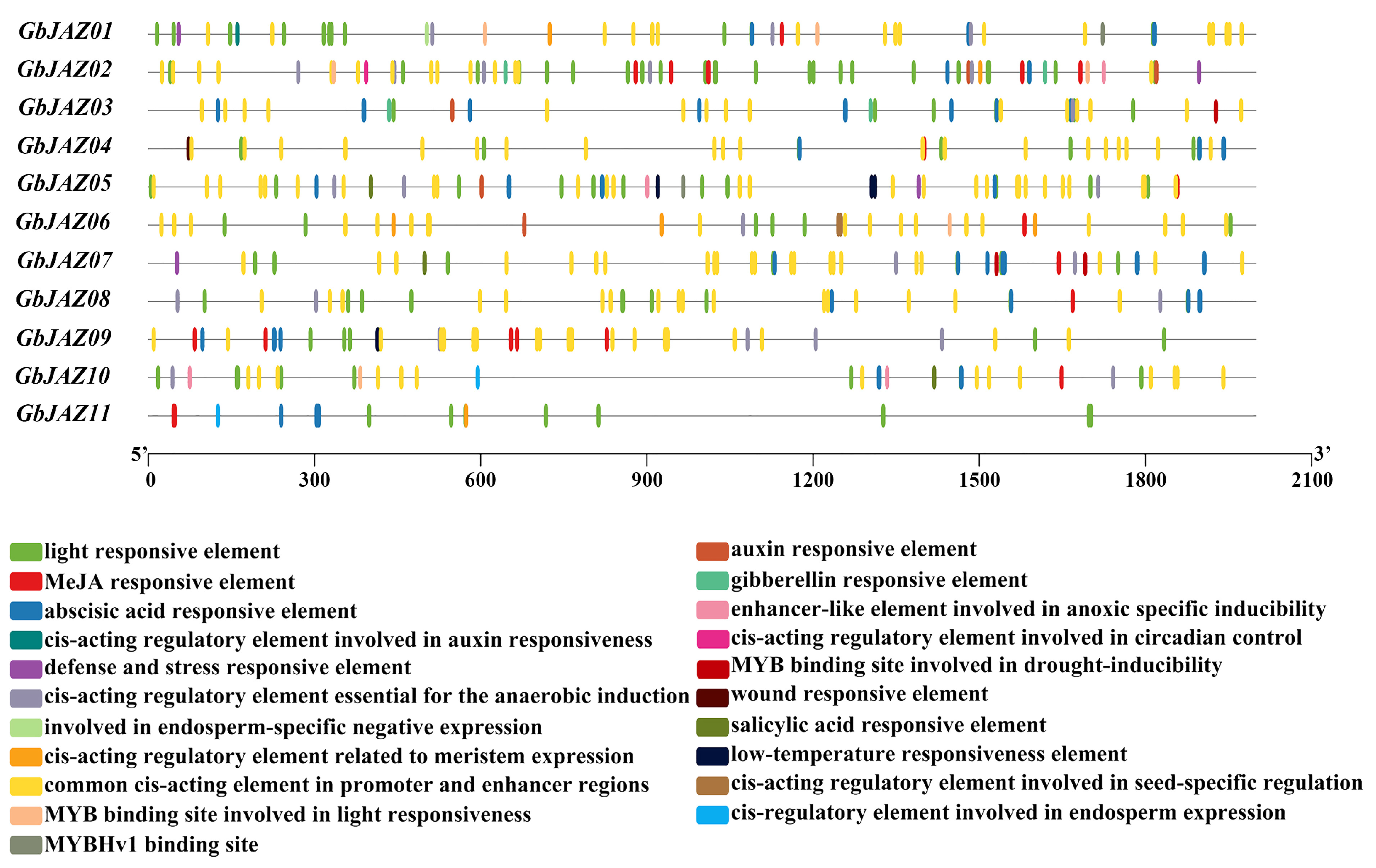
3.3. Cis-Acting Elements Analysis of GbJAZs
3.4. Phylogenetic Analysis of GbJAZs
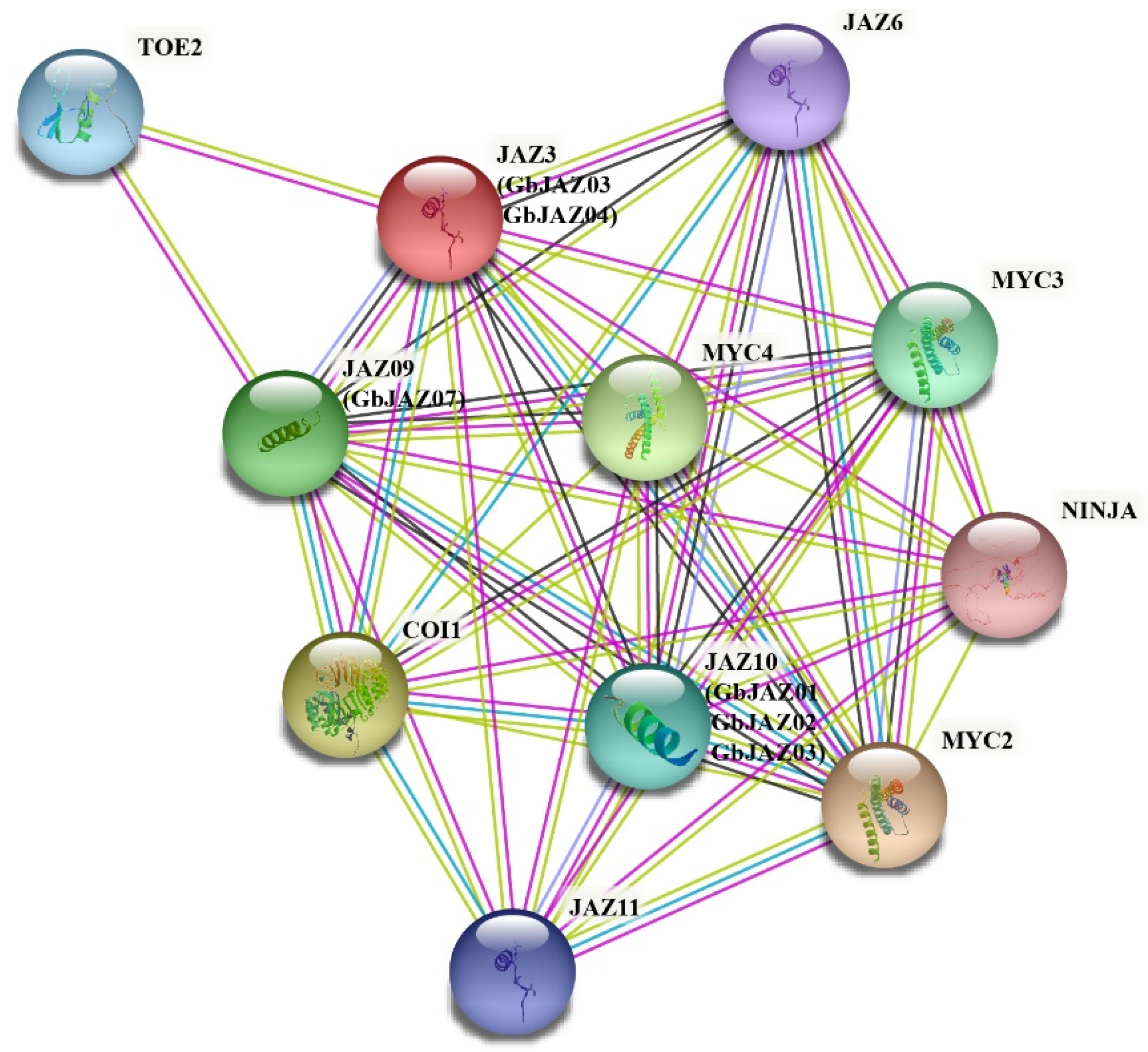
3.5. Protein–Protein Interaction Network Prediction of GbJAZs
3.6. Expression Patterns of GbJAZs
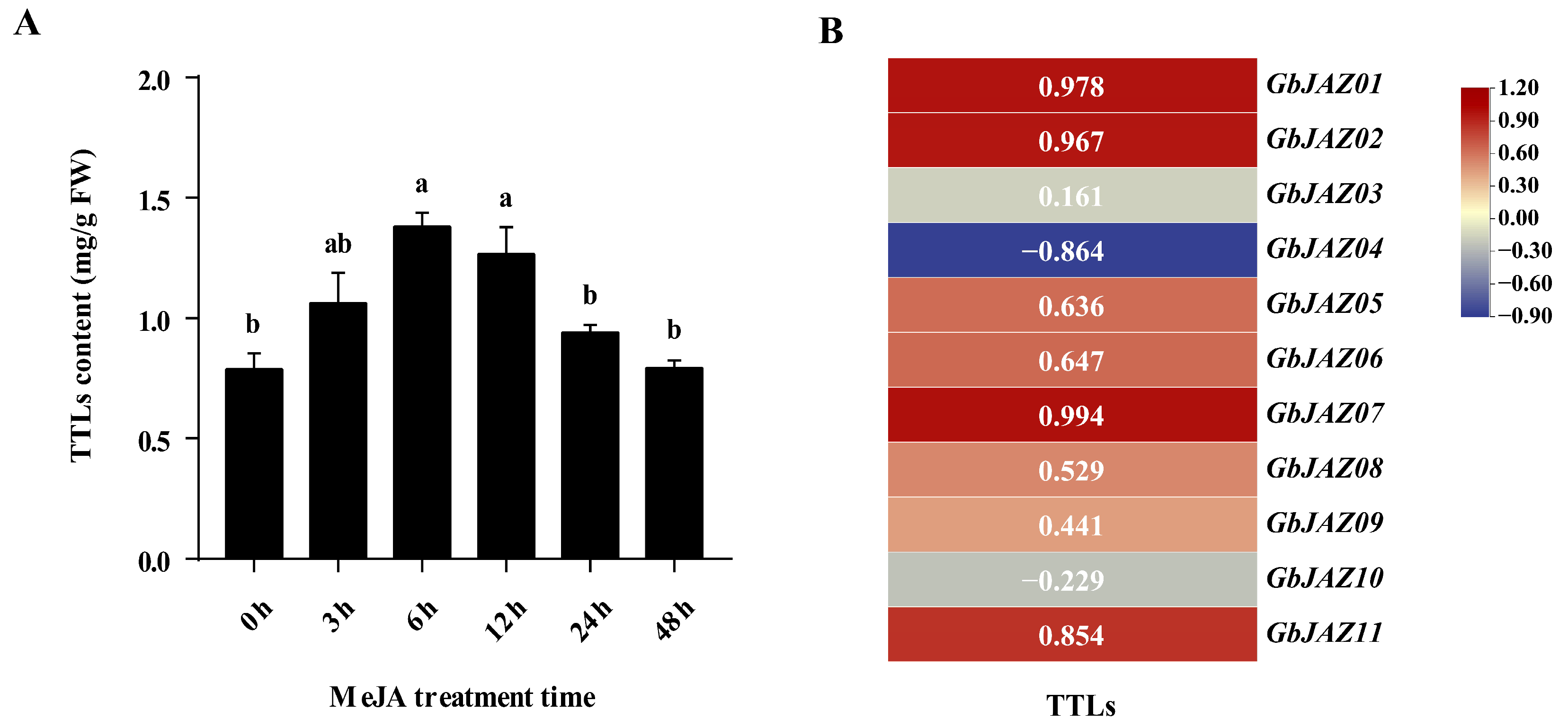
3.7. Correlation Analysis between the Expression Levels of GbJAZs and TTL Content in G. biloba
4. Discussion
4.1. Specific Motifs of GbJAZs
4.2. Different Expression Patterns of GbJAZs
4.3. Identification of Candidate GbJAZs Involved in Regulating the Biosynthesis of TTLs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cui, T. Studies on Regulation of Ginkgo Flavonesand and Ginkgolides Accumulation in Cultured Cells of Ginkgo biloba L. and Their Pharmacology; South China University of Technology: Guangzhou, China, 2002. (In Chinese) [Google Scholar]
- Hu, Z. A research on Extraction and Preparation Technology of Flavonoids from Ginkgo biloba Leaves; Jinan University: Guangzhou, China, 2017. (In Chinese) [Google Scholar]
- Mandaokar, A.; Thines, B.; Shin, B.; Lange, B.; Choi, G.; Koo, Y.; Yoo, Y.; Yang, D.; Choi, G.; Browse, J.; et al. Transcriptional regulators of stamen development in Arabidopsis identified by transcriptional profiling. Plant J. Cell Mol. Biol. 2010, 46, 984–1008. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, L.; Morreel, K.; Witte, E.D.; Lammertyn, F.; Montagu, M.; Boerjan, W.; Inze, D.; Goossens, A. Mapping methyl jasmonate-mediated transcriptional reprogramming of metabolism and cell cycle progression in cultured Arabidopsis cells. Proc. Natl. Acad. Sci. USA 2008, 105, 1380–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauwels, L.; Goossens, A. The JAZ proteins: A crucial interface in the jasmonate signaling cascade. Plant Cell 2011, 23, 3089–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wager, A.; Browse, J. Social network: JAZ protein interactions expand our knowledge of Jasmonate signaling. Front. Plant Sci. 2012, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Staswick, P.E. JAZing up jasmonate signaling. Trends Plant Sci. 2008, 13, 66–71. [Google Scholar] [CrossRef]
- Vanholme, B.; Grunewald, W.; Bateman, A.; Kohchi, T.; Gheysenet, G. The Tify family previously known as ZIM. Trends Plant Sci. 2007, 12, 239–244. [Google Scholar] [CrossRef]
- Bai, Y.; Meng, Y.; Huang, D.; Qi, Y.; Chen, M. Origin and evolutionary analysis of the plant-specific TIFY transcription factor family. Genomics 2011, 98, 128–136. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.S.; Howe, G.A. A critical role for the TIFY motif in repression of jasmonate signalng by a stabilized splice variant of the JASMONATE ZIM-domain protein JAZ10 in Arabidopsis. Plant Cell 2009, 21, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Han, B.; Liu, J.; Zhao, X.; Tang, Z. Bioinformatics analysis of JAZ gene family in maize. J. Shanxi Agric. Sci. 2020, 48, 1552–1556. (In Chinese) [Google Scholar]
- Sheard, L.B.; Tan, X.; Mao, H.B.; Withers, J.; Nissan, G.B.; Hinds, T.R.; Kobayashi, Y.; Hsu, F.F.; Sharon, M.; Browse, J.; et al. Jasmonate perception by inositol-phosphate-potentiated COI1–JAZ co-receptor. Nature 2010, 468, 400–405. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. JAZ repressors and the orchestration of phytohormone crosstalk. Trends Plant Sci. 2012, 17, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Sonbol, F.M.; Huot, B.; Gu, Y.; Withers, J.; Mwimba, M.; Jian, Y.; Sheng, Y.; Dong, X. Salicylic acid receptors activate jasmonic acid signalling through a non-canonical pathway to promote effector-triggered immunity. Nat. Commun. 2016, 7, 13099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, L.; Jing, Y.; Shi, P.; Liu, J. JAZ proteins modulate seed germination through interaction with ABI5 in bread wheat and Arabidopsis. New Phytol. 2019, 223, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Liu, Z.; Qu, S.; Yu, L.; Zhan, Y. Effects of transient infection of FmJAZ1 gene on JA pathway related gene expression in Fraxinus mandshurica. Guihaia 2021, 41, 662–670. (In Chinese) [Google Scholar]
- Taniguchi, S.; Hosokawa, S.Y.; Tamaoki, D.; Yamada, S.; Akimitsu, K.; Gomi, K. Jasmonate induction of the monoterpene linalool confers resistance to rice bacterial blight and its biosynthesis is regulated by JAZ proteinin rice. Plant Cell Environ. 2014, 37, 451–461. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, Y.; Nie, L.; Jin, X.; Fu, C.; Yu, L. Molecular, structural, and phylogenetic analyses of Taxus chinensis JAZs. Gene 2017, 620, 66–74. [Google Scholar] [CrossRef]
- Zhang, M. The JA Regulation Mechanism on Taxol Biosynthetic in Taxus chinensis; Huazhong University of Science and Technology: Wuhan, China, 2016. (In Chinese) [Google Scholar]
- Dong, Y.; Zhang, W.; Ling, Z. Advances in transcription factors regulating plant terpenoids biosynthesis. Bull. Bot. 2020, 55, 340–350. (In Chinese) [Google Scholar]
- Guan, R.; Zhao, Y.; Zhang, H.; Fan, G.; Liu, X.; Zhou, W.; Shi, C.; Wang, J.; Liu, W.; Liang, X.; et al. Draft genome of the living fossil Ginkgo biloba. Gigascience 2016, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Deng, S.; Shen, J. Identification and response to phytophthora sojae infections of JAZ gene familyin Soybean. Soybean Sci. 2019, 38, 868–878. (In Chinese) [Google Scholar]
- Chen, S.; Liu, X.; Liao, Y.; Zhang, W.; Xu, F. Genome-wide identification of WRKY family genes and analysis of their expression in response to abiotic stress in Ginkgo biloba L. Not. Bot. Horti Agrobot. Cluj Napoca 2019, 47, 1100–1115. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Xin, H.; Gu, X. Genome-wide identification and functional analysis of the Basic Helix-Loop-Helix (bHLH) transcription family reveals candidate PtFBH genes involved in the flowering process of Populus trichocarpa. Forests 2021, 12, 1439. [Google Scholar] [CrossRef]
- Zhou, X.; Liao, Y.; Kim, S.; Chen, Z.; Xu, F. Genome-wide identifcation and characterization of bHLH family genes from Ginkgo bilob. Sci. Rep. 2020, 10, 13723. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Xu, F.; Song, Q.; Ye, J.; Liao, Y.; Xu, F. Isolation, characterization and functional analysis of a novel 3-hydroxy-3-methylglutaryl-coenzyme A synthase gene (GbHMGS2) from Ginkgo biloba. Acta Physiol. Plant. 2018, 40, 72. [Google Scholar] [CrossRef]
- Ye, J.; Mao, D.; Cheng, S.; Zhang, X.; Xu, F. Comparative transcriptome analysis reveals the potential stimulatory mechanism of terpene trilactone biosynthesis by exogenous salicylic acid in Ginkgo biloba. Ind. Crops Prod. 2020, 145, 112104. [Google Scholar] [CrossRef]
- Xu, F.; Ning, Y.; Zhang, W.; Liao, Y.; Li, L.; Cheng, H.; Cheng, S. An R2R3-MYB transcription factor as a negative regulator of the flavonoid biosynthesis pathway in Ginkgo biloba. Funct. Integr. Genom. 2014, 14, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Zheng, J.; Zhang, X.; Fu, M.; Zeng, H.; Ye, J.; Zhang, W.; Liao, Y.; Xu, F. Effects of different stress treatments on the total terpene trilactone content and expression levels of key genes in Ginkgo biloba leaves. Plant Mol. Biol. Rep. 2020, 38, 521–530. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Chico, J.M.; Calvo, P.F.; Solano, R. The ZIM domain mediates homo- and heteromeric interactions between Arabidopsis JAZ proteins. Plant J. Cell Mol. Biol. 2009, 59, 77–87. [Google Scholar] [CrossRef]
- Melotto, M.; Mecey, C.; Niu, Y.; Chung, H.S.; Katsir, L.; Yao, J.; Zeng, W.; Thines, B.; Staswick, P.; Browse, J.; et al. A critical role of two positively charged amino acids in the Jas motif of Arabidopsis JAZ proteins in mediating coronatine and jasmonoyl isoleucine-dependent interactions with the COI1 F-box protein. Plant J. 2008, 55, 979–988. [Google Scholar] [CrossRef] [Green Version]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J.; et al. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef]
- Hou, X.; Li, Y.; Cheng, C.; Xia, K. DELLAs modulate jasmonate signaling via competitive binding to JAZs. Dev. Cell 2010, 19, 884–894. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ma, B.; Qi, X.; Guo, Q.; Wang, X.; Zeng, Q.; He, N. Identification and characterization of genes involved in the jasmonate biosynthetic and signaling pathways in mulberry (Morus notabilis). J. Integr. Plant Biol. 2014, 56, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K. Negative regulation of defence and stress genes by EAR-motif-containing repressors. Trends Plant Sci. 2006, 11, 109–112. [Google Scholar] [CrossRef]
- Kagale, S.; Rozwadowski, L.K. Genome-wide analysis of ethylene-responsive element binding factor-associated amphiphilic repression motif-containing transcriptional regulators in Arabidopsis. Plant Physiol. 2010, 152, 1109–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiratsu, K.; Matsui, K.; Koyama, T.; Takagi, M.O. Dominant repression of target genes by chimeric repressors that include the EAR motif, a repression domain, in Arabidopsis. Plant J. Cell Mol. Biol. 2003, 34, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, Z.; Wu, Z. Progress of the structural and functional analysis of plant transcription factor TIFY protein family. Biotechnol. Bull. 2020, 36, 121–128. (In Chinese) [Google Scholar]
- Li, X.; Yin, X.; Wang, H.; Guo, C.; Gao, H.; Zheng, Y.; Fan, C.; Wang, X. Genome-wide identification and analysis of the apple (Malus × domestica Borkh.) TIFY gene family. Tree Genet. Genomes 2015, 11, 1–13. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, M.; Singer, S.D.; Fei, Z.; Hua, W.; Wang, X.; Yang, H. Genome-wide identification and analysis of the TIFY gene family in grape. PLoS ONE 2012, 7, e44465. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhang, P.; Cheng, C.; Xia, G. The moss jasmonate ZIM-domain protein PnJAZ1 confers salinity tolerance via crosstalk with the abscisic acid signalling pathway. Plant Sci. 2019, 280, 1–11. [Google Scholar] [CrossRef]
- Wen, J.; Li, Y.; Qi, T.; Hua, G.; Song, S. The C-terminal domains of Arabidopsis GL3/EGL3/TT8 interact with JAZ proteins and mediate dimeric interactions. Plant Signal. Behav. 2018, 13, e1422460. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Zhen, J.; Jiang, Z. Cloning and expression analysis of cold response gene GhJAZ1 from Gossypium hirsutum L. Acta Agric. Boreali Sin. 2018, 33, 7–13. (In Chinese) [Google Scholar]
- Pei, T. The Function of SmJAZ in Regulating the Biosynthesis of Salvianolic Acids and Tanshinones in Salvia miltiorrhiza; Northwest A&F University: Xianyang, China, 2019. (In Chinese) [Google Scholar]
- Xue, L.; Liu, X.; Luo, Y. Expansion and expression analysis of PtMAP65 gene family in Populus trichocarpa. For. Res. 2021, 34, 92–101. (In Chinese) [Google Scholar]
- Hong, G.; Xue, X.; Mao, Y.; Wang, L.; Chen, X. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagee, D.; Hardan, A.; Botero, J.; Arnone, J.T. Genomic clustering within functionally related gene families in Ascomycota fungi. Comput. Struct. Biotechnol. J. 2020, 18, 3267–3277. [Google Scholar] [CrossRef]
- Arnone, T. Genomic considerations for the modification of Saccharomyces cerevisiae for biofuel and metabolite biosynthesis. Microorganisms 2020, 8, 321. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Zhou, W.; Zhang, J.; Huang, S.; Wang, H.; Kai, G. Methyl jasmonate induction of tanshinone biosynthesis in Salvia miltiorrhiza hairy roots is mediated by JASMONATE ZIM-DOMAIN repressor proteins. Sci. Rep. 2016, 6, 20919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, T.; Chen, M.; Shen, Q.; Li, L.; Fu, X.; Pan, Q.; Tang, Y.; Shi, P.; Lv, Z.; Jiang, W.; et al. HOMEODOMAIN PROTEIN 1 is required for jasmonate-mediated glandular trichome initiation in Artemisia annua. New Phytol. 2016, 213, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Yan, H. Role of JAZ2 and JAZ7 in Regulating Jasmonic Acid-Mediated Transcriptional and Metabolic Reprogramming; Zhejiang University: Hangzhou, China, 2014. (In Chinese) [Google Scholar]
- Chuquimarca, S.; Ayala-Ruano, S.; Goossens, J.; Pauwels, L.; Goossens, A.; Leon-Reyes, A.; Méndez, M.Á. The molecular basis of JAZ-MYC coupling, a protein-protein interface essential for plant response to stressors. Front. Plant Sci. 2020, 11, 1139. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence F | Primer Sequence R |
|---|---|---|
| GbJAZ01 | CGATCGTAGCTGTGGACTTT | GCTCTCTCGTTCTTCTGTTGAT |
| GbJAZ02 | GCTTGCTGGTAGCGGAAATA | TAATGGTGTTGTCGGGTTGAG |
| GbJAZ03 | GACTTTCTGGGTGTCGAGAAG | GCTAGGCCAAATGCCAAATC |
| GbJAZ04 | GCTTGGAATTGGCAGAATGG | GTCTTTGTCTGCGTCCTCTT |
| GbJAZ05 | TGGACTGCAGGCTTCAAAT | GACACTAACCATCCCAGCATAG |
| GbJAZ06 | TGGCGGTCTCTTCCTGTATAA | GTGCCTTCCAACGACTTCTATC |
| GbJAZ07 | GCCTGACCCACAAGAAGAAA | GTAGACCCTCCCTTGGTAGAA |
| GbJAZ08 | CATGCCCACATTCCCAAATTAC | TTTGCGTAGGCGTCTTGTAG |
| GbJAZ09 | GTGCCATTAAAGCTCCGAAATAG | GACCTTGCGTTGAGTCTCTTAT |
| GbJAZ10 | GGTTACAAGAACACCCGTATCA | GAACTGTTTGGTTCTGGTTTGG |
| GbJAZ11 | CCTGAGCATTCTTCCCATAGAG | CCATCCTGCTGTTGTGATTTG |
| Gene | Amino Acid | Molecular Mass/KDa | pI | Instability Index | Hydrophilicity | Chromosome Location | Subcellular Location |
|---|---|---|---|---|---|---|---|
| GbJAZ01 | 301 | 32.19 | 9.46 | 38.08 | −0.357 | chr5 | nucleus |
| GbJAZ02 | 312 | 33.82 | 9.36 | 42.47 | −0.60 | chr5 | nucleus |
| GbJAZ03 | 319 | 34.90 | 9.78 | 48.69 | −0.49 | chr5 | nucleus |
| GbJAZ04 | 410 | 43.92 | 7.59 | 60.03 | −0.64 | chr7 | nucleus |
| GbJAZ05 | 377 | 40.27 | 9.03 | 52.86 | −0.58 | chr8 | nucleus |
| GbJAZ06 | 366 | 41.02 | 9.77 | 56.90 | −0.75 | chr9 | nucleus |
| GbJAZ07 | 157 | 17.46 | 9.60 | 60.01 | −0.48 | chr9 | nucleus |
| GbJAZ08 | 180 | 19.67 | 7.94 | 54.52 | −0.34 | chr9 | nucleus |
| GbJAZ09 | 231 | 25.53 | 9.22 | 89.63 | −0.65 | chr9 | nucleus |
| GbJAZ10 | 263 | 28.24 | 9.54 | 50.62 | −0.19 | chr10 | membrane/nucleus |
| GbJAZ11 | 522 | 56.38 | 8.95 | 46.56 | −0.63 | chr11 | nucleus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Liu, X.; Zheng, J.; Ye, J.; Liao, Y.; Xu, F. Identification and Analysis of JAZ Gene Family in Ginkgo biloba Reveals Candidate Genes for Biosynthesis of Terpene Trilactones. Forests 2022, 13, 781. https://doi.org/10.3390/f13050781
He X, Liu X, Zheng J, Ye J, Liao Y, Xu F. Identification and Analysis of JAZ Gene Family in Ginkgo biloba Reveals Candidate Genes for Biosynthesis of Terpene Trilactones. Forests. 2022; 13(5):781. https://doi.org/10.3390/f13050781
Chicago/Turabian StyleHe, Xiao, Xiaomeng Liu, Jiarui Zheng, Jiabao Ye, Yongling Liao, and Feng Xu. 2022. "Identification and Analysis of JAZ Gene Family in Ginkgo biloba Reveals Candidate Genes for Biosynthesis of Terpene Trilactones" Forests 13, no. 5: 781. https://doi.org/10.3390/f13050781
APA StyleHe, X., Liu, X., Zheng, J., Ye, J., Liao, Y., & Xu, F. (2022). Identification and Analysis of JAZ Gene Family in Ginkgo biloba Reveals Candidate Genes for Biosynthesis of Terpene Trilactones. Forests, 13(5), 781. https://doi.org/10.3390/f13050781