
Functional Diversity and Its Influencing Factors in a Subtropical Forest Community in China


Abstract
:1. Introduction
2. Materials and Methods
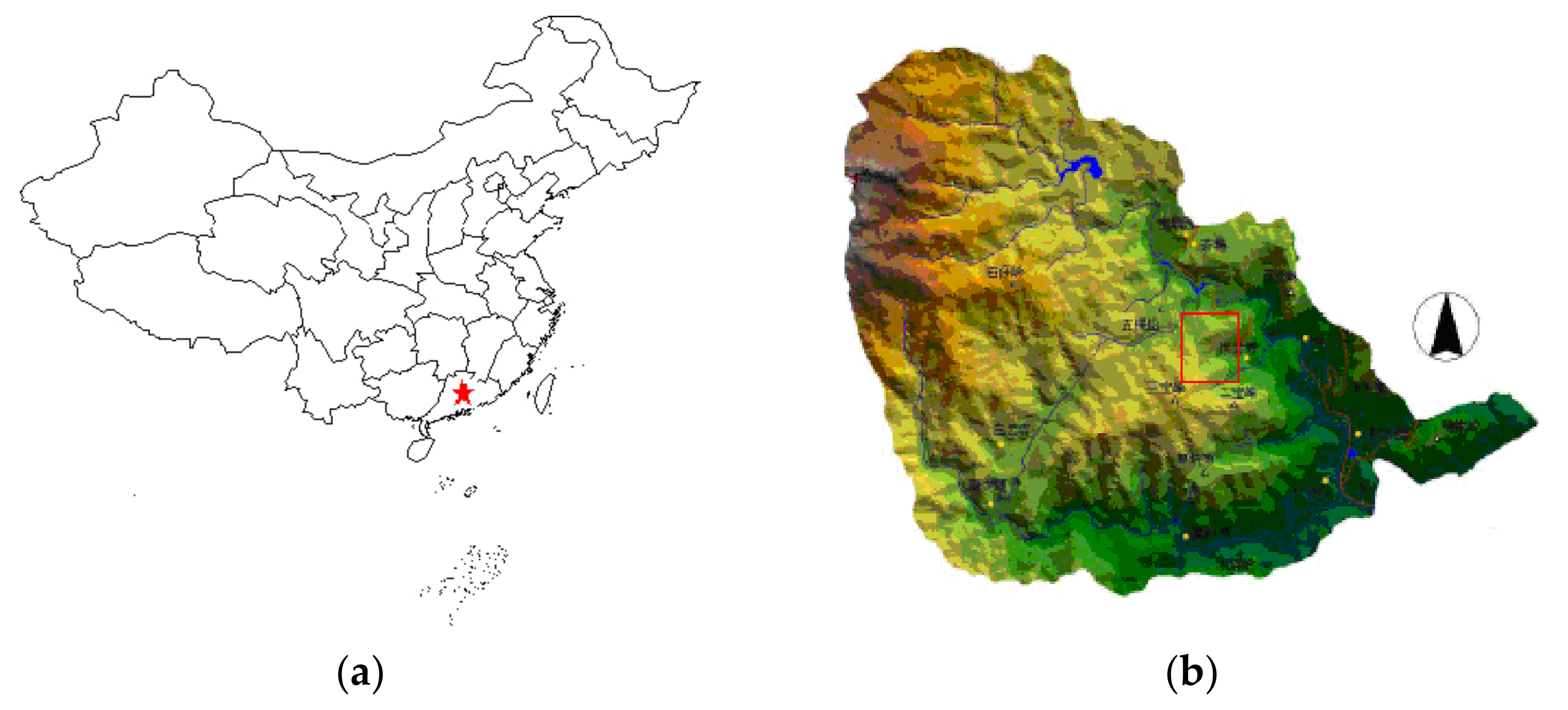
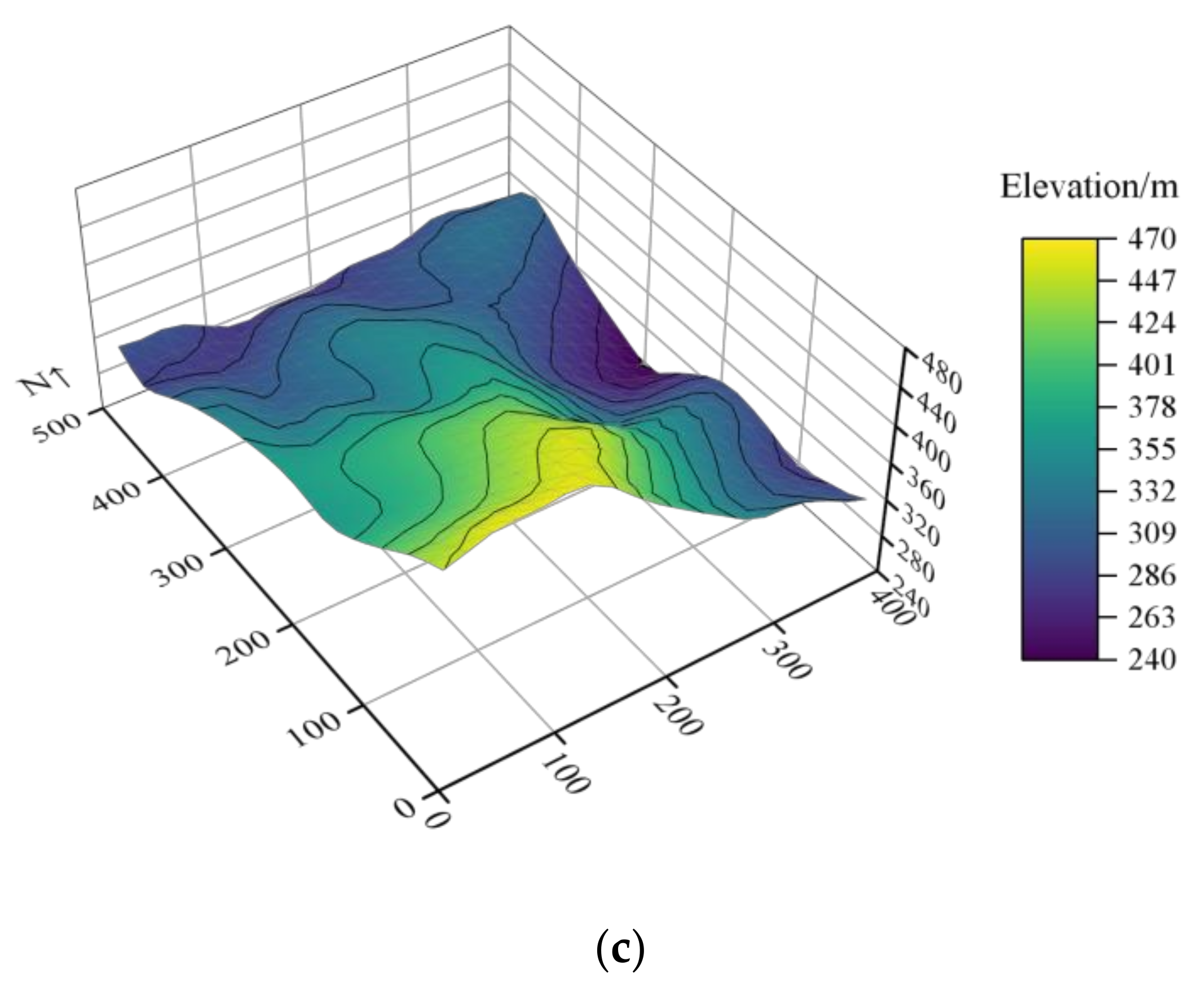
2.1. Study Area and Data
2.2. Functional Traits
2.3. Environmental Factors
2.4. Functional and Biodiversity Indices
2.4.1. Functional Diversity Indices
- (1)
- Average distance of functional traits (MFAD): MFAD is average of the Euclidean distances between species pairs in the trait space [31]. In order to avoid the known correlation between distance of functional traits and species richness, we used the average distance of functional traits (MFAD) to characterize the multidimensional functional richness [31]:where dij is the functional feature distance between species, and N is the number of species.
- (2)
- (3)
- (4)
- Functional evenness index (FEve): FEve describes the evenness of abundance distribution in a functional trait space [31]. FEve was calculated by taking the distance between all species pairs and generating a minimum spanning tree weighted by relative abundance, which connected all species in multi-dimensional character space. This index measures both the uniformity of the minimum spanning branch length and the uniformity of species abundance. The calculation formula is below [31]:where EW1 is the weighted evenness, PEW1 is the partial weighted evenness. dist(i,j) is the Euclidean distance between species i and species j, and Wi is the relative abundance of species i, S is the number of species.
- (5)
- Multidimensional functional divergence (FDiv): FDiv relates to how abundance is distributed within the volume of functional trait space occupied by species, FDiv was calculated according to [31]:where the Euclidean distance is obtained by calculating the square and square root of the distance between each species character value and the center of gravity of the convex polygon, is the mean value of the Euclidean distance of each species, is the absolute value of the distance when calculating , and is the relative abundance of species i.
- (6)
- Functional divergence index (FDis): FDis indicates the average weighted distance between each species and the center of gravity in multidimensional character space, where the center of gravity is the center of gravity of all species. It was calculated by this formula [32]:where is the abundance of species j and is the distance between species j and the weighted center of gravity.
- (7)
- Rao’s quadratic entropy index (RaoQ): RaoQ is a multidimensional functional dispersion used to measure diversity and difference within and between populations [33]. Calculation of RaoQ involved first obtaining the matrix of species eigenvalues and, second, calculating the relative abundance of species in different plots [33]:where, is the distance between species i and j, is the abundance of species i, n is the total number of traits, and uij is the number of traits with different traits values of species i and j.
2.4.2. Species Diversity Indices
2.5. Statistical Analyses
3. Results
3.1. Functional Diversity and Species Diversity of Different Plant Habits
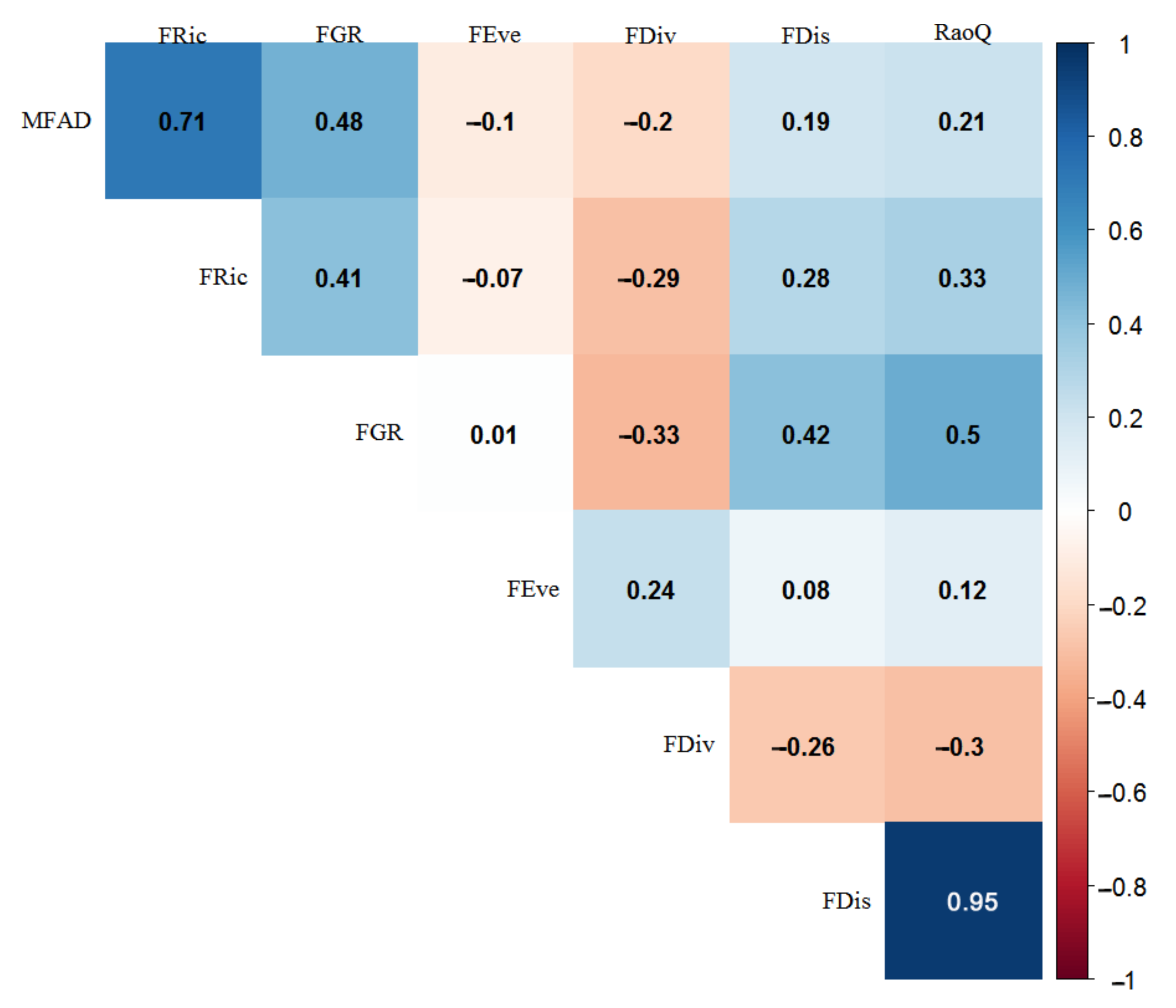
3.2. Correlation Analysis between Functional Indices
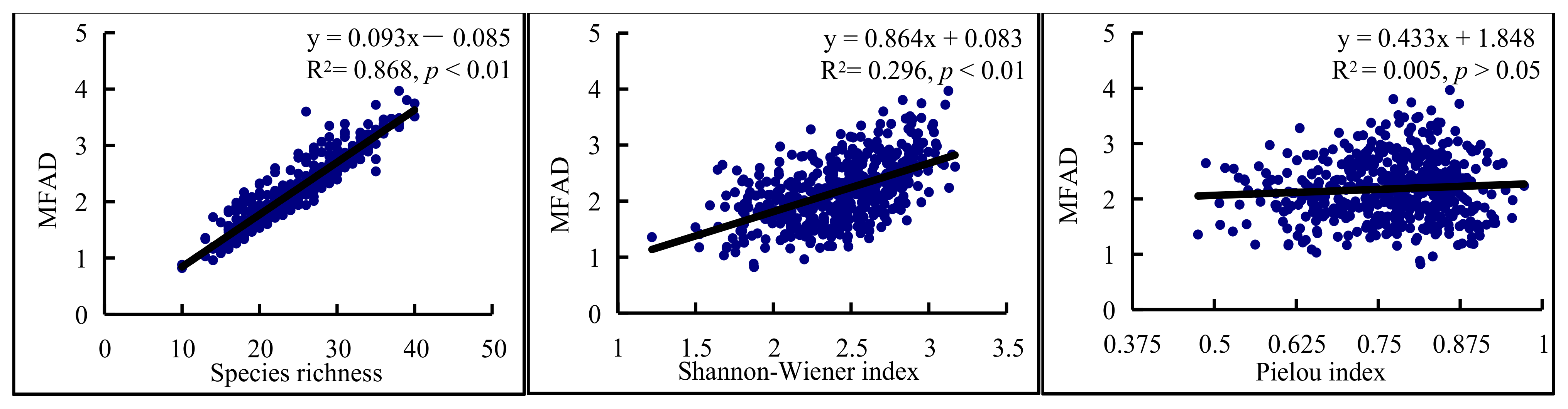
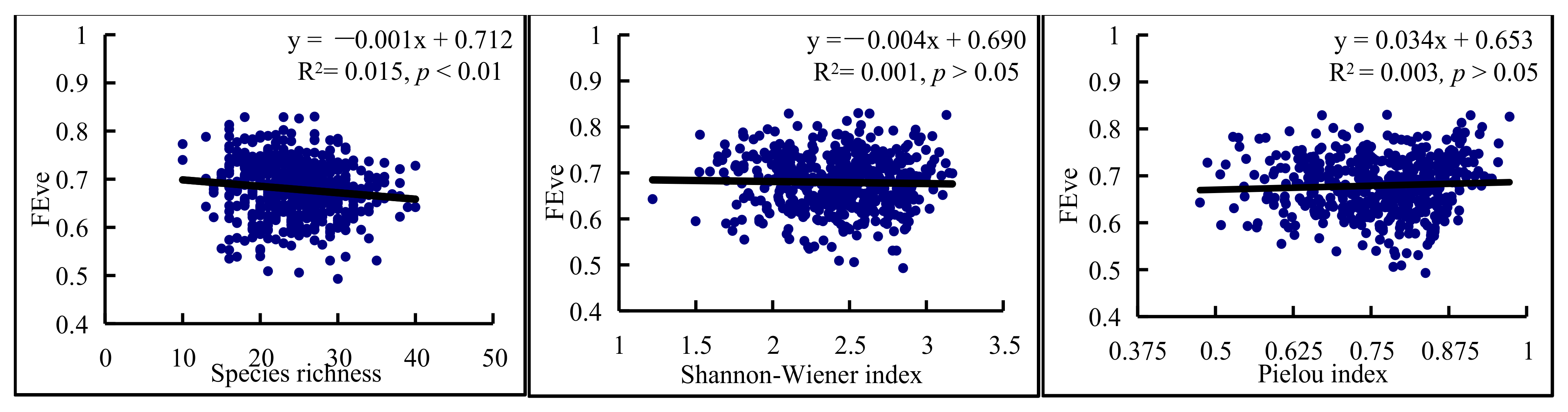
3.3. Relationships between Functional and Species Diversity
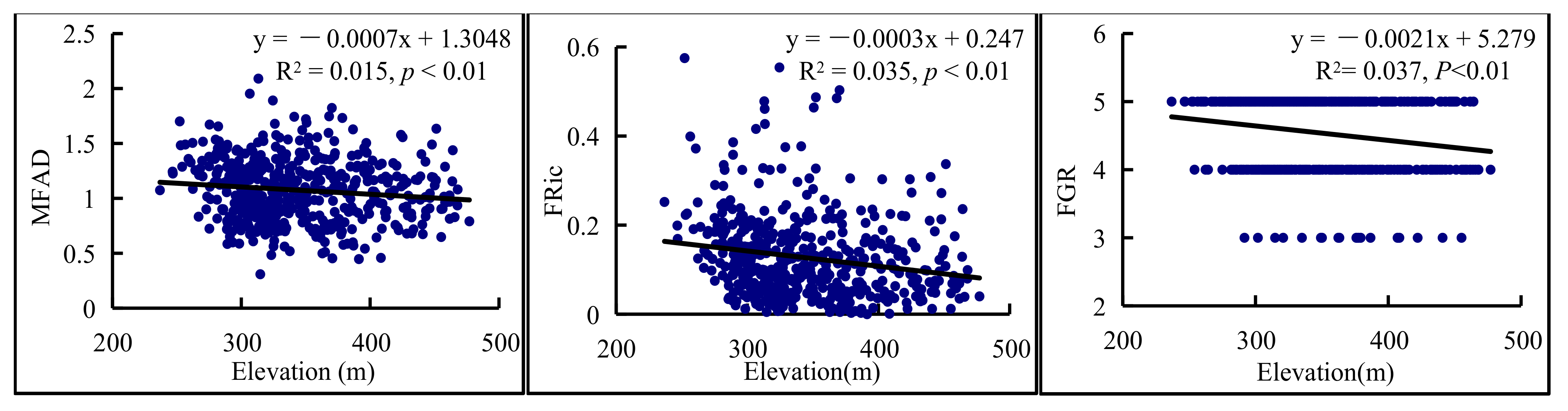
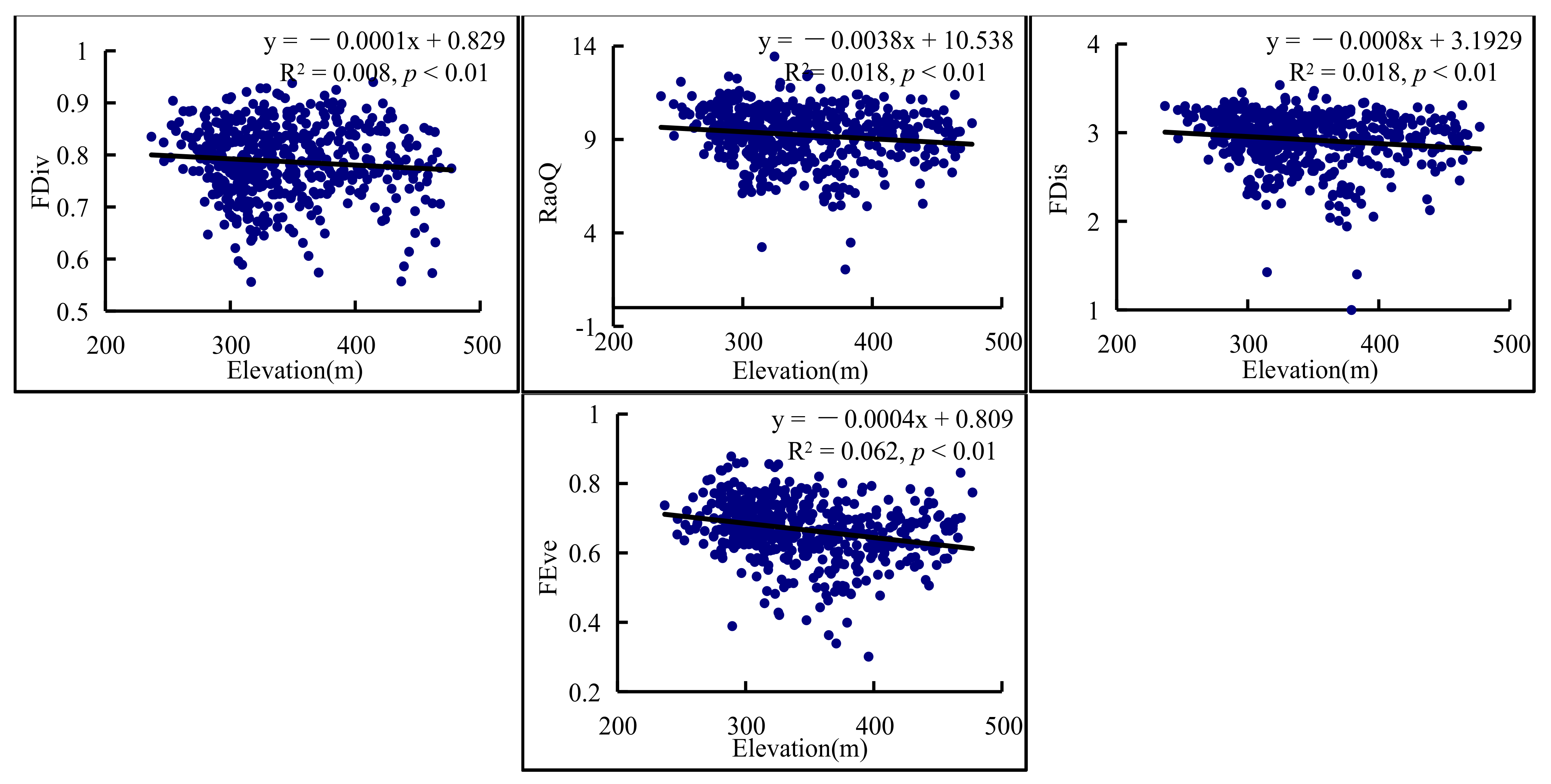
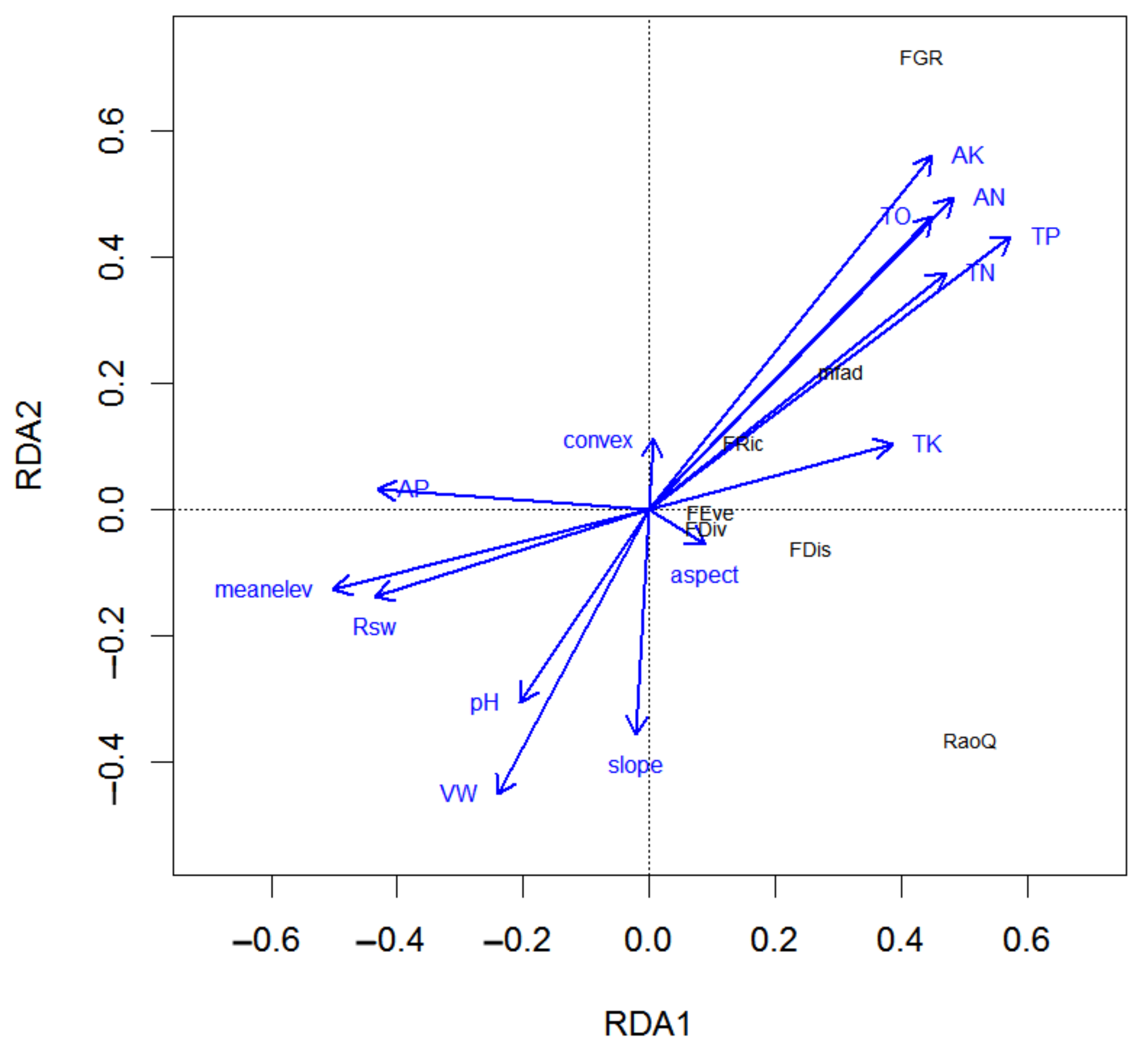
3.4. Environmental Factors Variation in Functional Diversity Indices
4. Discussions
4.1. Differences in Functional Diversity between Plant Habits
4.2. Relationships between Functional and Species Diversity
4.3. Functional Diversity Was Influenced by Environmental Effects
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schleuter, D.; Daufresne, M.; Massol, F.; Argillier, C. A user’s guide to functional diversity indices. Ecol. Monogr. 2010, 80, 469–484. [Google Scholar] [CrossRef] [Green Version]
- Mouchet, M.; Guilhaumon, F.; Villeger, S.; Mason, N.W.H.; Tomasini, J.A.; Mouillot, D. Towards a consensus for calculating dendrogram-based functional diversity indices. Oikos 2008, 117, 794–800. [Google Scholar] [CrossRef]
- Petchey, O.; Gaston, K. Dendrograms and measuring functional diversity. Oikos 2007, 116, 1422–1426. [Google Scholar] [CrossRef]
- Pimiento, C.; Leprieur, F.; Silvestro, D.; Lefcheck, J.S.; Albouy, C.; Rasher, D.B.; Davis, M.; Svenning, J.C.; Griffin, J.N. Functional diversity of marine megafauna in the Anthropocene. Sci. Adv. 2020, 6, eaay7650. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Knops, J.; Wedin, D.; Reich, P.; Ritchie, M.; Siemann, E. The influence of functional diversity and composition on ecosystem processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef] [Green Version]
- Kuebbing, S.E.; Maynard, D.S.; Bradford, M.A. Linking functional diversity and ecosystem processes: A framework for using functional diversity metrics to predict the ecosystem impact of functionally unique species. J. Ecol. 2018, 106, 687–698. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Functional diversity: Back to basics and looking forward. Ecol. Lett. 2006, 9, 741–758. [Google Scholar] [CrossRef]
- Mason, N.W.H.; Mouillot, D.; Lee, W.G.; Wilson, J.B. Functional richness, functional evenness and functional divergence: The primary components of functional diversity. Oikos 2005, 111, 112–118. [Google Scholar] [CrossRef]
- Walker, B.; Kinzig, A.; Langridge, J. Plant Attribute Diversity, Resilience, and Ecosystem Function: The Nature and Significance of Dominant and Minor Species. Ecosystems 1999, 2, 95–113. [Google Scholar] [CrossRef]
- Petchey, O.L.; Gaston, K.J. Extinction and the loss of functional diversity. Proc. R. Soc. B Biol. Sci. 2002, 269, 1721–1727. [Google Scholar] [CrossRef] [Green Version]
- Mason, N.W.H.; MacGillivray, K.; Steel, J.B.; Wilson, J.B. An index of functional diversity. J. Veg. Sci. 2003, 14, 571–578. [Google Scholar] [CrossRef]
- Ricotta, C. A note on functional diversity measures. Basic Appl. Ecol. 2005, 6, 479–486. [Google Scholar] [CrossRef]
- Mouillot, D.; Mason, W.H.N.; Dumay, O.; Wilson, J.B. Functional regularity: A neglected aspect of functional diversity. Oecologia 2005, 142, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Podani, J.; Schmera, D. On dendrogram-based measures of functional diversity. Oikos 2006, 115, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Garnier, E.; Navas, M.L.; Grigulis, K. Plant Functional Diversity: Organism Traits, Community Structure, and Ecosystem Properties; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Zhang, H.; Chen, H.Y.H.; Lian, J.Y.; John, R.; Li, R.H.; Liu, H.; Ye, W.H.; Berninger, F.; Ye, Q. Using functional trait diversity patterns to disentangle the scale-dependent ecological processes in a subtropical forest. Funct. Ecol. 2018, 32, 1379–1389. [Google Scholar] [CrossRef]
- Yang, X.X.; Li, M.Q.; He, X.D. Effects of Functional Diversity on Species Diversity of Plant Communities in Sandy Land. Acta Sci. Nat. Univ. Nankaiensis 2020, 53, 75–80. [Google Scholar]
- Funk, J.L.; Larson, J.E.; Ames, G.M.; Butterfield, B.J.; Cavender-Bares, J.; Firn, J.; Laughlin, D.C.; Sutton-Grier, A.E.; Williams, L.; Wright, J. Revisiting the Holy Grail: Using plant functional traits to understand ecological processes. Biol. Rev. 2017, 92, 1156–1173. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef]
- Butterfield, B.J.; Callaway, R.M. A functional comparative approach to facilitation and its context dependence. Funct. Ecol. 2013, 27, 907–917. [Google Scholar] [CrossRef]
- Shi, P.; Preisler, H.K.; Quinn, B.K.; Zhao, J.; Huang, W.; Röll, A.; Cheng, X.; Li, H.; Hölscher, D. Precipitation is the most crucial factor determining the distribution of moso bamboo in Mainland China. Global Ecology and Conservation. Glob. Ecol. Conserv. 2020, 22, e00924. [Google Scholar] [CrossRef]
- Duran, S.M.; Martin, R.E.; Diaz, S.; Maitner, B.S.; Malhi, Y.; Salinas, N.; Shenkin, A.; Silman, M.R.; Wieczynski, D.J.; Asner, G.P.; et al. Informing trait-based ecology by assessing remotely sensed functional diversity across a broad tropical temperature gradient. Sci. Adv. 2019, 5, eaaw8114. [Google Scholar] [CrossRef] [Green Version]
- Schrader, J.; Shi, P.J.; Royer, D.L.; Peppe, D.J.; Gallagher, R.V.; Li, Y.R.; Wang, R.; Wright, I.J. Leaf size estimation based on leaf length, width and shape. Ann. Bot. 2021, 128, 395–406. [Google Scholar] [CrossRef]
- Yu, X.; Shi, P.; Schrader, J.; Niklas, K.J. Nondestructive estimation of leaf area for 15 species of vines with different leaf shapes. Am. J. Bot. 2020, 107, 1481–1490. [Google Scholar] [CrossRef]
- Lin, G.; Stralberg, D.; Gong, G.; Huang, Z. Separating the Effects of Environment and Space on Tree Species Distribution: From Population to Community. PLoS ONE 2013, 8, e56171. [Google Scholar] [CrossRef] [Green Version]
- Condit, R.; Engelbrecht, B.M.J.; Pino, D.; Pérez, R.; Turner, B.L. Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical trees. Proc. Natl. Acad. Sci. USA 2013, 110, 5064–5068. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Ye, W.; Cao, H.; Huang, Z.; Lian, J.; Li, L.; Wei, S.; Sun, I.-F. Species-topography association in a species-rich subtropical forest of China. Basic Appl. Ecol. 2009, 10, 648–655. [Google Scholar] [CrossRef]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.F.; He, F. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Santiago, L.S.; Shen, H.; Ma, L.; Lian, J.Y.; Cao, H.L.; Lu, H.P.; Ye, W.H. Determinants of change in subtropical tree diameter growth with ontogenetic stage. Oecologia 2014, 175, 1315–1324. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Yu, S.X.; Lian, J.Y.; Shen, H.; Cao, H.L.; Lu, H.P.; Ye, W.H. Tree aboveground carbon storage correlates with environmental gradients and functional diversity in a tropical forest. Sci. Rep. 2016, 6, 25304. [Google Scholar] [CrossRef] [Green Version]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E.; Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Botta-Dukát, Z. Rao’s quadratic entropy as a measure of functional diversity based on multiple traits. J. Veg. Sci. 2005, 16, 533–540. [Google Scholar] [CrossRef]
- Lajoie, G.; Vellend, M. Understanding context dependence in the contribution of intraspecific variation to community trait–environment matching. Ecology 2016, 96, 2912–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, Y.; He, X.M.; Yang, X.D.; Lü, G.H. Responses of plant functional trait and diversity to soil water and salinity changes in desert ecosystem. Acta Ecol. Sin. 2019, 39, 1541–1550. [Google Scholar]
- Hooper, D.U.; Buchmann, N.; Degrange, V.; Díaz, S.M.; Spehn, E.M. Species diversity, functional diversity and ecosystem functioning. In Biodiversity and Ecosystem Functioning: Synthesis and Perspectives; Loreau, M., Naeem, S., Inchausti, P., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 195–208. [Google Scholar]
- Xue, Q.N.; Yan, M.; Bi, R.C. Functional Diversity Research of Tree and Shrub Layers in Forest Communities of the Wulu Mountains Nature Reserve in Shanxi, China. Acta Ecol. Sin. 2015, 35, 7023–7032. [Google Scholar]
- Dong, S.K.; Tang, L.; Zhang, X.F.; Liu, S.L.; Liu, Q.R.; Su, X.K.; Zhang, Y.; Wu, X.Y.; Zhao, Z.; Li, Y.; et al. Relationship between plant species diversity and functional diversity in alpine grasslands. Acta Ecol. Sin. 2017, 37, 1472–1483. [Google Scholar]
- Chen, C.; Zhu, Z.H.; Li, Y.N.; Yao, T.H.; Pan, S.Y.; Wei, X.H.; Kong, B.B.; Du, J.L. Effects of interspecific trait dissimilarity and species evenness on the relationship between species diversity and functional diversity in an alpine meadow. Acta Ecol. Sin. 2016, 36, 661–674. [Google Scholar]
- Liu, M.X.; Nan, X.N.; Zhang, G.J. Relationship between species diversity and functional diversity of plant communities on different slopes in alpine meadow. Acta Ecol. Sin. 2021, 41, 5398–5407. [Google Scholar]
- Wu, C.C.; Tsui, C.C.; Hseih, C.F.; Asio, V.B.; Chen, Z.S. Mineral nutrient status of tree species in relation to environmental factors in the subtropical rain forest of Taiwan. For. Ecol. Manag. 2007, 239, 81–91. [Google Scholar] [CrossRef]
- Ding, J.; Wu, Q.; Yan, H.; Zhang, S. Effects of topographic variations and soil characteristics on plant functional traits in a subtropical evergreen broad-leaved forest. Biodivers. Sci. 2011, 19, 158–167. [Google Scholar]
- Zhou, G.X.; Huang, L.X.; Zang, X.W.; Wei, X.; Ye, W.H.; Shen, H. Effects of Habitat Heterogeneity on Community Functional Diversity of Dinghu Mountain Evergreen Broad-leaved Forest. Guihaia 2016, 36, 127–136. [Google Scholar]
- Qin, H.; Zhang, Y.B.; Dong, G.; Zhang, F. Altitudinal patterns of taxonomic, phylogenetic and functional diversity of forest communities in Mount Guandi, Shanxi, China. Chin. J. Plant Ecol. 2019, 43, 762–773. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Column Header | Description | Performance |
|---|---|---|
| Nmass | Leaf nitrogen content per unit mass (g g−1) | Nitrogen economy of leaves |
| Pmass | Leaf phosphorus content per unit mass (g g−1) | Phosphorus economy of leaves |
| SLA | Specific leaf area (cm2 g−1) | Carbon economy of leaves |
| Gs | Stomatal conductance per unit mass (mmol g−1 s−1) | Light capture strategy |
| LA | Leaf area(cm2) | Light capture strategy |
| Pl | Petiole length (cm) | Light capture strategy |
| Chl | Leaf chlorophyll concentration (g m−2) | Light capture strategy |
| PNUE | Photosynthetic nitrogenuse efficiency (μmol mol−1 s−1) | Light capture strategy |
| PPUE | Photosynthetic phosphorus use efficiency (μmol mol−1 s−1) | Light capture strategy |
| Pdm | Petiole dry matter (mg kg−1) | Light capture strategy |
| WD | Sapwood density (g cm−3) | Hydraulic conductivity |
| Kl | Leaf-specific conductivity (kg m−1 s−1 MPa−1) | Hydraulic conductivity |
| Ks | Sapwood-specific conductivity (kg m−1 s−1 MPa−1) | Hydraulic conductivity |
| Pd | Petiole density (g cm−2) | Hydraulic conductivity |
| LS | Leaf shape: oval, long elliptic, lanceolate, palmate leaf | Ecological adaptability |
| Plant habit | Three forms: large tree, small tree, and shrub | Ecological adaptability |
| Seed Dispersal Mode | Three modes: dispersal by animals, dispersal by wind, and dispersal both by animals and wind | Ecological adaptability |
| NO. | Abbreviation | Description |
|---|---|---|
| 1 | AK | Rapidly available potassium |
| 2 | AN | Soil available nitrogen |
| 3 | AP | Soil available phosphorus |
| 4 | TO | Soil organic matter |
| 5 | TP | Soil total phosphorus |
| 6 | TK | Soil total potassium |
| 7 | TN | Soil total nitrogen |
| 8 | VW | Volume weight of soil |
| 9 | RSW | Soil moisture content |
| 10 | pH | Soil pH |
| 11 | Aspect | Aspect |
| 12 | Meanelev | Average elevation |
| 13 | Convex | Convex |
| 14 | Slope | Slope |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Wen, Z.; Wei, S.; Lian, J.; Ye, W. Functional Diversity and Its Influencing Factors in a Subtropical Forest Community in China. Forests 2022, 13, 966. https://doi.org/10.3390/f13070966
Li L, Wen Z, Wei S, Lian J, Ye W. Functional Diversity and Its Influencing Factors in a Subtropical Forest Community in China. Forests. 2022; 13(7):966. https://doi.org/10.3390/f13070966
Chicago/Turabian StyleLi, Lin, Zhifeng Wen, Shiguang Wei, Juyu Lian, and Wanhui Ye. 2022. "Functional Diversity and Its Influencing Factors in a Subtropical Forest Community in China" Forests 13, no. 7: 966. https://doi.org/10.3390/f13070966
APA StyleLi, L., Wen, Z., Wei, S., Lian, J., & Ye, W. (2022). Functional Diversity and Its Influencing Factors in a Subtropical Forest Community in China. Forests, 13(7), 966. https://doi.org/10.3390/f13070966