
Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation


Abstract
:1. Introduction
2. Materials and Methods
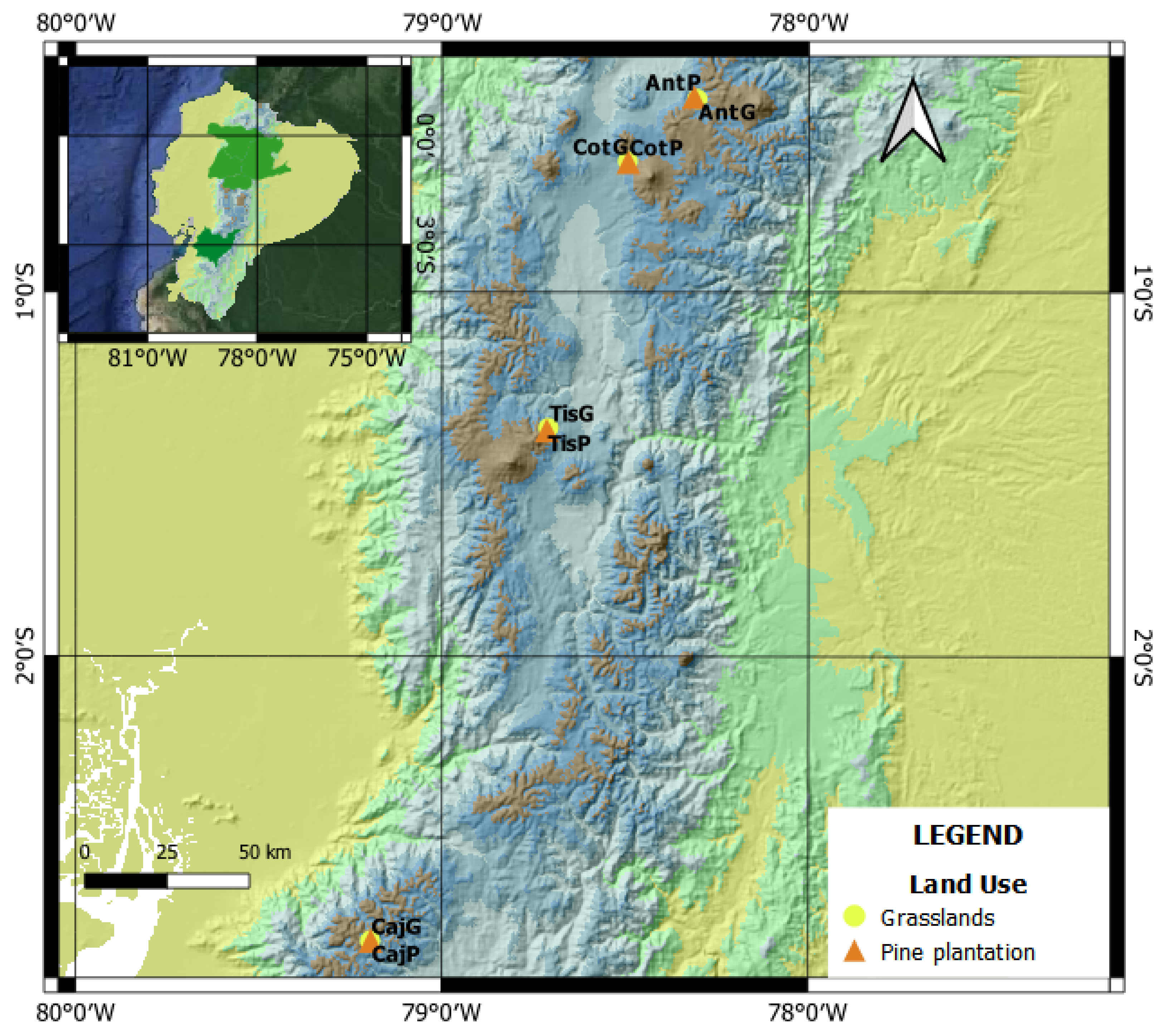
2.1. Study Zones
2.2. Experimental Design and Data Collection
2.3. Data Analysis
3. Results
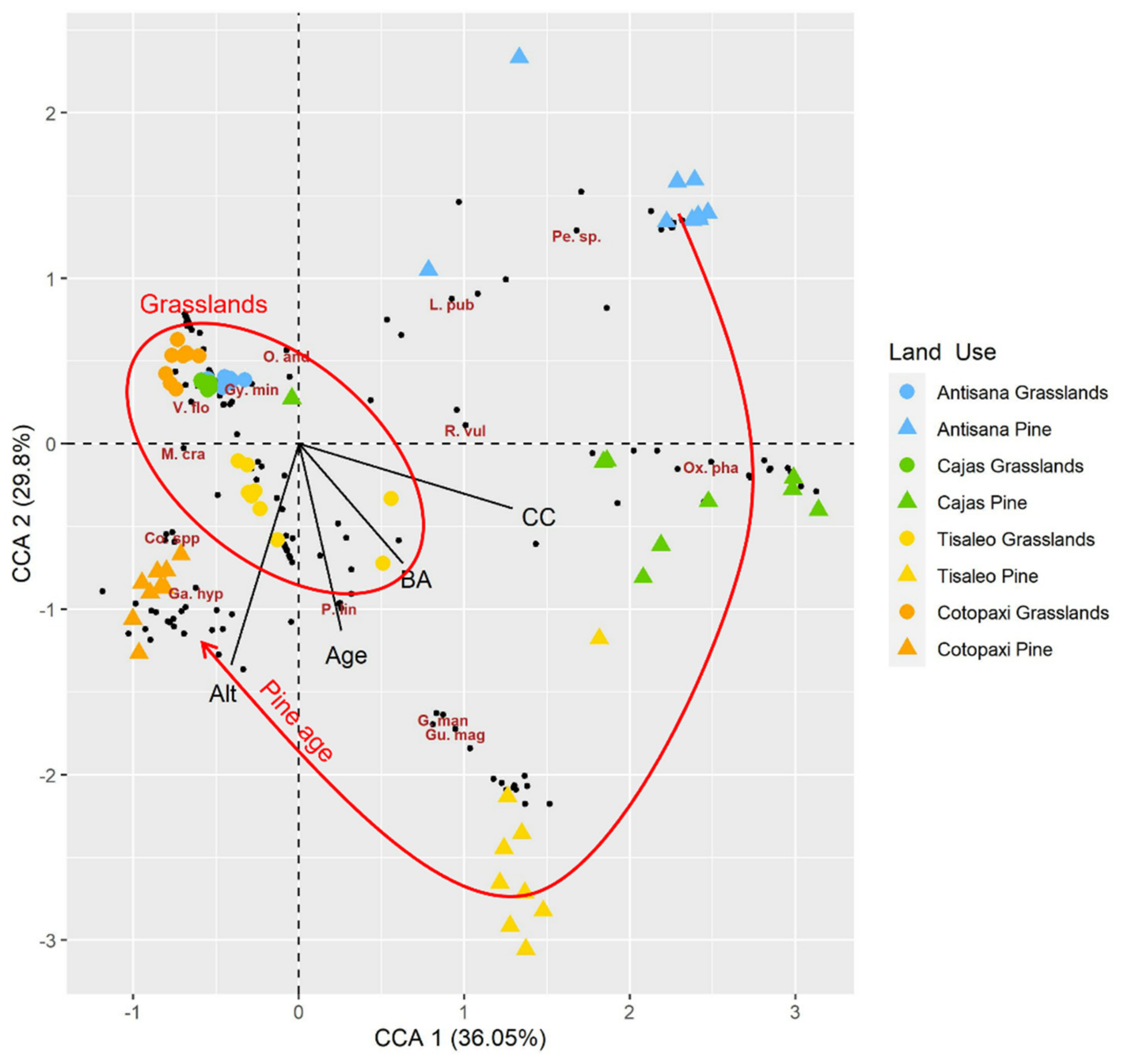
3.1. Multivariate Analysis (CCA) Illustrating Pine Plantation Effects on Plant Community
3.2. Land Use and Zone Effects on Plant Cover and Species Richness
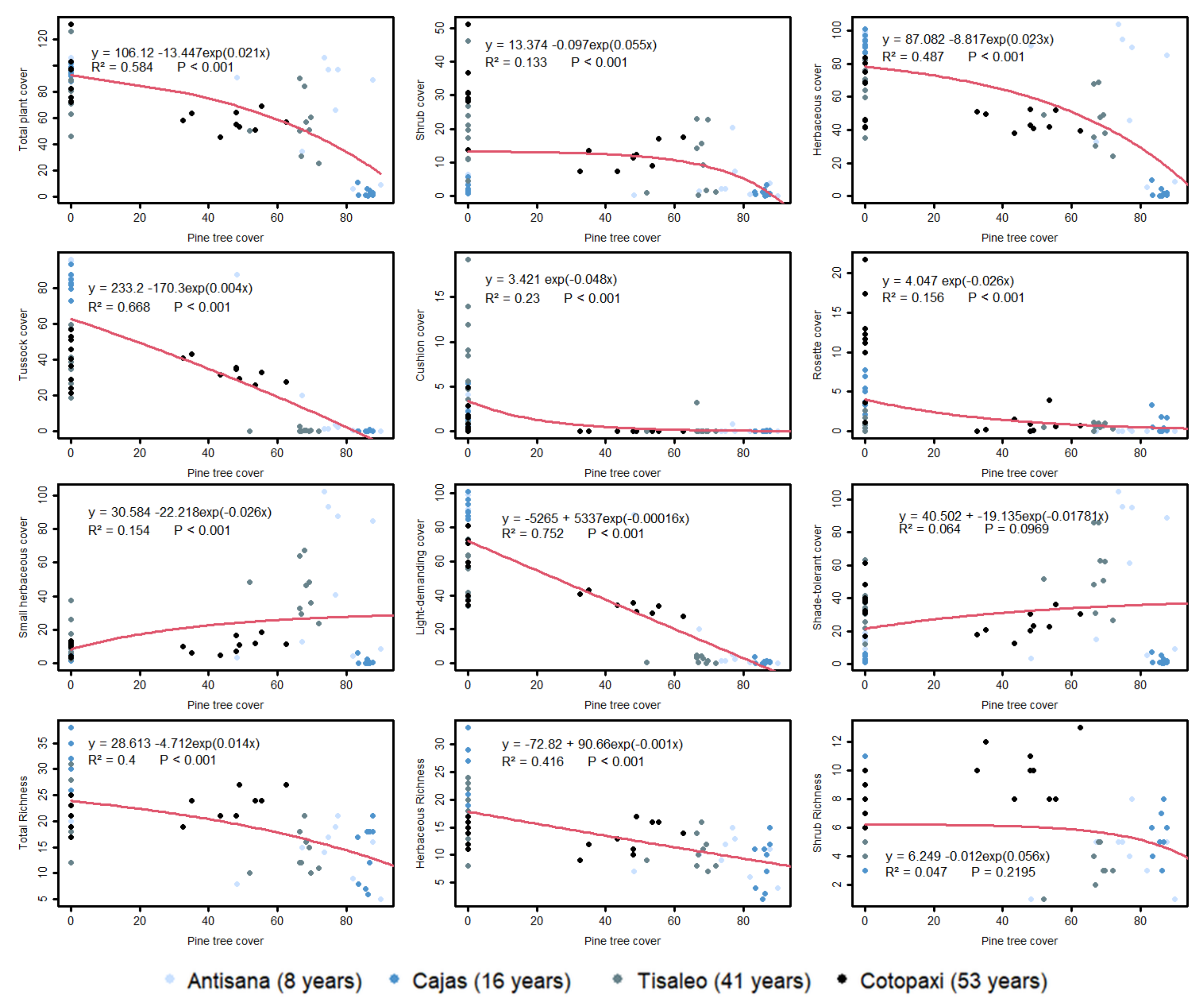
3.3. Relationships between Pine Cover and Plant Cover/Richness
3.4. Indicator Species of the Study Zone and of the Pine Effect
4. Discussion
4.1. What Is the Impact of Pine Plantations on the Cover and Diversity of Understory Species?
4.2. Does the Zone/Age Influence the Pine Plantation Impact on Páramo Vegetation?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Antisana | Cajas | Tisaleo | Cotopaxi | Shade | Life | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Family | Species | Grasslands | Pine | Grasslands | Pine | Grasslands | Pine | Grasslands | Pine | Tolerance | Form |
| Poaceae | Calamagrostis intermedia | 86.3 | 1.9 | 83.7 | 0.1 | 37.3 | 0.2 | 28.4 | 6.7 | LD | T |
| Rosaceae | Lachemilla orbiculata | 1.9 | 42.9 | <0.1 | 0.2 | 3.4 | 5 | 0.9 | 0.5 | ST | H |
| Poaceae | Cortaderia jubata | 7.8 | 18.3 | LD | T | ||||||
| Gunneraceae | Gunnera magellanica | 4.9 | 14.4 | ST | H | ||||||
| Plantaginaceae | Sibthorpia repens | <0.1 | <0.1 | 12 | ST | H | |||||
| Poaceae | Agrostis breviculmis | <0.1 | 11 | LD | T | ||||||
| Bromeliaceae | Puya hamata | <0.1 | 10.7 | 0.2 | LD | R | |||||
| Ericaceae | Disterigma empetrifolium | 0.3 | 0.6 | 0.1 | 8.9 | 0.1 | ST | S | |||
| Rosaceae | Alchemilla hirta | 0.2 | 0.1 | 2.9 | 8.8 | ST | H | ||||
| Asteraceae | Baccharis caespitosa | <0.1 | 7.8 | LD | C | ||||||
| Asteraceae | Loricaria ilinissae | 0.4 | 7.5 | ST | S | ||||||
| Asteraceae | Diplostephium ericoidese | 6.3 | ST | S | |||||||
| Ericaceae | Gaultheria myrsinoides | 0.7 | <0.1 | <0.1 | 4.3 | 1.4 | 5.4 | 0.3 | ST | S | |
| Poaceae | Cortaderia nitida | 2.2 | 5.3 | LD | T | ||||||
| Hypericaceae | Hypericum laricifolium | 0.1 | 0.1 | 2.5 | 4.3 | <0.1 | ST | S | |||
| Asteraceae | Aristeguietia sp. | 4.1 | ST | S | |||||||
| Caprifoliaceae | Valeriana microphylla | 2.7 | 0.5 | 3.7 | 2.4 | 0.1 | 0.1 | ST | S | ||
| Ericaceae | Vaccinium floribundum | <0.1 | 0.1 | 3.6 | 1.3 | ST | S | ||||
| Poaceae | Paspalum bonplandianum | 3.4 | 0.1 | LD | T | ||||||
| Cyperaceae | Carex polystachya | 1.5 | 3.3 | LD | T | ||||||
| Geraniaceae | Geranium reptans | 2.9 | 3 | 0.1 | <0.1 | 0.4 | 0.6 | ST | H | ||
| Asteraceae | Werneria nubigena | 0.2 | 2.8 | 0.1 | 0.2 | LD | R | ||||
| Poaceae | Cortaderia sericantha | 2.8 | LD | T | |||||||
| Lycopodiaceae | Lycopodium magellanicum | 2.8 | ST | H | |||||||
| Asteraceae | Senecio tephrosioides | 2.5 | ST | H | |||||||
| Rubiaceae | Galium hypocarpium | <0.1 | <0.1 | 0.3 | 2.4 | ST | H | ||||
| Asteraceae | Baccharis teindalensis | 2.4 | <0.1 | ST | S | ||||||
| Melastomataceae | Brachyotum ledifolium | 0.1 | 2.2 | ST | S | ||||||
| Rubiaceae | Arcytophyllum nitidum | 2.1 | ST | S | |||||||
| Geraniaceae | Geranium maniculatum | <0.1 | 0.9 | 2.1 | ST | H | |||||
| Rosaceae | Rubus coriaceus | 0.2 | 2 | ST | S | ||||||
| Rosaceae | Rubus nubigenus | 1.8 | ST | S | |||||||
| Rubiaceae | Nertera granadensis | 0.4 | 0.1 | 0.2 | 1.8 | ST | H | ||||
| Rosaceae | Margyricarpus pinnatus | 1.8 | ST | S | |||||||
| Geraniaceae | Geranium antisanaee | 1.4 | 1.7 | ST | H | ||||||
| Apiaceae | Azorella biloba | 1.5 | LD | C | |||||||
| Apiaceae | Azorella pedunculata | 1.4 | 0.1 | 0.2 | 0.6 | 1.4 | LD | C | |||
| Cyatheaceae | Cyathea xenoxyla | 1.4 | ST | S | |||||||
| Rosaceae | Polylepis sp. | <0.1 | 1.3 | ST | S | ||||||
| Lycopodiaceae | Lycopodium clavatum | 0.1 | 1.2 | ST | H | ||||||
| Gentianaceae | Gentianella cerastioides | 0.1 | <0.1 | 1.2 | LD | H | |||||
| Solanaceae | Solanum stenophyllum | 1 | ST | S | |||||||
| Cyperaceae | Carex sp.1 | 1 | 0.3 | LD | T | ||||||
| Lycopodiaceae | Phlegmariurus crassus | 0.3 | <0.1 | 1 | ST | H | |||||
| Alstroemeriaceae | Bomarea multiflora | 0.9 | ST | H | |||||||
| Asteraceae | Aristeguietia lamiifolia | 0.8 | ST | S | |||||||
| Caprifoliaceae | Aretiastrum aretioidese | 0.8 | LD | C | |||||||
| Poaceae | Bromus lanatus | 0.7 | <0.1 | 0.4 | LD | H | |||||
| Dryopteridaceae | Elaphoglossum engelii | 0.7 | 0.2 | ST | H | ||||||
| Araliaceae | Hydrocotyle bonplandii | 0.6 | 0.2 | <0.1 | 0.7 | 0.1 | ST | H | |||
| Rosaceae | Acaena ovalifolia | 0.2 | 0.7 | ST | H | ||||||
| Rosaceae | Lachemilla hispidula | <0.1 | 0.6 | ST | H | ||||||
| Grossulariaceae | Ribes lehmanniie | <0.1 | 0.6 | ST | S | ||||||
| Cyperaceae | Rhynchospora vulcani | 0.6 | 0.5 | <0.1 | LD | R | |||||
| Cyperaceae | Carex sp. 2 | 0.5 | LD | T | |||||||
| Asteraceae | Senecio sp. | 0.5 | ST | H | |||||||
| Asteraceae | Galinsoga parviflora | <0.1 | 0.5 | ST | H | ||||||
| Apiaceae | Eryngium humile | <0.1 | 0.5 | 0.1 | LD | R | |||||
| Caryophyllaceae | Arenaria serpyllifolia | 0.1 | 0.1 | <0.1 | 0.5 | ST | H | ||||
| Lamiaceae | Clinopodium nubigenum | 0.5 | 0.3 | 0.2 | LD | H | |||||
| Asteraceae | Hieracium frigidum | <0.1 | <0.1 | 0.5 | <0.1 | ST | R | ||||
| Piperaceae | Peperomia sp. | 0.1 | 0.5 | ST | H | ||||||
| Lycopodiaceae | Austrolycopodium magellanicum | 0.4 | <0.1 | 0.4 | LD | H | |||||
| Berberidaceae | Berberis rigida | 0.1 | 0.1 | 0.4 | 0.2 | ST | S | ||||
| Geraniaceae | Geranium sibbaldioides | 0.4 | <0.1 | ST | H | ||||||
| Cyperaceae | Uncinia hamata | 0.4 | 0.3 | LD | R | ||||||
| Plantaginaceae | Plantago rigida | 0.4 | LD | C | |||||||
| Melastomataceae | Brachyotum jamesoniie | 0.1 | 0.4 | ST | S | ||||||
| Grossulariaceae | Ribes ecuadorense | 0.3 | ST | S | |||||||
| Cyperaceae | Uncinia tenuis | 0.3 | <0.1 | 0.3 | <0.1 | LD | R | ||||
| Gentianaceae | Halenia weddelliana | 0.3 | <0.1 | 0.1 | LD | H | |||||
| Asteraceae | Taraxacum campylodes | 0.3 | ST | H | |||||||
| Caryophyllaceae | Cerastium mollissimum | 0.3 | ST | H | |||||||
| Dennstaedtiaceae | Pteridium arachnoideum | 0.3 | ST | R | |||||||
| Asteraceae | Gynoxys buxifolia | 0.3 | LD | S | |||||||
| Polygalaceae | Monnina crassifolia | 0.3 | 0.2 | ST | S | ||||||
| Asteraceae | Gamochaeta cf. humilis | 0.3 | LD | H | |||||||
| Asteraceae | Bidens andicola | 0.2 | LD | H | |||||||
| Hypericaceae | Hypericum lancioides | 0.2 | ST | S | |||||||
| Cyperaceae | Rhynchospora sp. | 0.2 | LD | R | |||||||
| Unidentified | Unknown 3 | <0.1 | 0.2 | ST | R | ||||||
| Apiaceae | Azorella aretioides | <0.1 | 0.2 | LD | C | ||||||
| Asteraceae | Bidens pilosa | 0.2 | LD | H | |||||||
| Lamiaceae | Salvia pichinchensis | 0.2 | LD | S | |||||||
| Asteraceae | Gynoxys miniphyllae | 0.2 | <0.1 | LD | S | ||||||
| Aspleniaceae | Asplenium monanthes | 0.2 | ST | R | |||||||
| Melastomataceae | Miconia chionophila | 0.2 | ST | S | |||||||
| Blechnaceae | Blechnum loxense | 0.1 | 0.2 | ST | R | ||||||
| Polypodiaceae | Polypodium mindensee | 0.2 | ST | H | |||||||
| Asteraceae | Diplostephium sp. | 0.2 | 0.1 | ST | S | ||||||
| Poaceae | Calamagrostis ligulata | 0.2 | LD | T | |||||||
| Calceolariaceae | Calceolaria crenata | 0.2 | ST | S | |||||||
| Plantaginaceae | Plantago linearis | 0.2 | <0.1 | LD | T | ||||||
| Asteraceae | Ageratina dendroidese | 0.2 | ST | S | |||||||
| Hypericaceae | Hypericum decandrum | 0.2 | <0.1 | ST | S | ||||||
| Iridaceae | Orthrosanthus chimboracensis | 0.1 | 0.1 | LD | H | ||||||
| Ranunculaceae | Ranunculus praemorsus | 0.1 | <0.1 | LD | H | ||||||
| Asteraceae | Pentacalia peruviana | 0.1 | LD | S | |||||||
| Aspleniaceae | Unknown 2 | <0.1 | 0.1 | ST | H | ||||||
| Apiaceae | Oreomyrrhis andicola | 0.1 | <0.1 | <0.1 | ST | H | |||||
| Asteraceae | Dorobaea pimpinellifolia | <0.1 | 0.1 | <0.1 | LD | H | |||||
| Geraniaceae | Geranium multipartitum | 0.1 | ST | H | |||||||
| Poaceae | Agrostis sp. 1 | 0.1 | LD | H | |||||||
| Rubiaceae | Arcytophyllum filiforme | 0.1 | ST | S | |||||||
| Aspleniaceae | Asplenium cuspidatum | 0.1 | ST | R | |||||||
| Asteraceae | Gynoxys sp. | 0.1 | <0.1 | LD | S | ||||||
| Blechnaceae | Blechnum auratum | 0.1 | ST | R | |||||||
| Cyatheaceae | Cyathea caracasana | 0.1 | ST | S | |||||||
| Asteraceae | Gynoxys halliie | 0.1 | LD | S | |||||||
| Escalloniaceae | Escallonia myrtilloides | 0.1 | <0.1 | LD | S | ||||||
| Pteridaceae | Jamesonia alstonii | 0.1 | ST | H | |||||||
| Hypericaceae | Hypericum aciculare | 0.1 | ST | S | |||||||
| Asteraceae | Hypochaeris sessiliflora | 0.1 | 0.1 | ST | R | ||||||
| Rosaceae | Lachemilla nivalis | 0.1 | LD | H | |||||||
| Bromeliaceae | Puya clava-herculis | <0.1 | 0.1 | LD | R | ||||||
| Asteraceae | Monticalia myrsinitese | 0.1 | <0.1 | LD | S | ||||||
| Asteraceae | Loricaria azuayensise | 0.1 | ST | S | |||||||
| Asteraceae | Erigeron ecuadoriensis | 0.1 | <0.1 | LD | H | ||||||
| Dryopteridaceae | Polystichum orbiculatum | 0.1 | ST | H | |||||||
| Melastomataceae | Brachyotum sp. | 0.1 | <0.1 | ST | S | ||||||
| Melastomataceae | Miconia salicifolia | 0.1 | ST | S | |||||||
| Asteraceae | Gamochaeta americana | <0.1 | <0.1 | 0.1 | LD | H | |||||
| Ophioglossaceae | Ophioglossum crotalophoroides | 0.1 | <0.1 | LD | H | ||||||
| Poaceae | Festuca andicola | 0.1 | LD | T | |||||||
| Rubiaceae | Galium corymbosum | 0.1 | ST | H | |||||||
| Asteraceae | Aphanactis ollgaardiie | <0.1 | LD | H | |||||||
| Gentianaceae | Halenia serpyllifoliae | <0.1 | LD | H | |||||||
| Oxalidaceae | Oxalis phaeotricha | <0.1 | <0.1 | LD | H | ||||||
| Dryopteridaceae | Polystichum sp. | <0.1 | ST | H | |||||||
| Fabaceae | Trifolium pratense | <0.1 | <0.1 | ST | H | ||||||
| Gentianaceae | Gentianella hyssopifoliae | <0.1 | ST | H | |||||||
| Polypodiaceae | Polypodium thyssanolepis | <0.1 | ST | H | |||||||
| Asteraceae | Cronquistianthus pseudoriganoidese | <0.1 | ST | S | |||||||
| Unidentified | Unknown 5 | <0.1 | ST | R | |||||||
| Plantaginaceae | Plantago australis | <0.1 | <0.1 | LD | H | ||||||
| Asteraceae | Monticalia arbutifolia | <0.1 | LD | S | |||||||
| Asteraceae | Baccharis genistelloides | <0.1 | ST | H | |||||||
| Cystopteridaceae | Cystopteris fragilis | <0.1 | <0.1 | ST | H | ||||||
| Dryopteridaceae | Elaphoglossum mathewsii | <0.1 | ST | H | |||||||
| Oxalidaceae | Oxalis lotoides | <0.1 | ST | H | |||||||
| Unidentified | Unknown 4 | <0.1 | ST | H | |||||||
| Aspleniaceae | Asplenium fragans | <0.1 | <0.1 | ST | R | ||||||
| Asteraceae | Baccharis tricuneata | <0.1 | <0.1 | LD | S | ||||||
| Apiaceae | Oreomyrrhis sp. | <0.1 | ST | H | |||||||
| Dryopteridaceae | Polystichum polyphyllum | <0.1 | ST | H | |||||||
| Fabaceae | Lupinus microphyllus | <0.1 | ST | H | |||||||
| Polygonaceae | Muehlenbeckia tamnifolia | <0.1 | ST | S | |||||||
| Asteraceae | Cotula mexicana | <0.1 | LD | H | |||||||
| Asteraceae | Gnaphalium dombeyanum | <0.1 | LD | H | |||||||
| Gentianaceae | Gentiana sedifolia | <0.1 | <0.1 | LD | H | ||||||
| Ranunculaceae | Ranunculus limoselloides | <0.1 | LD | H | |||||||
| Cyperaceae | Rhynchospora ruiziana | <0.1 | LD | R | |||||||
| Aspleniaceae | Asplenium sp. 2 | <0.1 | ST | H | |||||||
| Asteraceae | Achyrocline alata | <0.1 | ST | H | |||||||
| Asteraceae | Pluchea biformis | <0.1 | ST | H | |||||||
| Unidentified | Unknown 6 | <0.1 | ST | H | |||||||
| Ericaceae | Disterigma sp. 1 | <0.1 | ST | S | |||||||
| Fabaceae | Lupinus pubescens | <0.1 | <0.1 | ST | S | ||||||
| Aspleniaceae | Asplenium polyphyllum | <0.1 | ST | R | |||||||
| Asteraceae | Unknown 7 | <0.1 | ST | R | |||||||
| Apiaceae | Daucus montanus | LD | H | ||||||||
| Apiaceae | Niphogeton dissecta | LD | H | ||||||||
| Asteraceae | Taraxacum sp. 1 | ST | H | ||||||||
| Unidentified | Unknown 1 | ST | H | ||||||||
| Asteraceae | Aristeguietia cf. glutinosa | ST | S | ||||||||
| Asteraceae | Hypochaeris sonchoidese | ST | R | ||||||||
| Asteraceae | Ageratina sp. | ST | S | ||||||||
| Total cover (%) | 101.5 | 66.3 | 99 | 3.4 | 85 | 56.5 | 92.2 | 57.4 | |||
| Total species richness | 29 | 25 | 52 | 42 | 45 | 35 | 36 | 48 | |||
References
- FAO. Global Forest Resources Assessment 2015. Desk Reference; 1st ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; pp. 34–69. [Google Scholar]
- FAO. Global Forest Resources Assessment 2010. Desk Reference; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010; pp. 9–48. [Google Scholar]
- FAO. Global Planted Forests Thematic Study: Results and Analysis; Planted Forests and Trees Working Paper 38; Food and Agriculture Organization of the United Nations: Rome, Italy, 2006; pp. 13–28. [Google Scholar]
- Farley, K.A. Grasslands to Tree Plantations: Forest Transition in the Andes of Ecuador. Ann. Assoc. Am. Geogr. 2007, 97, 755–771. [Google Scholar] [CrossRef]
- Vanacker, V.; von Blanckenburg, F.; Govers, G.; Molina, A.; Poesen, J.; Deckers, J.; Kubik, P. Restoring Dense Vegetation Can Slow Mountain Erosion to near Natural Benchmark Levels. Geology 2007, 35, 303–306. [Google Scholar] [CrossRef]
- Geldenhuys, C. Native Forest Regeneration in Pine and Eucalypt Plantations in Northern Province, South Africa. For. Ecol. Manag. 1997, 99, 101–115. [Google Scholar] [CrossRef]
- Parrotta, J.A.; Turnbull, J.W.; Jones, N. Catalyzing Native Forest Regeneration on Degraded Tropical Lands. For. Ecol. Manag. 1997, 99, 1–7. [Google Scholar] [CrossRef]
- Parrotta, J.A. Influence of Overstory Composition on Understory Colonization by Native Species in Plantations on a Degraded Tropical Site. J. Veg. Sci. 1995, 6, 627–636. [Google Scholar] [CrossRef]
- Fimbel, R.A.; Fimbel, C.C. The Role of Exotic Conifer Plantations in Rehabilitating Degraded Tropical Forest Lands: A Case Study from the Kibale Forest in Uganda. For. Ecol. Manag. 1996, 81, 215–226. [Google Scholar] [CrossRef]
- Richardson, D.M.; Van Wilgen, B.W.; Nuñez, M.A. Alien Conifer Invasions in South America: Short Fuse Burning? Biol. Invasions 2008, 10, 573–577. [Google Scholar] [CrossRef]
- Simberloff, D.; Nuñez, M.A.; Ledgard, N.J.; Pauchard, A.; Richardson, D.M.; Sarasola, M.; Van Wilgen, B.W.; Zalba, S.M.; Zenni, R.D.; Bustamante, R.; et al. Spread and Impact of Introduced Conifers in South America: Lessons from Other Southern Hemisphere Regions. Austral Ecol. 2010, 35, 489–504. [Google Scholar] [CrossRef]
- Freeman, J.; Marques, C.M.P.; Carocha, V.; Borralho, N.; Potts, B.M.; Vaillancourt, R. Origins and Diversity of the Portuguese Landrace of Eucalyptus globulus. Ann. For. Sci. 2007, 64, 639–647. [Google Scholar] [CrossRef]
- Estades, C.F.; Escobar, M.A. los Ecosistemas de las Plantaciones de Pino de la Cordillera de la Costa. In Historia, Biodiversidad y Ecología de los Bosques Costeros de Chile; Smith-Ramírez, C., Armesto, J., Valdovinos, C., Eds.; Editorial Universitaria: Santiago de Chile, Chile, 2015; pp. 600–616. [Google Scholar]
- Carle, J.B.; Holmgren, L.P.B. Wood from Planted Forests: Global Outlook to 2030. Plant. For. Uses Impacts Sustain. 2009, 58, 47–59. [Google Scholar] [CrossRef]
- Payn, T.; Carnus, J.M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfield, M.J. Changes in Planted Forests and Future Global Implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef]
- Indústria Brasileira de Árvores. Report 2019; Brasilan Tree Industry: Brasilia, Brasil, 2019; pp. 32–41. [Google Scholar]
- Instituto Forestal. Anuario Forestal 2019; Área de información y economía del Instituto Forestal: Santiago de Chile, Chile, 2019; pp. 19–72. [Google Scholar]
- Ministerio de Agricultura Ganadería y Pesca de Argentina. Mapa de Plantaciones Forestales Promocionadas (MPF) de la República Argentina; Ministerio de Agricultura Ganadería y Pesca de Argentina: Buenos Aires, Argentina, 2018. [Google Scholar]
- Jonard, M.; Colmant, R.; Heylen, C.; Ysebaert, C.; Carton, C.; Picard, L.; Cassart, B.; Hounzandji, A.P.I.; Ponette, Q. Impact de Boisements Résineux Sur la Séquestration du Carbone dans les Andes Péruviennes: Cas des Plantations de Pinus patula dans la Coopérative de Granja Porcon (Cajamarca), Pérou. Bois. For. Trop. 2014, 68, 17–27. [Google Scholar] [CrossRef]
- Chávez, J.; Sabogal, C. Restoring Degraded Forest Land with Native Tree Species: The Experience of “Bosques Amazónicos” in Ucayali, Peru. Forests 2019, 10, 851. [Google Scholar] [CrossRef]
- Aguirre, N. Silvicultural Contributions to the Reforestation with Native Species in the Tropical Mountain Rainforest Region of South Ecuador; Technischen Universität München: Munchen, Germany, 2007. [Google Scholar]
- Avendaño-Yáñez, M.; Sánchez-Velásquez, L.; Meave, J.A.; Pineda-López, M. Can Pinus Plantations Facilitate Reintroduction of Endangered Cloud Forest Species? Landsc. Ecol. Eng. 2015, 12, 99–104. [Google Scholar] [CrossRef]
- Bremer, L.L.; Farley, K.A.; Chadwick, O.A.; Harden, C.P. Changes in Carbon Storage with Land Management Promoted by Payment for Ecosystem Services. Environ. Conserv. 2016, 43, 397–406. [Google Scholar] [CrossRef]
- Cargua, F.E.; Rodríguez, M.V.; Recalde, C.G.; Vinueza, L.M. Cuantificación del Contenido de Carbono En una Plantación de Pino Insigne (Pinus radiata) y En Estrato de Páramo de Ozogoche Bajo, Parque Nacional Sangay, Ecuador. Inf. Tecnol. 2014, 25, 83–92. [Google Scholar] [CrossRef]
- Farley, K.A.; Bremer, L.L.; Harden, C.P.; Hartsig, J. Changes in Carbon Storage under Alternative Land Uses in Biodiverse Andean Grasslands: Implications for Payment for Ecosystem Services. Consservation Lett. 2013, 6, 21–27. [Google Scholar] [CrossRef]
- Feyera, S.; Beck, E.; Lüttge, U. Exotic Trees as Nurse-Trees for the Regeneration of Natural Tropical Forests. Trees Struct. Funct. 2002, 16, 245–249. [Google Scholar] [CrossRef]
- Hofstede, R.G.; Groenendijk, J.P.; Coppus, R.; Fehse, J.C.; Sevink, J. Impact of Pine Plantations on Soils and Vegetation in the Ecuadorian High Andes. Mt. Res. Dev. 2002, 22, 159–167. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Fernández-Palacios, J.M. From Pine Plantations to Natural Stands. Ecological Restoration of a Pinus Canariensis Sweet, Ex Spreng Forest. Plant Ecol. 2005, 181, 217–226. [Google Scholar] [CrossRef]
- Luteyn, J.L.; Churchill, S.P. Paramos: A Checklist of Plant Diversity, Geographical Distribution and Botanical Literature; Luteyn, J.L., Ed.; New York Botanical Garden: New York, NY, USA, 1999; ISBN 0893274275. [Google Scholar]
- Ramsay, P.M. The Páramo Vegetation of Ecuador: The Community Ecology, Dynamics and Productivity of Tropical Grasslands in the Andes; University of Wales, Bangor: Bangor, Wales, 1992. [Google Scholar]
- Hofstede, R.G.; Calles, J.; López, V.; Polanco, R.; Torres, F.; Ulloa, J.; Vásquez, A.; Cerra, M. Los Páramos Andinos ¿Qué Sabemos? Estado de Conocimiento Sobre El Impacto del Cambio Climático En El Ecosistema Páramo, 1st ed.; International Union for Conservation of Nature: Quito, Ecuador, 2014. [Google Scholar]
- Vásconez, P.M.; Hofstede, R.G. Los Páramos Ecuatorianos. In Botánica Económica de los Andes Centrales; Moraes, R.M., Øllgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andrés: La Paz, Bolivia, 2006; pp. 91–109. ISBN 9995401215. [Google Scholar]
- Albán, M. Mecanismos Relacionados Con Servicios Ambientales Como una Herramienta para Conservación de los Páramos. In Páramo. Paisaje Estudiado, Habitado, Manejado e Institucionalizado; Mena Vásconez, P., Campaña, J., Castillo, A., Flores, S., Hofstede, R.G., Josse, C., Lasso, S., Medina, G., Ochoa, N., Ortiz, D., Eds.; Abya-Yala: Quito, Ecuador, 2011; pp. 231–246. [Google Scholar]
- Quiroz-Dahik, C.; Crespo, P.; Stimm, B.; Murtinho, F.; Weber, M.; Hildebrandt, P. Contrasting Stakeholders’ Perceptions of Pine Plantations in the Páramo Ecosystem of Ecuador. Sustainability 2018, 10, 1707. [Google Scholar] [CrossRef]
- León-Yánez, S.; Valencia, R.; Pitman, N.; Endara, L.; Ulloa-Ulloa, C.; Navarrete, H. Libro Rojo de las Plantas Endémicas del Ecuador, Segunda Edición, 2nd ed.; Publicaciones del Herbario QCA; León-Yánez, S., Valencia, R., Pitman, N., Endara, L., Ulloa Ulloa, C., Navarrete, H., Eds.; Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2012; ISBN 9789942033932. [Google Scholar]
- Froude, V. Wilding Conifers in New Zealand: Status Report; Pacific Eco-Logic Ltd.: Northland, New Zealand, 2011. [Google Scholar]
- Williams, C.M.; Wardle, M.G. Pinus radiata Invasion in New South Wales: The Extent of Spread. Plant Prot. Q. 2009, 24, 146–156. [Google Scholar]
- Palmberg-Lerche, C.; Ball, J.B. Present Status of Forest Plantations in Latin America and the Caribbean and Review of Related Activities in Tree Improvement. Proceedings of El Manejo Sustentable de los Recursos Forestales, Desafío del Siglo XXI, Valdivia, Chile, 22 November 1998. [Google Scholar]
- Vásquez, D.L.A.; Balslev, H.; Sklenář, P. Human Impact on Tropical-Alpine Plant Diversity in the Northern Andes. Biodivers. Conserv. 2015, 24, 2673–2683. [Google Scholar] [CrossRef]
- Verweij, P.A.; Budde, P.E. Burning and Grazing Gradients in Páramo Vegetation: Initial Ordination Analyses. In Páramo. An Andean Ecoystem under Human Influence; Balslev, H., Luteyn, J.L., Eds.; Academic Press Limited: London, UK, 1992; pp. 177–195. [Google Scholar]
- Cavelier, J.; Santos, C. Efectos de Plantaciones Abandonadas de Especies Exóticas y Nativas Sobre la Regeneración Natural de Un Bosque Montano En Colombia. Rev. De Biol. Trop. 1999, 47, 775–784. [Google Scholar] [CrossRef]
- Van Wesenbeeck, B.K.; Van Mourik, T.; Duivenvoorden, J.F.; Cleef, A.M. Strong Effects of a Plantation with Pinus patula on Andean Subpáramo Vegetation: A Case Study from Colombia. Biol. Conserv. 2003, 114, 207–218. [Google Scholar] [CrossRef]
- Bremer, L.L. Land-Use Change, Ecosystem Services, and Local Livelihoods: Ecological and Socioeconomic Outcomes of Payment for Ecosystem Services in Ecuadorian Páramo Grasslands. Ph.D Thesis, San Diego State University, University of California, Santa Barbara, CA, USA, 2012. [Google Scholar]
- Gloser, V.; Gloser, J. Acclimation Capability of Calamagrostis epigejos and C. arundinacea to Changes in Radiation Environment. Photosynthetica 1996, 32, 203–212. [Google Scholar]
- Quiroz-Dahik, C.; Marín, F.; Arias, R.; Crespo, P.; Weber, M.; Palomeque, X. Comparison of Natural Regeneration in Natural Grassland and Pine Plantations across an Elevational Gradient in the Páramo Ecosystem of Southern Ecuador. Forests 2019, 10, 745. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Ecroyd, C.E.; Leckie, A.C.; Kimberley, M.O. Diversity and Succession of Adventive and Indigenous Vascular Understorey Plants in Pinus radiata Plantation Forests in New Zealand. For. Ecol. Manag. 2003, 185, 307–326. [Google Scholar] [CrossRef]
- Hall, J.M.; Daniels, A.E.; Lambin, E.F.; Van Holt, T.; Balthazar, V. Trade-Offs between Tree Cover, Carbon Storage and Floristic Biodiversity in Reforesting Landscapes. Landsc. Ecol. Eng. 2012, 27, 1135–1147. [Google Scholar] [CrossRef]
- Sarmiento, L.; Llambí, L.D.; Escalona, A.; Marquez, N. Vegetation Patterns, Regeneration Rates and Divergence in an Old-Field Succession of the High Tropical Andes. Plant Ecol. 2003, 166, 63–74. [Google Scholar] [CrossRef]
- Van der Hammen, T.; Cleef, A. Development of the High Andean Paramo Flora and Vegetation. In High Altitude Tropical Biogeography; Vuilleumier, F., Monasterio, M., Eds.; Oxford University Press: New York, NY, USA, 1986; pp. 153–201. [Google Scholar]
- Secretaria General de la Comunidad Andina; Ministerio del Ambiente del Ecuador; Proyecto de Adaptación al Impacto del Retroceso Acelerado de Glaciares en los Andes Tropicales; Fondo para la Protección del Agua. Diseño de la Red de Monitoreo Hidrológico en las Microcuencas de los Ríos Pita, San Pedro, Papallacta y Antisana; Ministerio del Ambiente del Ecuador: Quito, Ecuador, 2011; pp. 60–104. [Google Scholar]
- FONAG; EPMAPS. Anuario Hidrometeorológico 2019; Fondo para la Protección del Agua: Quito, Ecuador, 2019; Volume 13. [Google Scholar]
- FONAG; EPMAPS. Anuario Hidrometeorológico 2020; Fondo para la Protección del Agua: Quito, Ecuador, 2020; Volume 14. [Google Scholar]
- INAMHI. Anuario Meteorológico; Instituto Nacional de Meteorología e Hidrología del Ecuador: Quito, Ecuador, 2013. [Google Scholar]
- Aguirre, N.; Torres, J.; Velasco-Linares, P. Guía para la Restauración Ecológica En los Páramos del Antisana; FONAG, Ed.; Fondo para la Protección del Agua-FONAG: Quito, Ecuador, 2014; ISBN 9942-9983-3. [Google Scholar]
- Sklenár, P.; Ramsay, P.M. Diversity of Zonal Páramo Plant Communities in Ecuador. Divers. Distrib. 2001, 7, 113–124. [Google Scholar] [CrossRef]
- Beltrán, K.; Salgado, S.; Cuesta, F.; León-Yánez, S.; Romoleroux, K.; Ortiz, E.; Cárdenas, A.; Velástegui, A. Distribución Espacial Sistemas Ecológicos y Caracterización Florística de los Páramos En El Ecuador; EcoCiencia, Proyecto Páramo Andino y Herbario QCA, Ed.; EcoCiencia: Quito, Ecuador, 2009. [Google Scholar]
- Lemmon, P.E. A New Instrument for Measuring Forest Overstory Density. J. For. 1957, 55, 667–668. [Google Scholar]
- Ramsay, P.M.; Oxley, E.R.B. The Growth Form Composition of Plant Communities in the Ecuadorian Páramos. Plant Ecol. 1997, 131, 173–192. [Google Scholar] [CrossRef]
- Romoleroux, K.; Cárate Tandalla, D.; Erler, R.; Navarrete, H. Plantas Vasculares de los Bosques de Polylepis En los Páramos de Oyacachi; Centro de Publicaciones de la Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2016; ISBN 9978772731. [Google Scholar]
- MECN-INB. Plantas de los Páramos del Distrito Metropolitano de Quito, Ecuador; Ulloa-Ulloa, C., Fernández, D., Eds.; Publicacio.: Quito, Ecuador, 2015; ISBN 9789942998842. [Google Scholar]
- Aguilar, Z.; Hidalgo, P.; Ulloa Ulloa, C. Plantas Útiles de los Páramos de Zuleta, Ecuador; Proyecto d.; PPA-Ecociencia: Quito, Ecuador, 2009; ISBN 9789978994023. [Google Scholar]
- Chimbolema, S.; Suárez-Duque, D.; Peñafiel Cevallos, M.; Acurio, C.; Paredes, T. Guía de Plantas de la Reserva Ecológica El Ángel; Peñafiel Cevallos, M., Ed.; Corporación Grupo Randi Randi: Quito, Ecuador, 2014; ISBN 978-9942-13-550-6. [Google Scholar]
- Van der Maarel, E. Relations between Sociological-Ecological Species Groups and Ellenberg Indicator Values. Phytocoenologia 1993, 23, 343–362. [Google Scholar] [CrossRef]
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Gloser, V.; Scheurwater, I.; Lambers, H. The Interactive Effect of Irradiance and Source of Nitrogen on Growth and Root Respiration of Calamagrostis Epigejos. New Phytol. 1996, 134, 407–412. [Google Scholar] [CrossRef]
- Bader, M.Y.; Van Geloof, I.; Rietkerk, M. High Solar Radiation Hinders Tree Regeneration above the Alpine Treeline in Northern Ecuador. Plant Ecol. 2007, 191, 33–45. [Google Scholar] [CrossRef]
- Jørgensen, P.M.; León-Yánez, S. (Eds.) Catalogue of the Vascular Plants of Ecuador; Missouri Botanical Garden Press: St Louis, MO, USA, 1999. [Google Scholar]
- Ministerio del Ambiente del Ecuador. Sistema de Clasificación de Ecosistemas del Ecuador Continental, 1st ed; Galeas, R., Guevara, J.E., Medina-Torres, B., Chinchero, M.Á., Herrera, X., Eds.; Subsecretaria de Patrimonio Natural: Quito, Ecuador, 2013. [Google Scholar]
- Subsecretaría de Patrimonio Natural; Acosta-Buenaño, N.; Beltrán, L.; Chinchero, M.Á.; Galeas, R.; Herrera, X.; Iglesias, J.; Medina-Torres, B.; Morales, C.; Rivas-Torres, G.; et al. Información florística del Proyecto Mapa de Vegetación para la Clasificación de Ecosistemas del Ecuador Continental. Available online: https://www.gbif.org/dataset/9bbdb0ea-b792-4047-be06-c4285ad47f3f (accessed on 14 December 2020).
- Bremer, L.L.; Farley, K.A. Does Plantation Forestry Restore Biodiversity or Create Green Deserts? A Synthesis of the Effects of Land-Use Transitions on Plant Species Richness. Biodivers. Conserv. 2010, 19, 3893–3915. [Google Scholar] [CrossRef]
- Mena Vasconez, P.; Medina, G.; Hofstede, R.G.M. los Páramos del Ecuador. Particularidades, Problemas y Perspectivas; Mena Vásconez, P., Medina, G., Hofstede, R.G., Eds.; Abya Yala/Proyecto Páramo: Quito, Ecuador, 2001; ISBN 9978047271. [Google Scholar]
- Lemenih, M.; Gidyelew, T.; Teketay, D. Effects of Canopy Cover and Understory Environment of Tree Plantations on Richness, Density and Size of Colonizing Woody Species in Southern Ethiopia. For. Ecol. Manag. 2004, 194, 1–10. [Google Scholar] [CrossRef]
- Corredor-Velandia, S.; Vargas Ríos, O. Efectos de la Creación de Claros Experimentales Con Diferentes Densidades, Sobre los Patrones Iniciales de Sucesión Vegetal En Plantaciones de Pinus patula. In Restauración Ecológica del Bosque Altoandino. Estudios Diagnósticos y Experimentales en los Alrededores del Embalse de Chisacá; Universidad Nacional de Colombia: Bogotá, Colombia, 2007; pp. 336–352. [Google Scholar]
- Matson, E.C.; Bart, D.J. Plant—Community Responses to Shrub Cover in a Páramo Grassland Released from Grazing and Burning. Austral Ecol. 2014, 39, 918–928. [Google Scholar] [CrossRef]
- Grubb, P.J.; Lloyd, J.R.; Pennington, T.D.; Páez-Bimos, S. A Historical Baseline Study of the Páramo of Antisana in the Ecuadorian Andes Including the Impacts of Burning, Grazing and Trampling. Plant Ecol. Divers. 2020, 13, 225–256. [Google Scholar] [CrossRef]
- Podwojewski, P.; Poulenard, J.; Zambrana, T.; Hofstede, R.G. Overgrazing Effects on Vegetation Cover and Properties of Volcanic Ash Soil in the Páramo of Llangahua and la Esperanza (Tungurahua, Ecuador). Soil Use Manag. 2002, 18, 45–55. [Google Scholar] [CrossRef]
- Hofstede, R.G.; Rossenaar, A.J. Biomass of Grazed, Burned, and Undisturbed Paramo Grasslands, Colombia. II. Root Mass and Aboveground: Belowground Ratio. Arct. Alp. Res. 1995, 27, 13–18. [Google Scholar] [CrossRef]



| Environmental Variables | Antisana | Cajas | Tisaleo | Cotopaxi |
|---|---|---|---|---|
| Mean annual temperature (°C) [50,51,52,53] | 8.3 | 10 * | 10 * | 11 * |
| Mean annual rainfall (mm) [52,53] | 900 | 1200 * | 900 * | 1110 * |
| Mean altitude in grasslands (m a.s.l.) | 3739 | 3734 | 3858 | 3695 |
| Mean altitude in pine plantations (m a.s.l.) | 3556 | 3699 | 3874 | 3740 |
| Age of pine plantation | 8 | 16 | 41 | 53 |
| Pine tree cover (%) | 75 | 86 | 66 | 47 |
| Basal area of pine trees (m2/ha) | 17 | 10.5 | 26 | 29.5 |
| Mean height of pine trees (m) | 8 | 15 | 15 | 18 |
| pH H2O under grassland and pine plantation (10–30 cm depth) + | 5.8–6.2 | 6.2–6.4 | 6.1–5.7 | 6.2–5.6 |
| Mean slope of grasslands and pine plantation plots + | 15.8–19.1 | 26.1–30.1 | 14.4–14.7 | 27.7–27.6 |
| Slope of grasslands and pine plantation plots | S–SW | NE–SE | SE–SE | N–N |
| Antisana | Cajas | Tisaleo | Cotopaxi | Contribution to R2 | Change under Pine (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Grassland | Pine Plantation | Grassland | Pine Plantation | Grassland | Pine Plantation | Grassland | Pine Plantation | Zone | Land Use | Z × LU | ||
| Location WGS 84 | 0°00′13″ S 78°05′11″ W | 0°07′1.2 S 78°05′36″ W | 2°01′00″ S 79°03′0.4″ W | 2°01′00″ S 79°03′36″ W | 1°06′00″ S 78°01′31″ W | 1°06′53″ S 78°11′0.1″ W | 0°01′11″ S 78°08′36″ W | 0°10′12″ S 78°08′00″ W | ||||
| Altitude (m a.s.l.) | 3740 | 3560 | 3735 | 3698 | 3858 | 3871 | 3695 | 3742 | ||||
| Herbaceous height (cm) | 80 | 15 | 60 | 20 | 60 | 25 | 100 | 60 | ||||
| Shrub height (cm) | 50 | 20 | 140 | 40 | 100 | 50 | 120 | 40 | ||||
| Plant cover (%) | ||||||||||||
| Total cover | 101.5 ± 3.3 | 66.3 ± 39.72 | 94.0 ± 4.7 | 3.4 ± 3.4 *** | 85.0 ± 23.5 | 55.5 ± 21.3 ** | 92.2 ± 19.1 | 57.4 ± 7.3 *** | 0.14 | 0.46 | 0.13 | −47.5 *** |
| Moss cover | 2.5 ± 1.2 | 1.7 ± 2.1 | 2.5 ± 1.7 | 10.5 ± 9.4 | 23.6 ± 14 | 9.9 ± 5.4 | 17.3 ± 15.2 | 8.6 ± 10.6 ** | 0.26 | 0.03 | 0.13 | −3.8 |
| Woody species | 3.9 ± 2.1 | 4.3 ± 6.5 | 2.0 ± 1.7 | 1.1 ± 1.0 | 20.2 ± 12.0 | 9.9 ± 9.3 * | 30.8 ± 9.7 | 12.0 ± 3.7 ** | 0.48 | 0.10 | 0.11 | −7.4 * |
| Herbaceous | 97.6 ± 2.6 | 62.0 ± 38.9 * | 92.0 ± 5.0 | 2.3 ± 3.0 *** | 64.8 ± 15.8 | 45.6 ± 15.4 * | 61.4 ± 17.3 | 45.4 ± 5.9 | 0.15 | 0.39 | 0.21 | −40.1 *** |
| Tussocks | 86.3 ± 5.4 | 13.1 ± 28.6 *** | 83.1 ± 5.7 | 0.1 ± 0.3 *** | 41.3 ± 13.7 | 0.5 ± 0.9 *** | 39.9 ± 13 | 33.6 ± 5.8 | 0.10 | 0.57 | 0.20 | −50.8 *** |
| Cushions | 2.2 ± 1.0 | 0.1 ± 0.3 * | 1.6 ± 1.5 | 0.03 ± 0.05 | 8.6 ± 5.7 | 0.4 ± 1.1 *** | 1.4 ± 1.6 | 0.0 | 0.21 | 0.24 | 0.17 | −3.4 *** |
| Rosettes | 0.5 ± 0.3 | 0.04 ± 0.1 * | 4.3 ± 2.2 | 0.9 ± 1.2 *** | 0.8 ± 0.8 | 0.7 ± 0.4 | 11.3 ± 6.3 | 0.9 ± 1.2 *** | 0.29 | 0.18 | 0.24 | −3.6 *** |
| Small herbs | 8.6 ± 3.9 | 48.8 ± 42.8 | 3.0 ± 1.1 | 1.2 ± 2.0 | 14.1 ± 11.2 | 44.1 ± 14.9 *** | 8.8 ± 3.8 | 10.9 ± 4.5 | 0.26 | 0.14 | 0.15 | 17.6 ** |
| Light-demanding species cover | 89.7 ± 5.0 | 13.3 ± 28.5 *** | 91.2 ± 5.3 | 1.1 ± 1.2 *** | 51.9 ± 14.9 | 2.0 ± 1.9 *** | 54 ± 18.2 | 34.5 ± 5.1 ** | 0.07 | 0.67 | 0.14 | −59.0 *** |
| Shade-tolerant species cover | 11.8 ± 5.5 | 53.2 ± 44.4 * | 2.8 ± 1.7 | 2.4 ± 2.4 | 33.2 ± 14.3 | 56.1 ± 20.8 | 38.2 ± 12.1 | 23.8 ± 7.2 | 0.34 | 0.05 | 0.16 | 12.4 |
| Species richness | ||||||||||||
| Total | 21.8 ± 1.6 | 13.8 ± 5.4 ** | 29.7 ± 4.9 | 13.9 ± 5.7 *** | 22.2 ± 6.3 | 13.9 ± 3.9 *** | 21.9 ± 2.8 | 23.1 ± 2.8 | 0.10 | 0.32 | 0.20 | −7.7 *** |
| Herbaceous | 17.7 ± 1.5 | 9.7 ± 3.5 *** | 23.4 ± 5.8 | 8.3 ± 4.5 *** | 17.2 ± 5.5 | 10.6 ± 3 ** | 13.4 ± 2.3 | 13 ± 2.8 | 0.03 | 0.40 | 0.19 | −7.5 *** |
| Woody species | 4.1 ± 0.6 | 4.1 ± 2.2 | 6.2 ± 2.3 | 5.6 ± 1.5 | 5.0 ± 1.0 | 3.3 ± 1.3 | 8.4 ± 1.5 | 10.0 ± 1.8 | 0.62 | 0.00 | 0.05 | −0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Torres, B.; Jonard, M.; Rendón, M.; Jacquemart, A.-L. Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation. Forests 2022, 13, 1499. https://doi.org/10.3390/f13091499
Medina-Torres B, Jonard M, Rendón M, Jacquemart A-L. Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation. Forests. 2022; 13(9):1499. https://doi.org/10.3390/f13091499
Chicago/Turabian StyleMedina-Torres, Byron, Mathieu Jonard, Melina Rendón, and Anne-Laure Jacquemart. 2022. "Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation" Forests 13, no. 9: 1499. https://doi.org/10.3390/f13091499
APA StyleMedina-Torres, B., Jonard, M., Rendón, M., & Jacquemart, A.-L. (2022). Effects of Pine Plantation on Native Ecuadorian Páramo Vegetation. Forests, 13(9), 1499. https://doi.org/10.3390/f13091499