
Phytochemical Composition of Extractives in the Inner Cork Layer of Cork Oaks with Low and Moderate Coraebus undatus Attack

 ,
,  ,
, 
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling
2.2. Lipophilic Compounds Extraction
2.3. Hydrophilic Compounds Extraction
2.4. Chemical Characterization of the Lipophilic Extract
2.5. Ethanol-Water Extract Composition
2.6. Antioxidant Activity of Ethanol–Water Extracts
2.7. Statistical Analysis
3. Results and Discussion
3.1. Incidence of C. undatus
3.2. Extractable Components in the “Inner Cork”
3.3. Lipophilic Extractives Composition
3.4. Composition of Hydrophilic Extractives
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Bugalho, M.N.; Pinto-Correia, T.; Pulido, F. Human use of natural capital generates cultural and other ecosystem services in montado and dehesa oak woodlands. Reconnecting Nat. Cult. Cap. 2018, 3, 115–123. [Google Scholar]
- Bugalho, M.N.; Caldeira, M.C.; Pereira, J.S.; Aronson, J.; Pausas, J.G. Mediterranean cork oak savannas require human use to sustain biodiversity and ecosystem services. Front. Ecol. Environ. 2011, 9, 278–286. [Google Scholar] [CrossRef]
- Pereira, H. Cork: Biology, Production and Uses; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- de Castro, A.; Avillez, F.; Rodrigues, V.; da Silva, F.G.; Santos, F.; Rebelo, F.; Jorge, M.N.; Aires, N. The Cork Sector: From the Forest to the Consumer; APCOR, Associação Portuguesa da Cortiça: Santa Maria de Lamas, Portugal, 2020. [Google Scholar]
- Jiménez, A.; Gallardo, A.; Antonietty, C.A.; Villagrán, M.; Ocete, M.E.; Soria, F.J. Distribution of Coraebus undatus (Coleoptera: Buprestidae) in cork oak forests of southern Spain. Int. J. Pest Manag. 2012, 58, 281–288. [Google Scholar] [CrossRef]
- Sallé, A.; Nageleisen, L.-M.; Lieutier, F. Bark and wood boring insects involved in oak declines in Europe: Current knowledge and future prospects in a context of climate change. For. Ecol. Manag. 2014, 328, 79–93. [Google Scholar] [CrossRef]
- Martín, C. Notas sobre Coraebus undatus Fabr y Mars, plaga del alcornoque. Bol. Serv. Plagas For. 1964, 7, 95–101. [Google Scholar]
- Gallardo, A.; Jiménez, A.; Antonietty, C.A.; Villagrán, M.; Ocete, M.E.; Soria, F.J. Forecasting infestation by Coraebus undatus (Coleoptera, Buprestidae) in cork oak forests. Int. J. Pest Manag. 2012, 58, 275–280. [Google Scholar] [CrossRef]
- Cárdenas, A.M.; Toledo, D.; Gallardo, P.; Guerrero-Casado, J. Assessment and distribution of damages caused by the trunk-boring insects Coraebus undatus (Fabricius) (Coleoptera: Buprestidae) and Reticulitermes grassei Clément (Blattodea: Rhinotermitidae) in Mediterranean restored cork-oak forests. Forests 2021, 12, 1501. [Google Scholar] [CrossRef]
- Gonzalez-Adrados, J.R.; Pereira, H. Classification of defects in cork planks using image analysis. Wood Sci. Technol. 1996, 30, 207–215. [Google Scholar] [CrossRef]
- Oliveira, V.; Lopes, P.; Cabral, M.; Pereira, H. Influence of cork defects in the oxygen ingress through wine stoppers: Insights with X-ray tomography. J. Food Eng. 2015, 165, 66–73. [Google Scholar] [CrossRef]
- Lauw, A.; Oliveira, V.; Lopes, F.; Pereira, H. Variation of cork quality for wine stoppers across the production regions in Portugal. Eur. J. Wood Prod. 2018, 76, 123–132. [Google Scholar] [CrossRef]
- Pereira, H. Variability of the chemical composition of cork. Bioresources 2013, 8, 2246–2256. [Google Scholar] [CrossRef]
- Pereira, H. Chemical composition and variability of cork from Quercus suber L. Wood Sci. Technol. 1988, 22, 211–218. [Google Scholar] [CrossRef]
- Piasecka, A.; Jedrzejczak-Rey, N.; Bednarek, P. Secondary metabolites in plant innate immunity: Conserved function of divergent chemicals. New Phytol. 2015, 206, 948–964. [Google Scholar] [CrossRef]
- López-Goldar, X.; Villari, C.; Bonello, P.; Borg-Karlson, A.K.; Grivet, D.; Zas, R.; Sampedro, L. Inducibility of plant secondary metabolites in the stem predicts genetic variation in resistance against a key insect herbivore in Maritime pine. Front. Plant Sci. 2018, 9, 1651. [Google Scholar] [CrossRef]
- Chandramu, C.; Manohar, R.D.; Krupadanam, D.G.L.; Dashavantha, R. Isolation, characterization and biological activity of betulinic acid and ursolic acid from Vitex negundo L. Phytother. Res. 2003, 17, 129–134. [Google Scholar] [CrossRef]
- Baskar, K.; Duraipandiyan, V.; Ignacimuthu, S. Bioefficacy of the triterpenoid friedelin against Helicoverpa armigera (Hub.) and Spodoptera litura (Fab.) (Lepidoptera: Noctuidae). Pest Manag. Sci. 2014, 70, 1877–1883. [Google Scholar] [CrossRef]
- Punia, A.; Chauhan, N.S.; Kaur, S.; Sohal, S.K. Effect of ellagic acid on the larvae of Spodoptera litura (Lepidoptera: Noctuidae) and its parasitoid Bracon hebetor (Hymenoptera: Braconidae). J. Asia Pac. Entomol. 2020, 23, 660–665. [Google Scholar] [CrossRef]
- Lindroth, R.L.; Peterson, S.S. Effects of plant phenols of performance of Southern armyworm larvae. Oecologia 1988, 75, 185–189. [Google Scholar] [CrossRef]
- Fernandes, A.; Fernandes, I.; Cruz, L.; Mateus, N.; Cabral, M.; Freitas, V. Antioxidant and biological properties of bioactive phenolic compounds from Quercus suber L. J. Agric. Food Chem. 2009, 57, 11154–11160. [Google Scholar] [CrossRef]
- Fernandes, A.; Sousa, A.; Mateus, N.; Cabral, M.; Freitas, V. Analysis of phenolic compounds in cork from Quercus suber L. by HPLC–DAD/ESI–MS. Food Chem. 2011, 125, 1398–1405. [Google Scholar] [CrossRef]
- Castola, V.; Marongiu, B.; Bighelli, A.; Floris, C.; Lai, A.; Casanova, J. Extractives of cork (Quercus suber L.): Chemical composition of dichloromethane and supercritical CO2 extracts. Ind. Crops Prod. 2005, 21, 65–69. [Google Scholar] [CrossRef]
- Ferreira, F.; Santos, S.; Pereira, H. In vitro screening for acetylcholinesterase inhibition and antioxidant activity of Quercus suber cork and cork back extracts. Evid. Based Complement. Alternat. Med. 2020, 2020, 3825629. [Google Scholar] [CrossRef]
- Şen, A.; Zhianski, M.; Glushkova, M.; Petkova, K.; Ferreira, J.; Pereira, H. Chemical composition and cellular structure of corks from Quercus suber trees planted in Bulgaria and Turkey. Wood Sci. Technol. 2016, 50, 1261–1276. [Google Scholar] [CrossRef]
- Touati, R.; Santos, S.A.; Rocha, S.M.; Belhamel, K.; Silvestre, A.J. The potential of cork from Quercus suber L. grown in Algeria as a source of bioactive lipophilic and phenolic compounds. Ind. Crops Prod. 2015, 76, 936–945. [Google Scholar] [CrossRef]
- Conde, E.; Cadahía, E.; García-Vallejo, M.C.; Fernández de Simóm, B. Polyphenolic composition of Quercus suber cork from different Spanish provenances. J. Agric. Food Chem. 1998, 46, 3166–3171. [Google Scholar] [CrossRef]
- Conde, E.; García-Vallejo, M.; Pereira, C.; Cadahía, E. Waxes composition of reproduction cork from Quercus suber and its variability throughout the industrial processing. J. Wood Sci. Technol. 1999, 33, 229–244. [Google Scholar] [CrossRef]
- Du Merle, P.D.; Attie, M. Coroebus undatus (Coleoptera Buprestidae) sur chêne-liége dans le sud-est de la France: Estimation des dégâts, relations entre ceux-ci et certains facteurs du milieu. Ann Sci. For. 1992, 49, 571–588. [Google Scholar] [CrossRef]
- Eglinton, G.; Hunneman, D. Gas chromatographic-mass spectrometric studies of long-chain hydroxy acids-I. Phytochemistry 1968, 7, 313–322. [Google Scholar] [CrossRef]
- Kolattukudy, P.; Agrawal, V. Structure and composition of aliphatic constituents of potato tuber skin. Lipids 1974, 9, 682–691. [Google Scholar] [CrossRef]
- Marques, A.V.; Pereira, H. A methodological approach for the simultaneous quantification of glycerol and fatty acids from cork suberin in a single GC run. Phytochem. Anal. 2019, 30, 687–699. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Chang, C.; Yang, M.; Wen, H.; Chern, J. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Abdalla, S.; Pizzi, A.; Ayed, N.; Bouthoury, F.C.; Charrier, B.; Bahabri, F.; Ganash, A. MALDI-TOF analysis of Aleppo pine (Pinus halepensis) bark tannin. BioResource 2014, 9, 3396–3406. [Google Scholar] [CrossRef]
- Mirkin, B. Nonconvex Optimization and its Applications. In Mathematical Classification and Clustering; Springer Science & Business Media: New York, NY, USA, 1996. [Google Scholar]
- Rencher, A.C. Methods of Multivariate Analysis; John Wiley and Sons: Hoboken, NJ, USA, 1995. [Google Scholar]
- Branco, M.; Bragança, H.; Sousa, E.; Phillips, A.J. Pests and diseases in Portuguese forestry: Current and new threats. In Forest context and policies in Portugal; Reboredo, F., Ed.; Springer: Cham, Switzerland, 2014; pp. 117–154. [Google Scholar]
- Cunha, M.; Lourenço, A.; Barreiros, S.; Paiva, A.; Simões, P. Valorization of cork using subcritical water. Molecules 2020, 25, 4695. [Google Scholar] [CrossRef] [PubMed]
- Jové, P.; Olivella, M.À.; Cano, L. Study of the variability in chemical composition of bark layers of Quercus suber L. from different production areas. BioResources 2011, 6, 1806–1815. [Google Scholar]
- González-Coloma, A.; López-Balboa, C.; Santana, O.; Reina, M.; Fraga, B.M. Triterpene-based plant defenses. Phytochem. Rev. 2011, 10, 245–260. [Google Scholar] [CrossRef]
- Moiteiro, C.; Joao, M.; Curto, O.M.; Mohamed, N.; Bailen, M.; Martínez-Dıaz, R.; González-Coloma, A. Biovalorization of friedelane triterpenes derived from cork processing industry byproducts. J. Agric. Food Chem. 2006, 54, 3566–3571. [Google Scholar] [CrossRef]
- Santos, S.A.O.; Villaverde, J.J.; Sousa, A.F.; Coelho, J.F.J.; Neto, C.P.; Silvestre, A.J.D. Phenolic composition and antioxidant activity of industrial cork by-products. Ind. Crops Prod. 2013, 47, 262–269. [Google Scholar] [CrossRef]
- Santos, S.A.O.; Pinto, P.C.R.O.; Silvestre, A.J.D.; Neto, C.P. Chemical composition and antioxidant activity of phenolic extracts of cork from Quercus suber L. Ind. Crops Prod. 2010, 31, 521–526. [Google Scholar] [CrossRef]
- Witzell, J.; Martin, J.A. Phenolic metabolites in the resistance of northern forest trees to pathogens—past experiences and future prospects. Can. J. For. Res. 2008, 38, 2711–2727. [Google Scholar] [CrossRef]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. Phytochem. Adv. Research. 2006, 661, 23–67. [Google Scholar]
- Pimentel, C.S.; Firmino, P.N.; Calvão, T.; Ayres, M.P.; Miranda, I.; Pereira, H. Pinewood nematode population growth in relation to pinephloem chemical composition. Plant Pathol. 2016, 5, 856–865. [Google Scholar]
- Simões, R.; Pimentel, C.; Ferreira-Dias, S.; Miranda, I.; Pereira, H. Phytochemical characterization of phloem in maritime pine and stone pine in three sites in Portugal. Heliyon 2021, 7, e06718. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
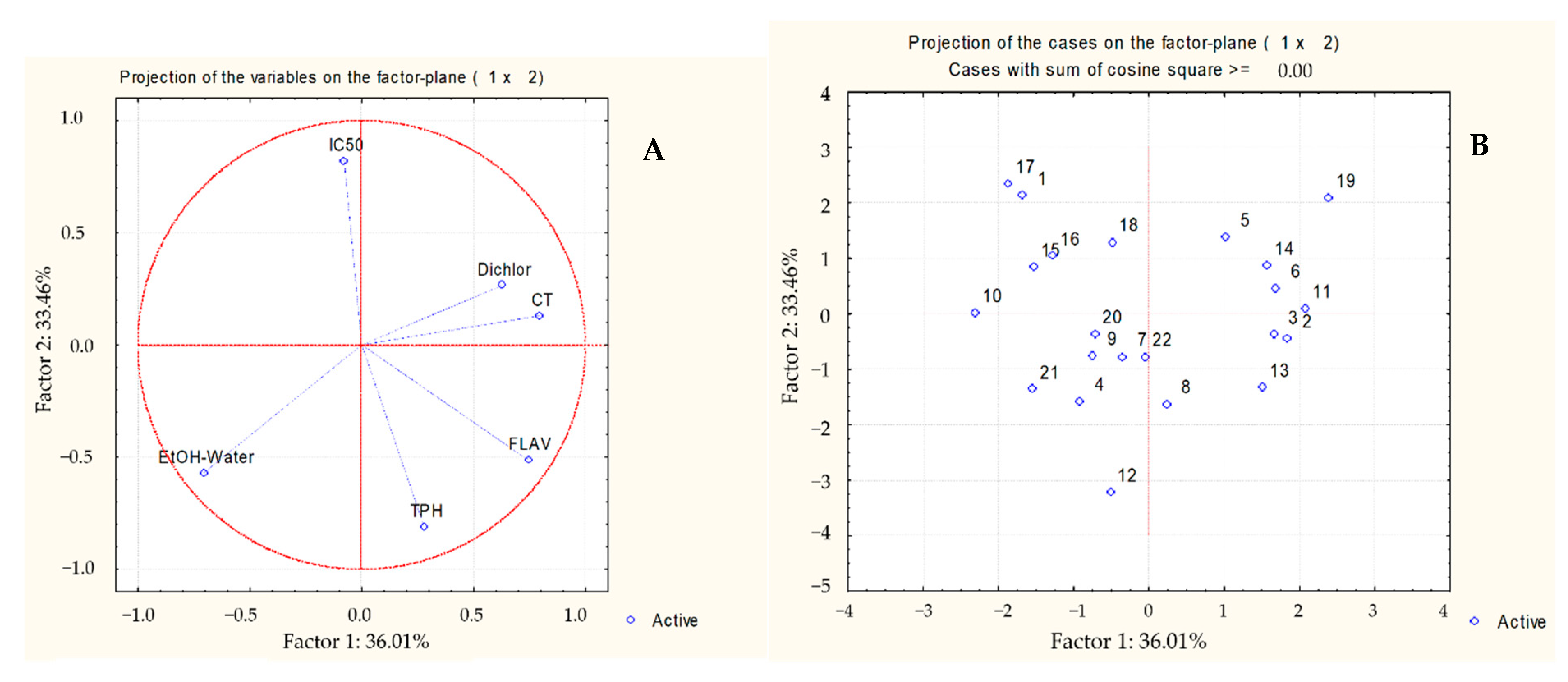{kind=link}
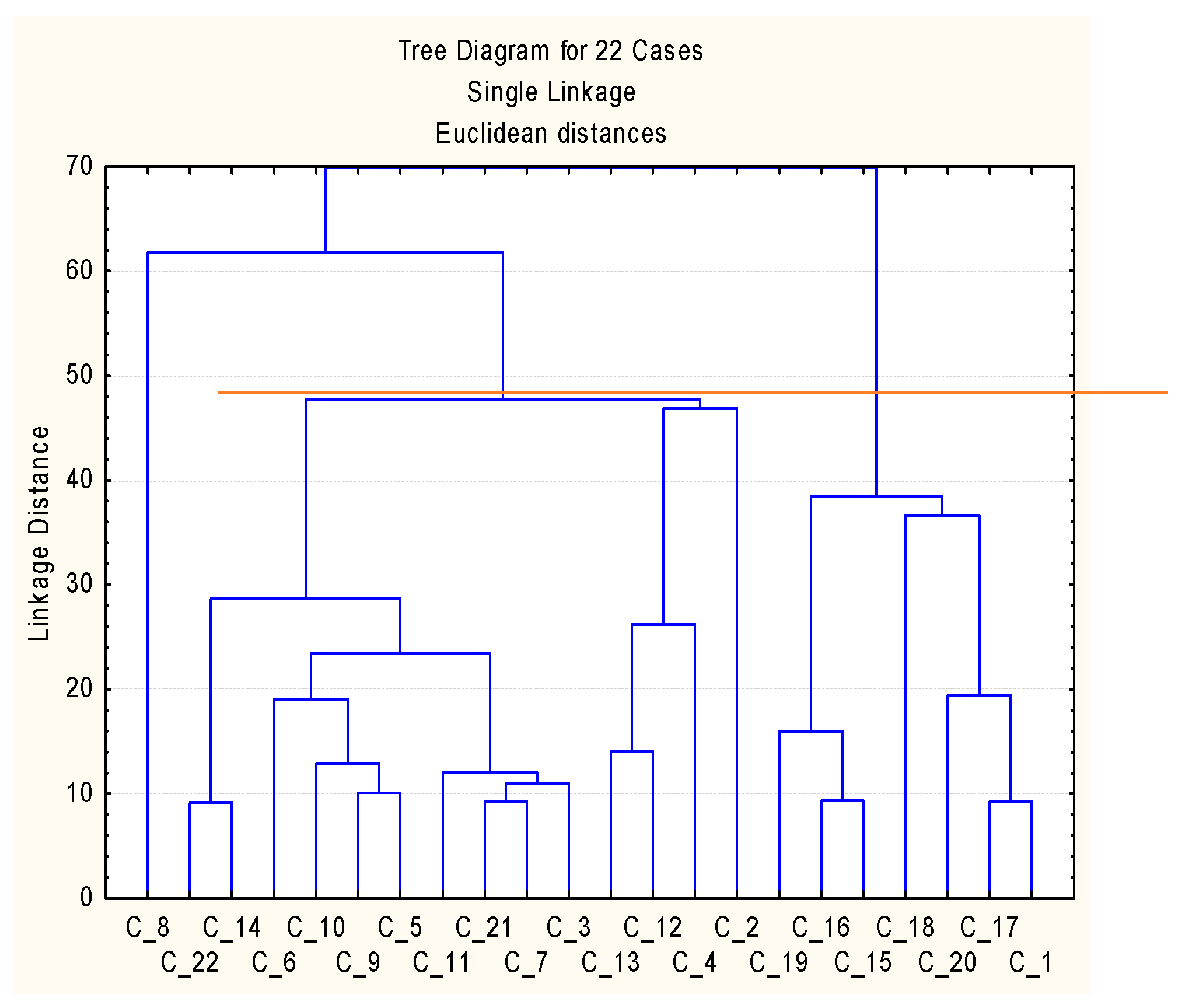{kind=link}
| Low Attack | Moderate Attack | |
|---|---|---|
| Total extractives | 21.6 a ± 2.7 | 22.0 a ± 1.7 |
| Dichloromethane extractives | 2.3 a ± 1.2 | 1.7 a ± 1.5 |
| Ethanol-water extractives | 19.3 a ± 3.3 | 20.3 a ± 2.5 |
| Low Attack | Moderate Attack | |
|---|---|---|
| Dichloromethane Extractives, % Dry Mass | 2.31 a ± 1.2 | 1.65 a ± 1.5 |
| Alkanols | 1.48 a ± 0.14 | 1.91 a ± 0.35 |
| Hexadecan-1-ol | 0.01 ± 0.01 | 0.04 ± 0.03 |
| Octadecan-1-ol | 0.04 ± 0.05 | 0.03 ± 0.05 |
| Eicosan-1-ol | 0.16 ± 0.11 | 0.23 ± 0.14 |
| Docosan1-ol | 0.67 ± 0.39 | 0.97 ± 1.07 |
| Tetracosan-1-ol | 0.53 ± 0.25 | 0.60 ± 0.74 |
| Hexacosan-1-ol | 0.07 ± 0.04 | 0.04 ± 0.06 |
| Alkanoic acids | 6.05 a ± 0.43 | 8.82 b ± 0.49 |
| Saturated fatty acids | 2.80 a ± 0.36 | 4.57 b ± 0.37 |
| Hexadecanoic acid | 1.45 ± 0.67 | 2.62 ± 0.87 |
| Heptadecanoic acid | 0.07 ± 0.03 | 0.08 ± 0.02 |
| Octadecanoic acid | 0.96 ± 0.85 | 1.47 ± 0.73 |
| Eicosanoic acid | 0.14 ± 0.16 | 0.19 ± 0.09 |
| Docosanoic acid | 0.18 ± 0.07 | 0.21 ± 0.05 |
| Unsaturated fatty acids | 3.30 a ± 0.60 | 4.35 a ± 0.64 |
| 9-cis-Hexadecenoic acid | 0.53 ± 0.01 | 0.58 ± 0.25 |
| 9,12-Octadecadienoic acid | 0.52 ± 0.51 | 1.34 ± 1.87 |
| 9,12,15-Octadecatrienoic acid | 0.07 ± 0.09 | 0.37 ± 0.52 |
| 9-Octadecenoic acid | 1.78 ± 2.03 | 1.89 ± 0.51 |
| 13-Octadecenoic acid | 0.35 ± 0.38 | 0.09 ± 0.03 |
| Substituted fatty acids | 0.93 a ± 0.19 | 1.24 a ± 0.37 |
| 2-Hydroxy-2-methylpropanoic acid | 0.76 ± 0.09 | 0.84 ± 0.57 |
| 2-Hydroxy-4-methylpentanoic acid | 0.17 ± 0.29 | 0.39 ± 0.17 |
| Dicarboxylic acids | 0.27 a ± 0.13 | 0.3 a ± 0.20 |
| Saturated dicarboxylic acid | 0.17 a ± 0.20 | 0.21 a ± 0.19 |
| Propanedioic acid | 0.04 ± 0.06 | 0.03 ± 0.05 |
| Nonanedioic acid | 0.13 ± 0.25 | 0.18 ± 0.32 |
| Substituted dicarboxylic acid | 0.10 a ± 0.08 | 0.13 a ± 0.22 |
| 2-Hydroxydecanedioic acid | 0.10 ± 0.08 | 0.13 ± 0.22 |
| Glycerol derivatives | 1.90 a ± 1.17 | 1.20 a ±0.21 |
| Glycerol | 1.52 ± 0.93 | 0.91 ± 0.37 |
| 2,3-Dihydroxypropyl hexadecanoate | 0.27 ± 0.19 | 0.14 ± 0.07 |
| 2,3-Dihydroxypropyl octadecanoate | 0.11 ± 0.03 | 0.15 ± 0.20 |
| Terpenes | 77.95 a ± 3.48 | 76.12 a ± 2.29 |
| Squalene | 0.72 ± 0.95 | 0.62 ± 0.60 |
| Lupeol | 0.65 ± 0.15 | 0.74 ± 0.42 |
| Friedelane-1-ene-3-one | 17.56 ± 9.18 | 16.92 ± 10.06 |
| Erythrodiol | 0.28 ± 0.17 | 0.40 ± 0.34 |
| Friedelin | 37.56 ± 14.41 | 36.59 ± 5.03 |
| Lup-20(29)-en-3-one | 0.24 ± 0.19 | 0.19 ± 0.24 |
| Betulin | 0.97 ± 0.00 | 1.13 ±0.55 |
| Betulinic acid | 15.71 ± 15.3 | 16.43 ± 10.21 |
| Betulinaldehyde | 0.32 ± 0.30 | 0.21 ± 0.15 |
| D:A-Friedooleanan-28-al, 3 oxo (Canophyllal) | 0.63 ± 0.22 | 0.87 ± 0.27 |
| D:A-Friedooleanan-3-one, 28-hydroxy- | 1.06 ± 1.28 | 0.68 ± 0.63 |
| D:A-Friedo-2,3-secooleanane-2,3-dioic acid, dimethyl ester, (4R)- | 2.25 ± 2.53 | 1.33 ± 1.18 |
| Sterols | 3.19 a ± 0.37 | 4.32 a ± 0.22 |
| β-Sitosterol | 1.86 ± 1.00 | 3.05 ± 0.62 |
| Lanosterol | 0.42 ± 0.04 | 0.34 ± 0.09 |
| Cycloartenol | 0.30 ± 0.14 | 0.30 ± 0.25 |
| Cycloeucalenol | 0.39 ± 0.00 | 0.50 ± 0.10 |
| Sitosteryl-3beta-D-Glucopiranoside | 0.22 ± 0.28 | 0.13 ± 0.17 |
| Aromatic compounds | 1.90 a ± 0.17 | 1.54 a ± 0.13 |
| Benzoic acid | 0.19 ± 0.11 | 0.21 ± 0.11 |
| Salicylic acid | 0.08 ± 0.11 | 0.11 ± 0.14 |
| Vanillin | 0.49 ± 0.28 | 0.31 ± 0.20 |
| Vanillin acid | 0.18 ± 0.08 | 0.18 ± 0.07 |
| Caffeic acid | 0.01 ± 0.01 | 0.04 ± 0.06 |
| Caffeic acids derivatives | 0.72 a ± 0.14 | 0.50 b ± 0.12 |
| Caffeic acid + Triacontanoic acid | 0.42 ± 0.24 | 0.21 ± 0.11 |
| Caffeic acid + Dotriacontanoic acid | 0.22 ± 0.18 | 0.17 ± 0.25 |
| Caffeic acid + Tetratiacontanoic acid | 0.08 ± 0.00 | 0.12 ± 0.00 |
| Others | 6.84 ± 1.53 | 1.94 ± 0.29 |
| Levoglucosan | 1.11 ± 2.06 | 0.70 ± 0.61 |
| Quinic acid | 4.68 ± 3.98 | - |
| Myo-inositol | 0.51 ± 0.00 | 0.66 ± 0.00 |
| (7E,11E,15E)-3-(methoxymethoxy)-3,7,16,20-tetramethylhenicosa-1,7,11,15,19-pentaene | 0.43 ± 0.06 | 0.45 ± 0.19 |
| Octacosahydro-9,9′-biphenanthrene | 0.11 ± 0.01 | 0.13 ± 0.05 |
| Identified | 96.96 ± 7.90 | 92.04 ± 7.27 |
| Low Attack | Moderate Attack | |
|---|---|---|
| Total phenolics (mg GAE g−1 extract) | 448.6 a ± 101.6 | 296.41 b± 78.1 |
| Total flavonoids (mg CE g−1 extract) | 41.5 a ± 9.4 | 36.7 a ± 6.2 |
| Proanthocyanidins (mg CE g−1 extract) | 10.1 a ± 3.1 | 10.6 a ± 2.7 |
| IC50 (μg extract mL−1) | 13.4 a ± 3.5 | 16.0 a ± 3.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simões, R.; Branco, M.; Nogueira, C.; Carvalho, C.; Santos-Silva, C.; Ferreira-Dias, S.; Miranda, I.; Pereira, H. Phytochemical Composition of Extractives in the Inner Cork Layer of Cork Oaks with Low and Moderate Coraebus undatus Attack. Forests 2022, 13, 1517. https://doi.org/10.3390/f13091517
Simões R, Branco M, Nogueira C, Carvalho C, Santos-Silva C, Ferreira-Dias S, Miranda I, Pereira H. Phytochemical Composition of Extractives in the Inner Cork Layer of Cork Oaks with Low and Moderate Coraebus undatus Attack. Forests. 2022; 13(9):1517. https://doi.org/10.3390/f13091517
Chicago/Turabian StyleSimões, Rita, Manuela Branco, Carla Nogueira, Carolina Carvalho, Conceição Santos-Silva, Suzana Ferreira-Dias, Isabel Miranda, and Helena Pereira. 2022. "Phytochemical Composition of Extractives in the Inner Cork Layer of Cork Oaks with Low and Moderate Coraebus undatus Attack" Forests 13, no. 9: 1517. https://doi.org/10.3390/f13091517
APA StyleSimões, R., Branco, M., Nogueira, C., Carvalho, C., Santos-Silva, C., Ferreira-Dias, S., Miranda, I., & Pereira, H. (2022). Phytochemical Composition of Extractives in the Inner Cork Layer of Cork Oaks with Low and Moderate Coraebus undatus Attack. Forests, 13(9), 1517. https://doi.org/10.3390/f13091517