The Effects of Dynamic Root Distribution on Land–Atmosphere Carbon and Water Fluxes in the Community Earth System Model (CESM1.2.0)


Abstract
:1. Introduction
2. Methods
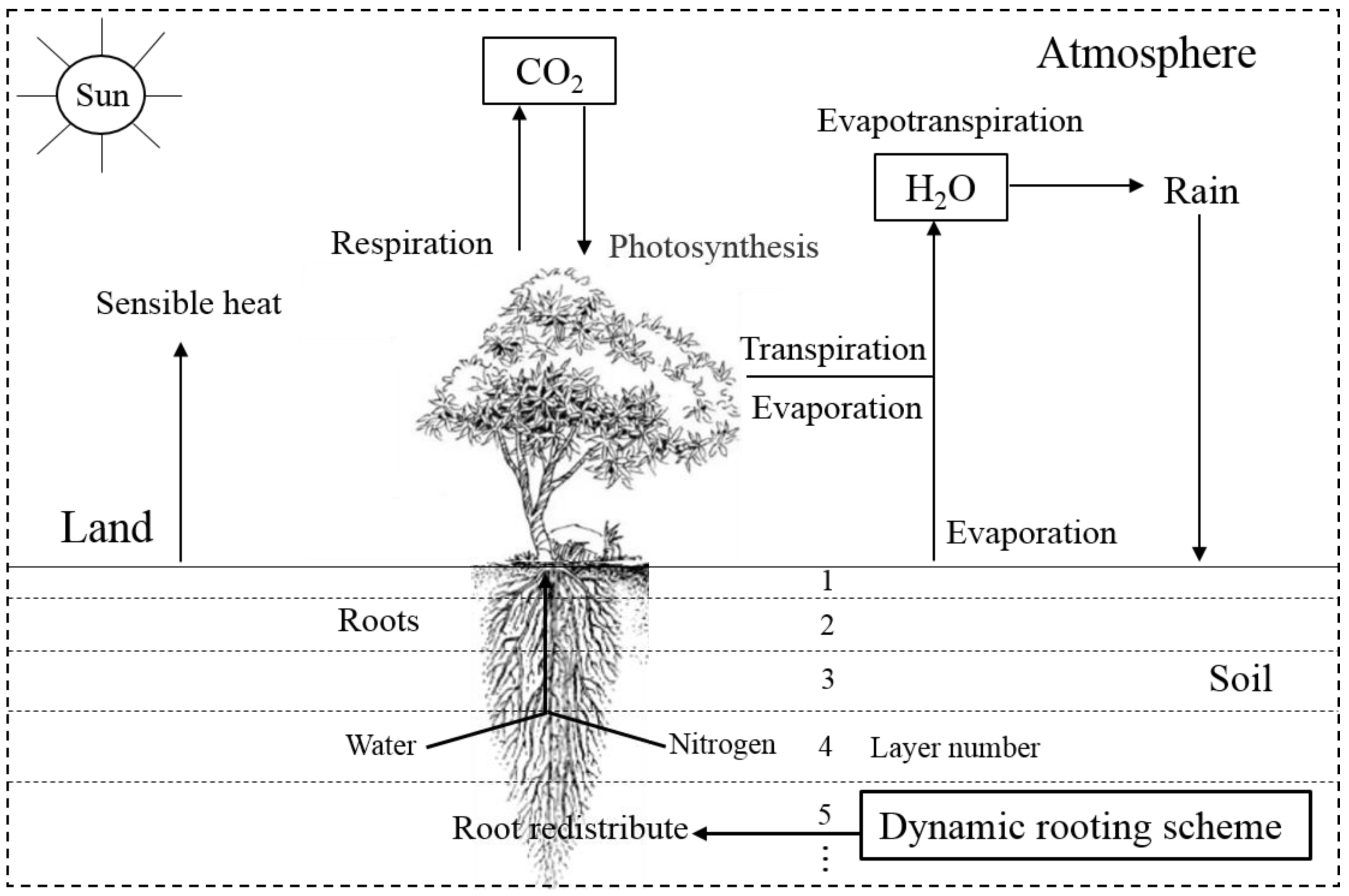
2.1. Model Development
2.1.1. The Community Earth System Model
2.1.2. Dynamic Rooting Scheme and Its Implementation
2.2. Experimental Design
2.3. Mathematical Indices of Model Performance
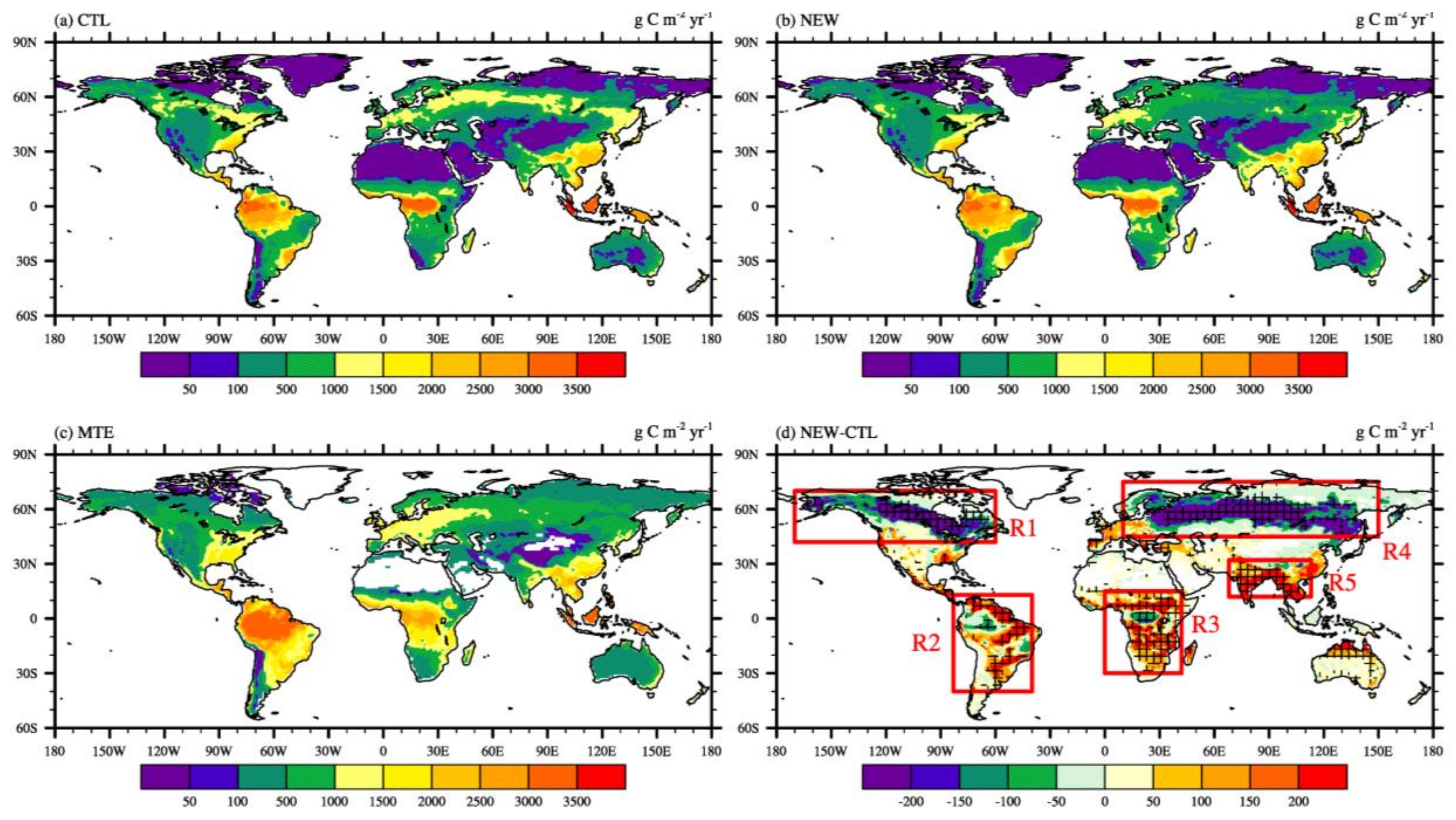
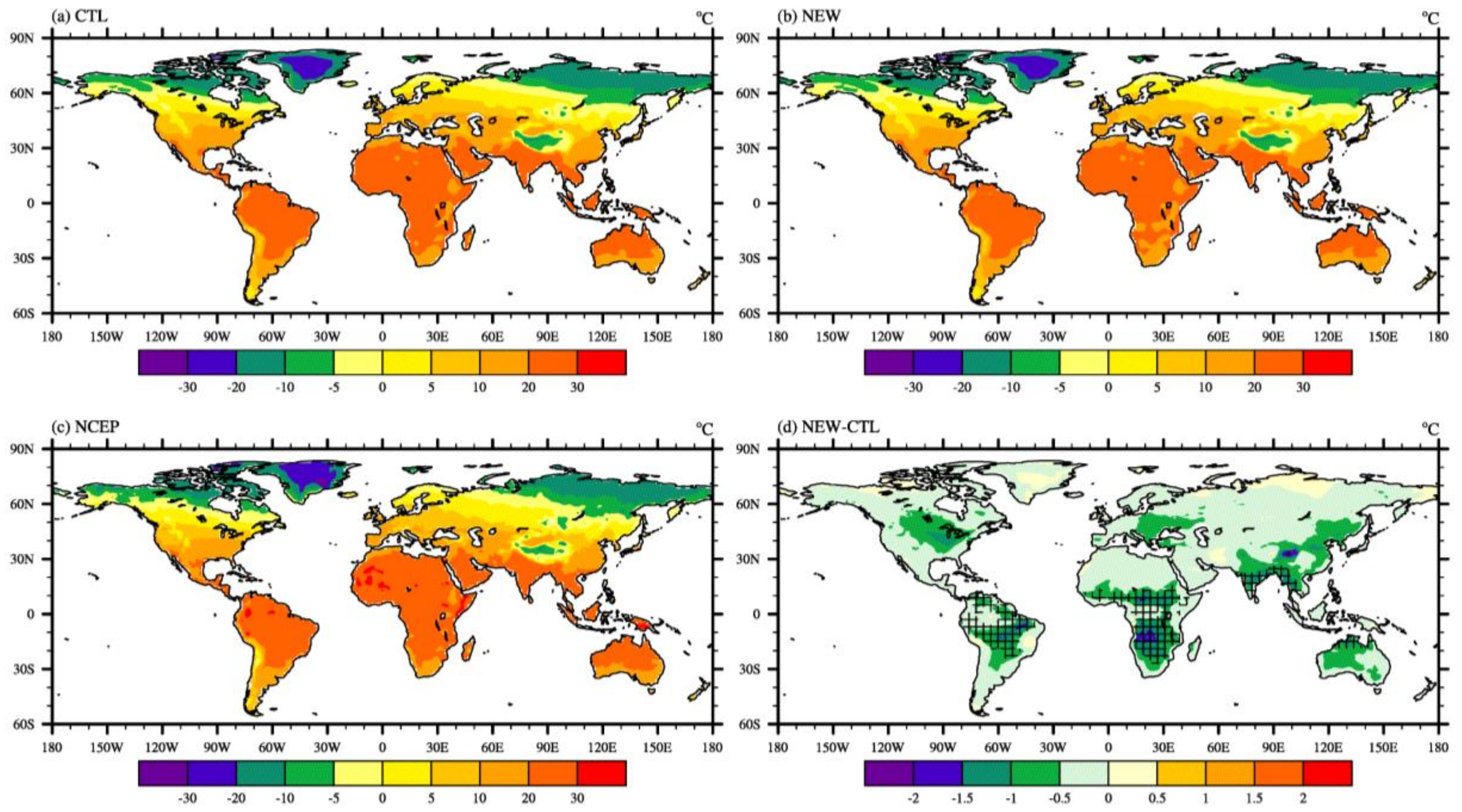
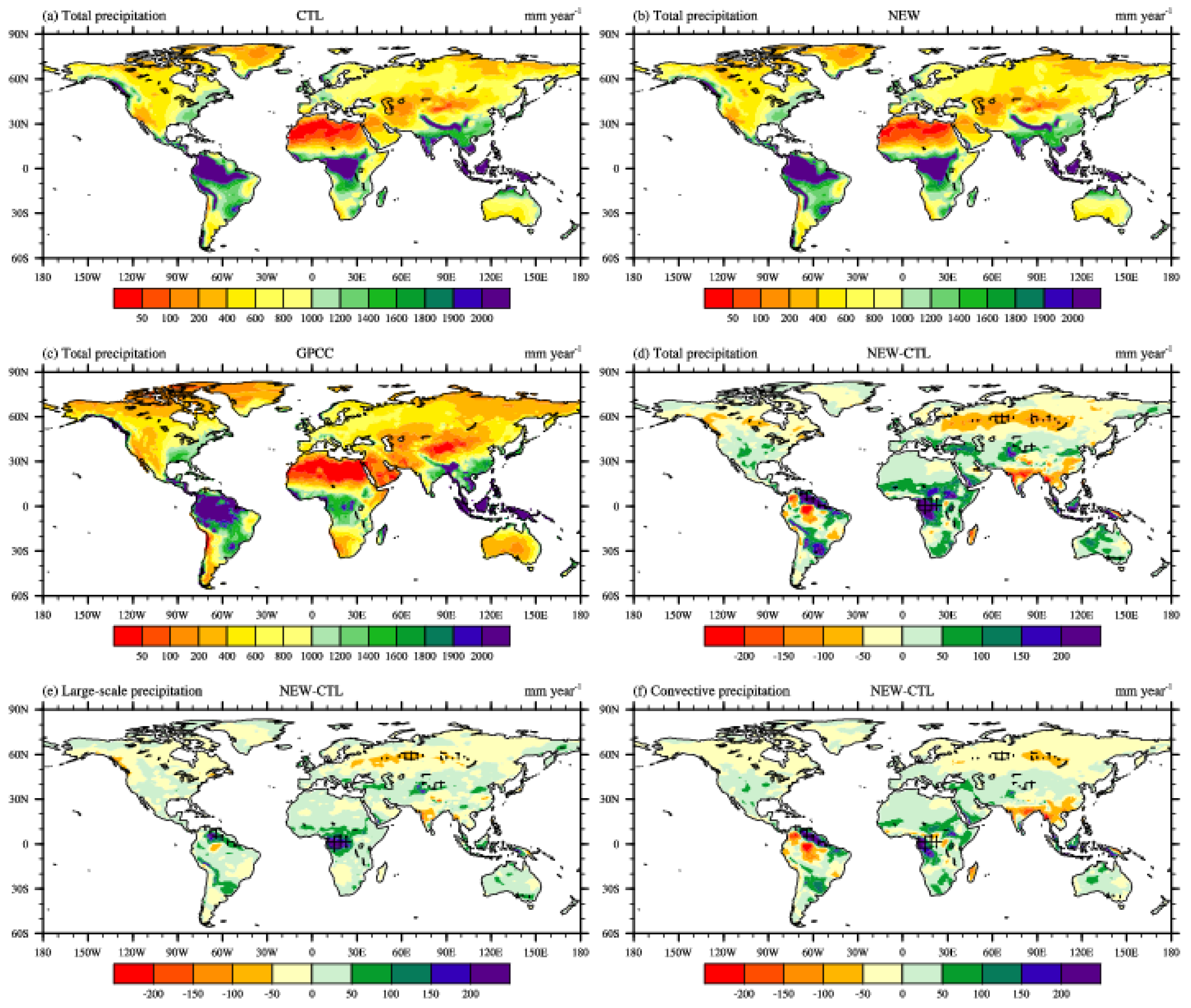
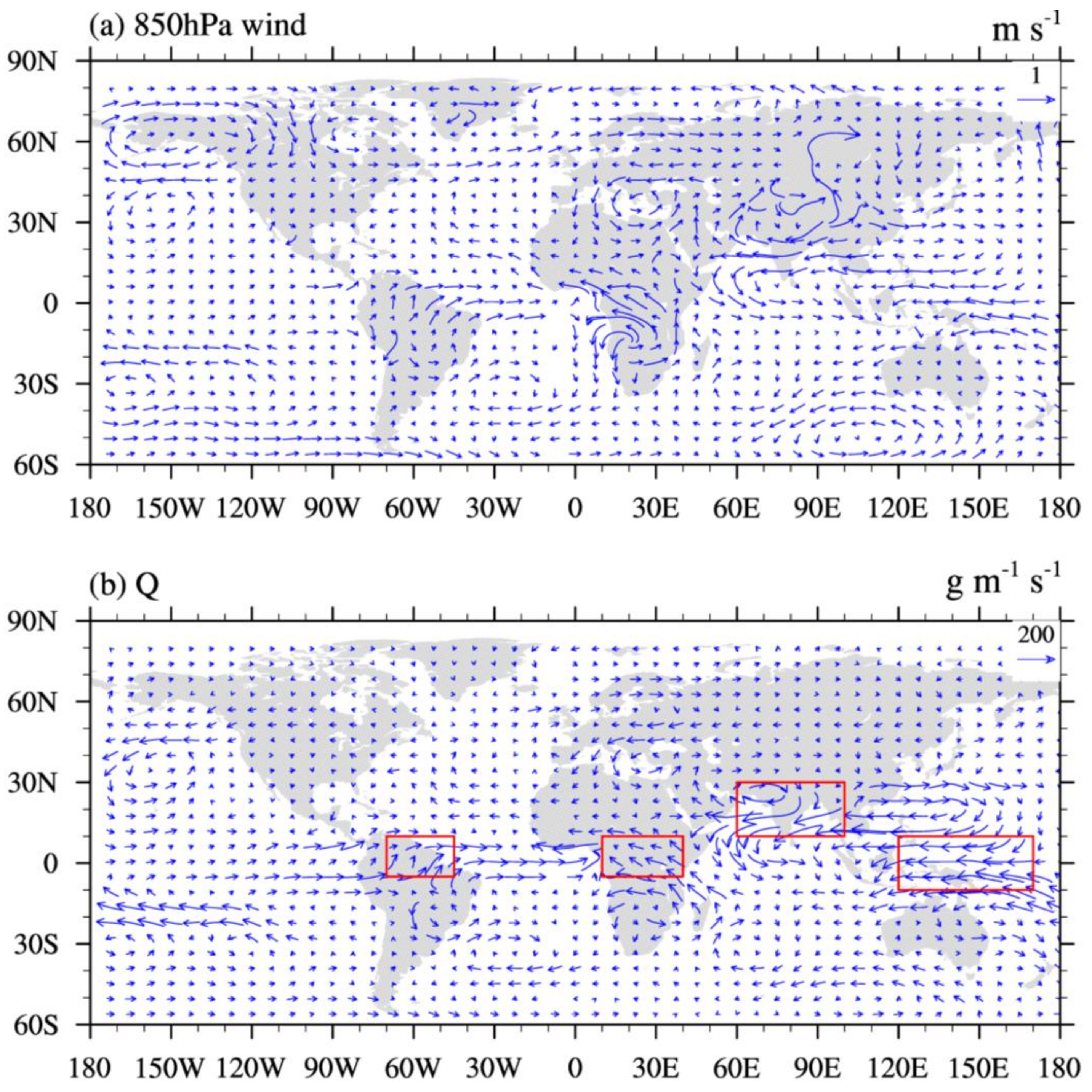
3. Results and Discussion
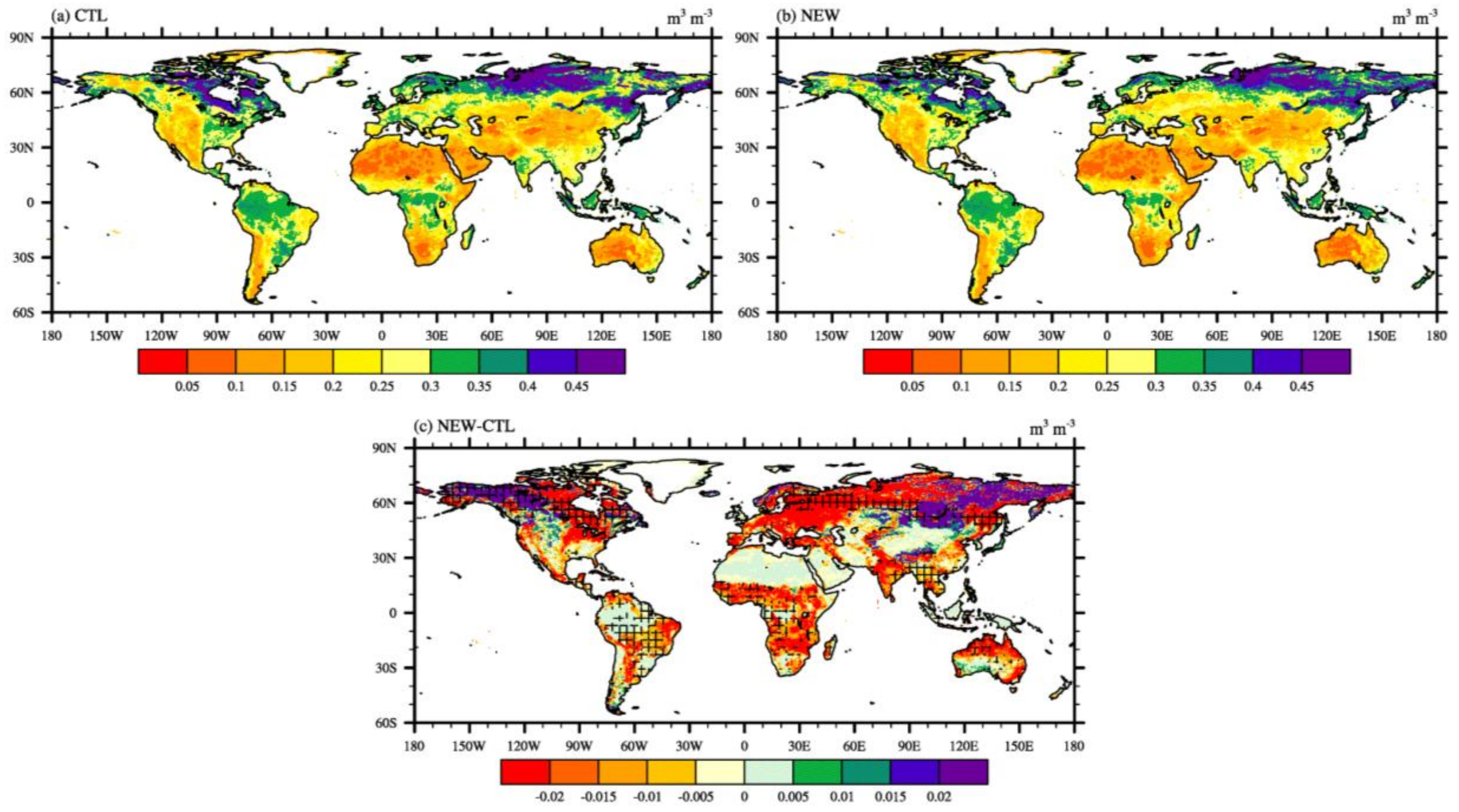
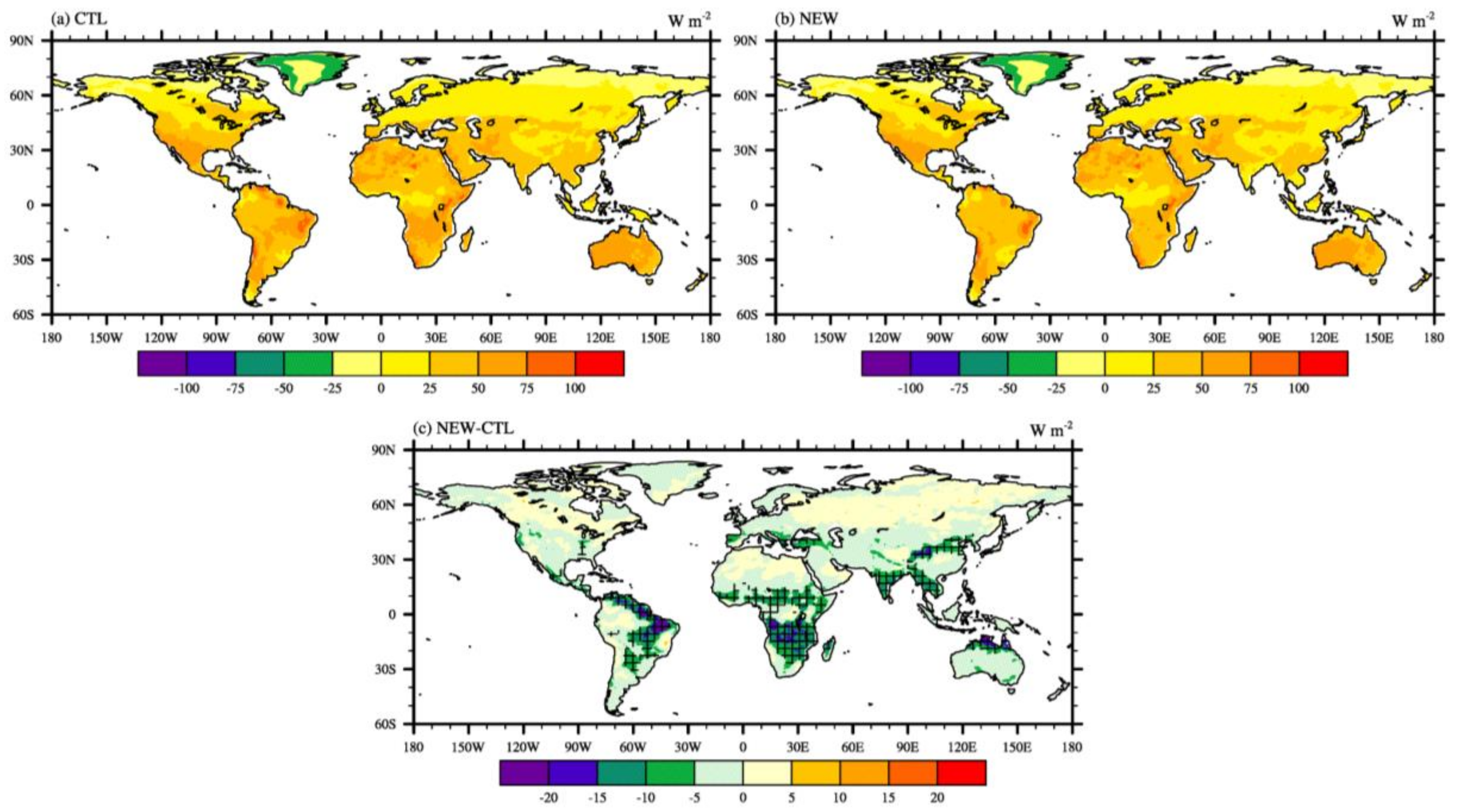
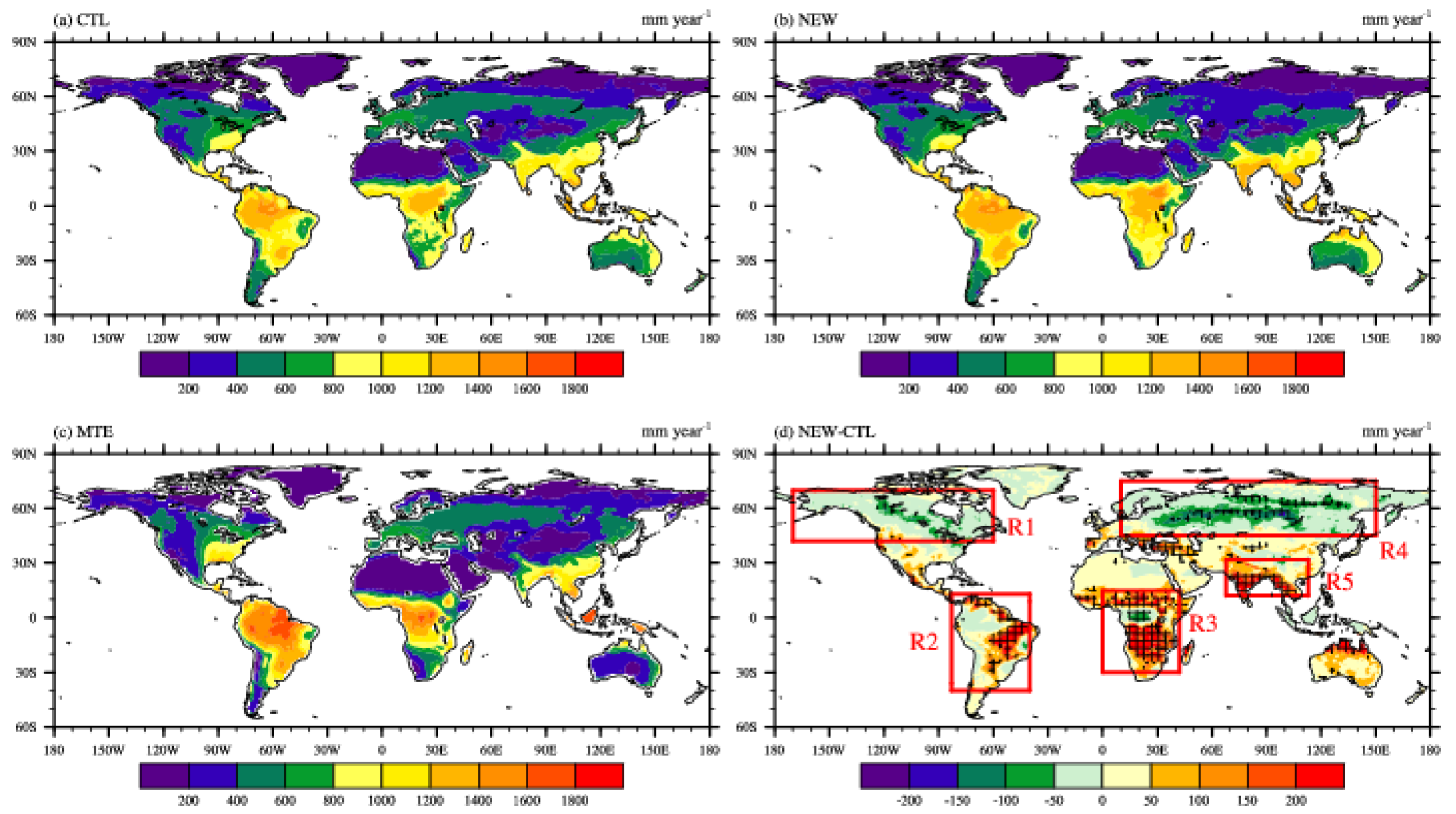
3.1. Effects on Land Variables
3.2. Effects on Atmospheric Variables
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chase, T.N.; Pielke, R.A.; Kittel, T.G.F.; Nemani, R.; Running, S.W. The sensitivity of a general circulation model to global changes in leaf area index. J. Geophys. Res. 1996, 101, 7393–7408. [Google Scholar] [CrossRef]
- Betts, R.A.; Cox, P.M.; Lee, S.E.; Woodward, F.I. Contrasting physiological and structural vegetation feedbacks in climate change simulations. Nature 1997, 387, 796–800. [Google Scholar] [CrossRef]
- Fu, C.B.; Dong, W.J.; Wen, G.; Ye, D.Z.; Ji, J.J.; Lu, J.H.; Yan, X.D. Regional response and adaption to global change. Acta Meteorol. Sin. 2003, 61, 245–249. [Google Scholar] [CrossRef]
- Yuan, F. Hydrological Process Modeling Considering the Effects of Vegetation. Ph.D. Thesis, Hohai University, Nanjing, China, 2006. [Google Scholar]
- Wang, Y.P.; Law, R.M.; Pak, B. A global model of carbon, nitrogen and phosphorus cycles for the terrestrial biosphere. Biogeosciences 2010, 7, 2261–2282. [Google Scholar] [CrossRef] [Green Version]
- Lai, C.; Katul, G. The dynamic role root-water uptake in coupling potential to actual transpiration. Adv. Water Resour. 2000, 23, 427–439. [Google Scholar] [CrossRef]
- Zeng, X.B.; Dai, Y.J.; Dickinson, R.E.; Shaikh, M. The role of root distribution for climate simulation over land. Geophys. Res. Lett. 1998, 25, 4533–4536. [Google Scholar] [CrossRef]
- Feddes, R.A.; Hoff, H.; Bruen, M.; Dawson, T.; Rosnay, P.D.; Dirmeyer, P.; Jackson, R.B.; Kabat, P.; Kleidon, A.; Lilly, A. Modeling root water uptake in hydrological and climate models. Bull. Am. Meteor. Soc. 2001, 82, 2797–2810. [Google Scholar] [CrossRef]
- El Maayar, M.; Sonnentag, O. Crop model validation and sensitivity to climate change scenarios. Clim. Res. 2009, 39, 47–59. [Google Scholar] [CrossRef]
- Bonan, G.B. A Land Surface Model (LSM Version 1.0) for Ecological, Hydrological, and Atmospheric Studies: Technical Description and User’s Guide. Tech. Note NCAR/TN-417-STR; National Center for Atmospheric Research: Boulder, CO, USA, 1996; Available online: https://daac.ornl.gov/data/model_archive/LSM/comp/NCAR_LSM_Users_Guide.pdf (accessed on 1 March 2018).
- Zeng, X.B.; Shaikh, M.; Dai, Y.J.; Dickinson, R.E.; Myneni, R. Coupling of the common land model to the NCAR community climate model. J. Clim. 2002, 15, 1832–1854. [Google Scholar] [CrossRef]
- Ivanov, V.Y.; Bras, R.L.; Vivoni, E.R. Vegetation hydrology dynamics in complex terrain of semiarid areas: 1.A mechanistic approach to modeling dynamic feedbacks. Water Resour. Res. 2008, 44, W03429. [Google Scholar] [CrossRef]
- Nepstad, D.C.; Carvalho, C.R.D.; Davidson, E.A.; Jipp, P.H.; Lefebvre, P.A.; Negreiros, G.H.; da Silva, E.D.; Stone, T.A.; Trumbore, S.E.; Vieira, S. The role of deep roots in the hydrological and carbon cycles of Amazonian forests and pastures. Nature 1994, 372, 666–669. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, R.E.; Shaikh, M.; Bryant, R.; Graumlich, L. Interactive canopies for a climate model. J. Clim. 1998, 11, 2823–2836. [Google Scholar] [CrossRef]
- Barlage, M.; Zeng, X.B. Impact of observed vegetation root distribution on seasonal global simulations of land surface processes. J. Geophys. Res. 2004, 109, D09101. [Google Scholar] [CrossRef]
- Zheng, Z.; Wang, G.L. Modeling the dynamic root water uptake and its hydrological impact at the Reserva Jaru site in Amazonia. J. Geophys. Res. 2007, 112, G04012. [Google Scholar] [CrossRef]
- Jing, C.Q.; Li, L.; Chen, X.; Luo, G.P. Comparison of root water uptake functions to simulate surface energy fluxes within a deep-rooted desert shrub ecosystem. Hydrol. Process. 2014, 28, 5436–5449. [Google Scholar] [CrossRef]
- Warren, J.M.; Hanson, P.J.; Iversen, C.M.; Kumar, J.; Walker, A.P.; Wullschleger, S.D. Root structural and functional dynamics in terrestrial biosphere models-evaluation and recommendations. New Phytol. 2015, 205, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.J.; Kroon, H.D. Foraging in plants: The role of morphological plasticity in resource acquisition. Adv. Ecol. Res. 1994, 25, 159–238. [Google Scholar] [CrossRef]
- Coelho, F.E.; Or, D. A model for soil water and matric potential distribution under drip irrigation with water extraction by roots. Pesqui. Agropecu. Bras. 1999, 34, 225–234. [Google Scholar] [CrossRef]
- Collins, D.B.G.; Bras, R.L. Plant rooting strategies in water-limited ecosystems. Water Resour. Res. 2007, 43, W06407. [Google Scholar] [CrossRef]
- Sivandran, G.; Bras, R.L. Dynamic root distributions in ecohydrological modeling: A case study at Walnut Gulch Experimental Watershed. Water Resour. Res. 2013, 49, 3292–3305. [Google Scholar] [CrossRef]
- McMurtrie, R.E.; Iversen, C.M.; Dewar, R.C.; Medlyn, B.E.; Näsholm, T.; Pepper, D.A.; Norby, R.J. Plant root distributions and nitrogen uptake predicted by a hypothesis of optimal root foraging. Ecol. Evol. 2012, 2, 1235–1250. [Google Scholar] [CrossRef] [PubMed]
- Schenk, H.J.; Jackson, R.B. The global biogeography of roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]
- Schenk, H.J. The shallowest possible water extraction profile: A null model for global root distributions. Vadose Zone J. 2008, 7, 1119–1124. [Google Scholar] [CrossRef]
- Smithwick, E.A.H.; Lucash, M.S.; McCormack, M.L.; Sivandran, G. Improving the representation of roots in terrestrial models. Ecol. Model. 2014, 291, 193–204. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Xie, Z.H.; Jia, B.H. Incorporation of a Dynamic Root Distribution into CLM4.5: Evaluation of Carbon and Water fluxes over the Amazon. Adv. Atmos. Sci. 2016, 33, 1047–1060. [Google Scholar] [CrossRef]
- Hurrell, J.W.; Holland, M.M.; Gent, P.R.; Ghan, S.; Kay, J.E.; Kushner, P.J.; Lamarque, J.F.; Large, W.G.; Lawrence, D.; Lindsay, K. The Community Earth System Model: A framework for collaborative research. Bull. Am. Meteor. Soc. 2013, 94, 1339–1360. [Google Scholar] [CrossRef]
- Neale, R.B.; Chen, C.C.; Gettelman, A.; Lauritzen, P.H.; Park, S.; Williamson, D.L.; Conley, A.J.; Garcia, R.; Kinnison, D.; Lamarque, J.F. Description of the NCAR Community Atmosphere Model (CAM 4.0). NCAR Tech. Note NCAR/TN-4851STR. 2010. Available online: http://www.cesm.ucar.edu/models/cesm1.0/cam/docs/description/cam4_desc.pdf (accessed on 1 March 2018).
- Neale, R.B.; Richter, J.; Park, S.; Lauritzen, P.H.; Vavrus, S.J.; Rasch, P.J.; Zhang, M.H. The mean climate of the Community Atmosphere Model (CAM4) in forced SST and fully coupled experiments. J. Clim. 2013, 26, 5150–5168. [Google Scholar] [CrossRef]
- Lawrence, D.M.; Oleson, K.W.; Flanner, M.G.; Thornton, P.E.; Swenson, S.C.; Lawrence, P.J.; Zeng, X.; Yang, Z.L.; Levis, S.; Sakaguchi, K. Parameterization Improvements and Functional and Structural Advances in Version 4 of the Community Land Model. J. Adv. Model. Earth Syst. 2011, 3, M03001. [Google Scholar] [CrossRef]
- Oleson, K.W.; Lawrence, D.M.; Bonan, G.B.; Drewniak, B.; Huang, M.; Koven, C.D.; Levis, S.; Li, F.; Riley, J.; Subin, M. Technical Description of Version 4.5 of the Community Land Model (CLM). NCAR Tech. Note NCAR/TN-503+STR; National Center for Atmospheric Research: Boulder, CO, USA, 2013; p. 420. [Google Scholar]
- Zeng, X.B. Global vegetation root distribution for land modeling. J. Hydrometeorol. 2001, 2, 525–530. [Google Scholar] [CrossRef]
- Shangguan, W.; Dai, Y.J.; Duan, Q.Y.; Liu, B.Y.; Yuan, H. A global soil data set for earth system modeling. J. Adv. Model. Earth Syst. 2014, 6, 249–263. [Google Scholar] [CrossRef]
- Rayner, N.A.; Parker, D.E.; Horton, E.B.; Folland, C.K.; Alexander, L.V.; Rowell, D.P.; Kent, E.C.; Kaplan, A. Global analyses of sea surface temperature, sea ice, and night arine air temperature since the late nineteenth century. Geophys. Res. 2003, 108, 4407. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Bondeau, A. Towards global empirical upscaling of FLUXNET eddy covariance observations: Validation of a model tree ensemble approach using a biosphere model. Biogeosciences 2009, 6, 2001–2013. [Google Scholar] [CrossRef]
- Jung, M.; Reichstein, M.; Margolis, H.A.; Cescatti, A.; Richardson, A.D.; Arain, M.A.; Arneth, A.; Bernhofer, C.; Bonal, D.; Chen, J. Global patterns of land-atmosphere fluxes of carbon dioxide, latent heat, and sensible heat derived from eddy covariance, satellite, and meteorological observations. J. Geophys. Res. 2011, 116, G00J07. [Google Scholar] [CrossRef]
- Shi, X.L.; Zhang, F.; Zhou, W.Y.; Zhang, Y. Impacts of CG-LTDR Land Cover Dataset Updates on the Ground Temperature Simulation with BCC_AVIM 1.0. J. Geo-Inf. Sci. 2015, 11, 1294–1303. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Ryel, R.J.; Caldwell, M.M.; Yoder, C.K.; Or, D.; Leffler, A. Hydraulic redistribution in a stand of Artemisia tridentate: Evaluation of benefits to transpiration assessed with a simulation model. Oecologia 2002, 130, 173–184. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| R1 | R2 | R3 | R4 | R5 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CTL | NEW | CTL | NEW | CTL | NEW | CTL | NEW | CTL | NEW | |
| R | 0.86 * | 0.88* | 0.75 | 0.77 | 0.59 | 0.60 | 0.85 * | 0.86 * | 0.76 | 0.77 |
| MBE | −37.61 | −20.89 | −208.6 | −98.4 | −60.1 | −30.3 | −18.1 | −36.6 | 70.2 | 81.5 |
| RMSE | 51.3 | 35.7 | 226.5 | 130.1 | 151.3 | 95.3 | 41.3 | 91.5 | 112.1 | 191.3 |
| R1 | R2 | R3 | R4 | R5 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CTL | NEW | CTL | NEW | CTL | NEW | CTL | NEW | CTL | NEW | |
| R | 0.98 * | 0.98 * | 0.70 | 0.82 | 0.60 | 0.50 | 0.98 * | 0.98 * | 0.96 * | 0.97 * |
| MBE | 1.09 | 0.05 | −7.28 | −5.04 | −2.41 | −1.21 | 2.77 | 0.06 | −1.17 | 0.03 |
| RMSE | 2.29 | 0.25 | 8.01 | 6.06 | 2.44 | 1.25 | 2.79 | 0.46 | 1.22 | 0.30 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Jia, B.; Xie, Z. The Effects of Dynamic Root Distribution on Land–Atmosphere Carbon and Water Fluxes in the Community Earth System Model (CESM1.2.0). Forests 2018, 9, 172. https://doi.org/10.3390/f9040172
Wang Y, Jia B, Xie Z. The Effects of Dynamic Root Distribution on Land–Atmosphere Carbon and Water Fluxes in the Community Earth System Model (CESM1.2.0). Forests. 2018; 9(4):172. https://doi.org/10.3390/f9040172
Chicago/Turabian StyleWang, Yuanyuan, Binghao Jia, and Zhenghui Xie. 2018. "The Effects of Dynamic Root Distribution on Land–Atmosphere Carbon and Water Fluxes in the Community Earth System Model (CESM1.2.0)" Forests 9, no. 4: 172. https://doi.org/10.3390/f9040172
APA StyleWang, Y., Jia, B., & Xie, Z. (2018). The Effects of Dynamic Root Distribution on Land–Atmosphere Carbon and Water Fluxes in the Community Earth System Model (CESM1.2.0). Forests, 9(4), 172. https://doi.org/10.3390/f9040172