ALV Integration-Associated Hypomethylation at the TERT Promoter Locus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Tumor Induction and Samples
2.2. DNA Extraction and Bisulfite Treatment
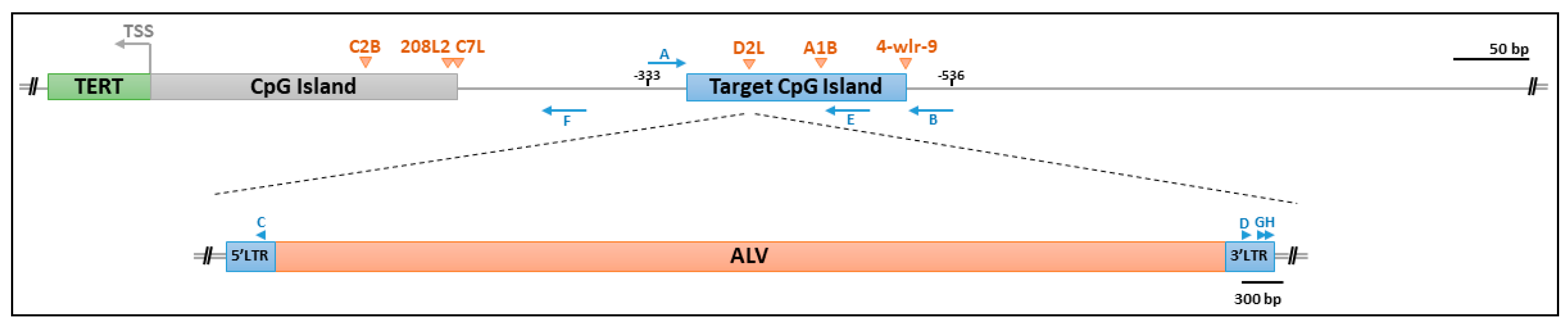
2.3. PCR Amplification and Sequencing Analysis
3. Results
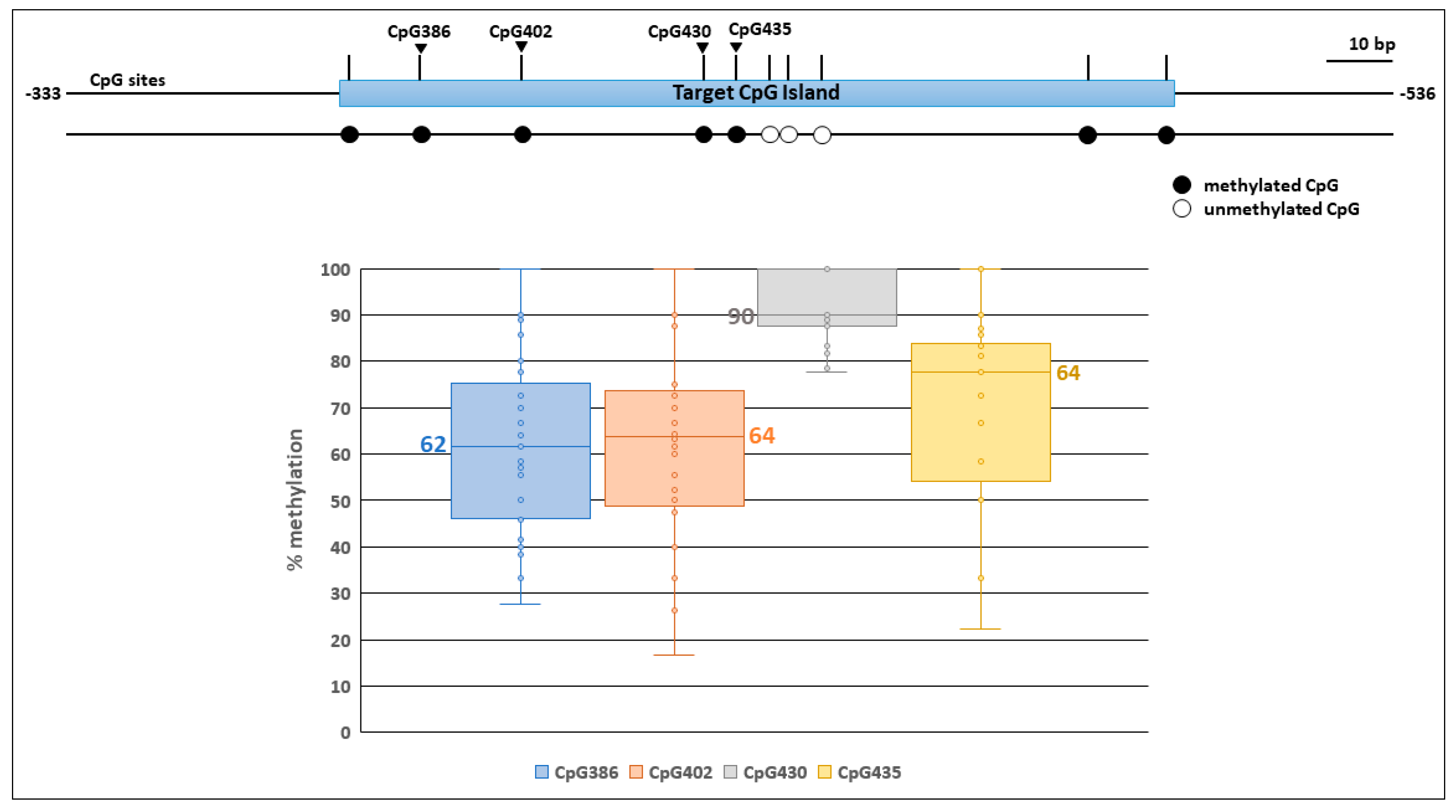
3.1. Chicken TERT Promoters Were Significantly Methylated in Unoccupied Alleles
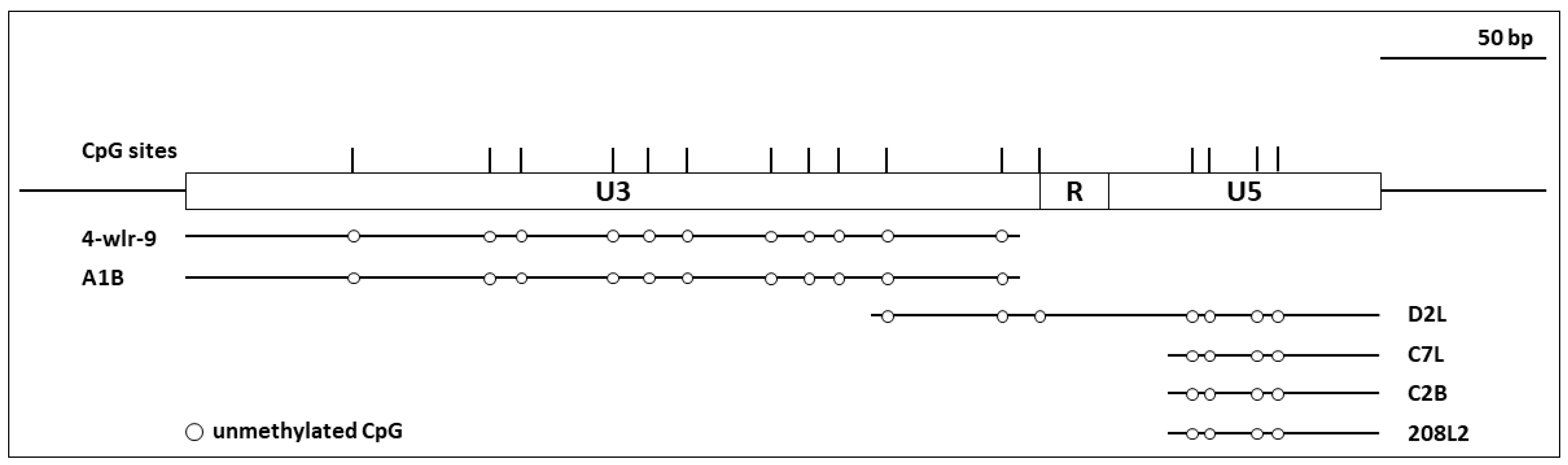
3.2. ALV LTRs of Proviruses in the TERT Promoter Were Not Methylated
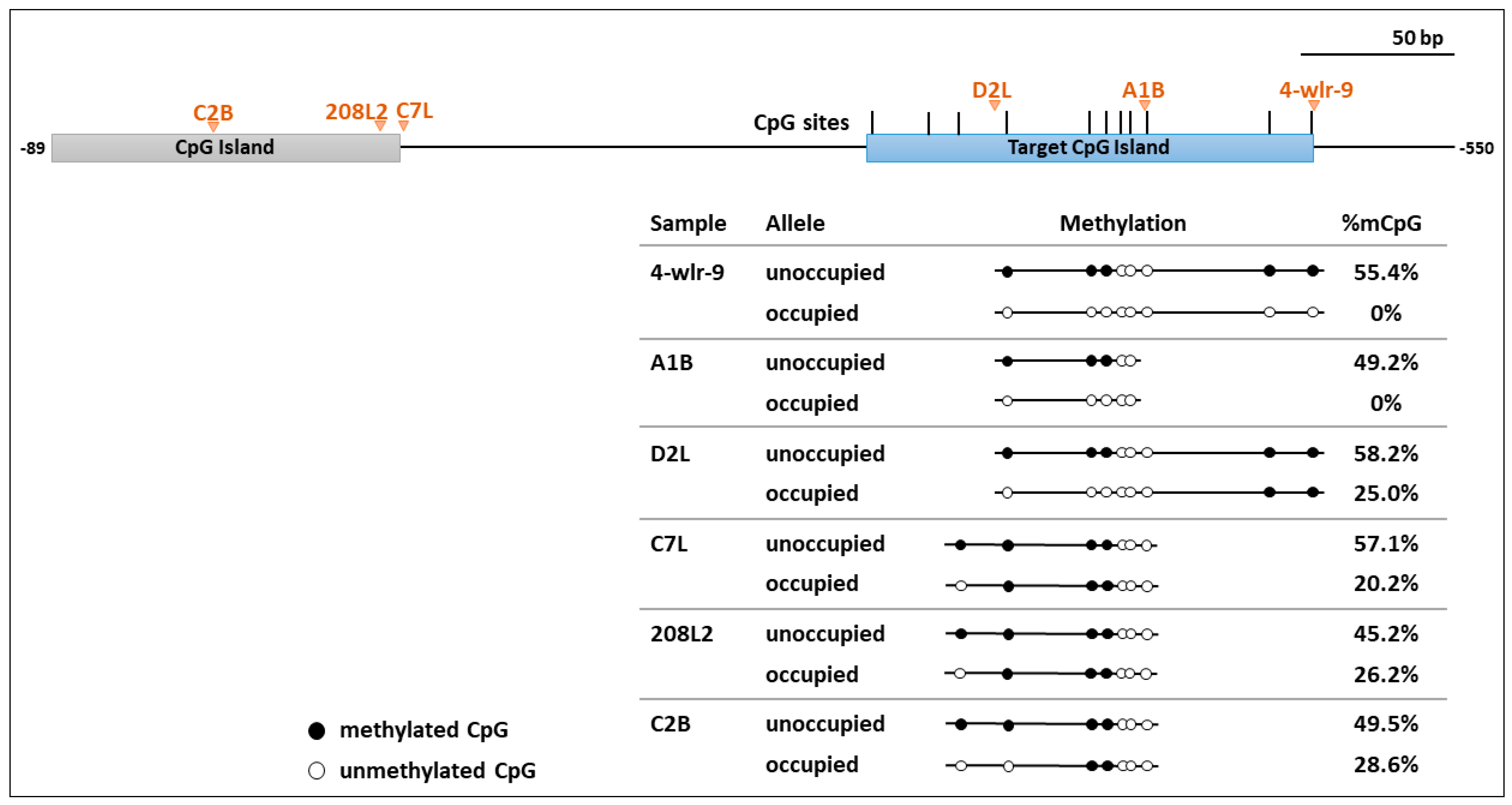
3.3. Occupied Alleles in the TERT Promoter had Decreased Methylation in Flanking Host DNA
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Beemon, K.; Rosenberg, N. Mechanisms of Oncogenesis by Avian and Murine Retroviruses; Springer: New York, NY, USA, 2012; ISBN 9781461400165. [Google Scholar]
- Justice, J.; Beemon, K.L. Avian retroviral replication. Curr. Opin. Virol. 2013, 3, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Justice, J.F.; Morgan, R.W.; Beemon, K.L. Common Viral Integration Sites Identified in Avian Leukosis Virus-Induced B-Cell Lymphomas. MBio 2015, 6, e01863-15. [Google Scholar] [CrossRef] [PubMed]
- Hayward, W.S.; Neel, B.G.; Astrin, S.M. Activation of a cellular ONC gene by promoter insertion in ALV-induced lymphoid leukosis. Nature 1981, 290, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.W.; Humphries, E.H. Selective integration of avian leukosis virus in different hematopoietic tissues. Virology 1986, 155, 557–566. [Google Scholar] [CrossRef]
- Clurman, B.E.; Hayward, W.S. Multiple proto-oncogene activations in avian leukosis virus-induced lymphomas: Evidence for stage-specific events. Mol. Cell. Biol. 1989, 9, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Xian, R.R.; Li, Y.; Polony, T.S.; Beemon, K.L. Telomerase reverse transcriptase expression elevated by avian leukosis virus integration in B cell lymphomas. Proc. Natl. Acad. Sci. USA 2007, 104, 18952–18957. [Google Scholar] [CrossRef] [PubMed]
- Nehyba, J.; Malhotra, S.; Winans, S.; O’Hare, T.H.; Justice, J.; Beemon, K. Avian Leukosis Virus Activation of an Antisense RNA Upstream of TERT in B-Cell Lymphomas. J. Virol. 2016, 90, 9509–9517. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.; Winans, S.; Lam, G.; Justice, J.; Morgan, R.; Beemon, K. Selection for avian leukosis virus integration sites determines the clonal progression of B-cell lymphomas. PLoS Pathog. 2017, 13, e1006708. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Blasco, M.A. Telomeres and human disease: Ageing, cancer and beyond. Nat. Rev. Genet. 2005, 6, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Bryan, T.M.; Englezou, A.; Gupta, J.; Bacchetti, S.; Reddel, R.R. Telomere elongation in immortal human cells without detectable telomerase activity. EMBO J. 1995, 14, 4240–4248. [Google Scholar] [PubMed]
- Bryan, T.M.; Englezou, A.; Dalla-Pozza, L.; Dunham, M.A.; Reddel, R.R. Evidence for an alternative mechanism for maintaining telomere length in human tumors and tumor-derived cell lines. Nat. Med. 1997, 3, 1271–1274. [Google Scholar] [CrossRef] [PubMed]
- O’Hare, T.H.; Delany, M.E. Molecular and cellular evidence for the alternative lengthening of telomeres (ALT) mechanism in chicken. Cytogenet. Genome Res. 2011, 135, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Delany, M.E.; Daniels, L.M. The chicken telomerase reverse transcriptase (chTERT): Molecular and cytogenetic characterization with a comparative analysis. Gene 2004, 339, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhao, Y.; Wang, S. Chromatin and epigenetic regulation of the telomerase reverse transcriptase gene. Protein Cell 2010, 1, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Barthel, F.P.; Wei, W.; Tang, M.; Martinez-Ledesma, E.; Hu, X.; Amin, S.B.; Akdemir, K.C.; Seth, S.; Song, X.; Wang, Q.; et al. Systematic analysis of telomere length and somatic alterations in 31 cancer types. Nat. Genet. 2017, 49, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Horn, S.; Figl, A.; Rachakonda, P.S.; Fischer, C.; Sucker, A.; Gast, A.; Kadel, S.; Moll, I.; Nagore, E.; Hemminki, K.; et al. TERT Promoter Mutations in Familial and Sporadic Melanoma. Science 2013, 339, 959–961. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Hodis, E.; Xu, M.J.; Kryukov, G.V.; Chin, L.; Garraway, L.A. Highly Recurrent TERT Promoter Mutations in Human Melanoma. Science 2013, 339, 957–959. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Li, N.; Hu, X.; Li, J.; Du, Z.; Chen, L.; Yin, G.; Duan, J.; Zhang, H.; Zhao, Y.; et al. Genome-Wide Mapping of DNA Methylation in Chicken. PLoS ONE 2011, 6, e19428. [Google Scholar] [CrossRef] [PubMed]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Schübeler, D. Genomic patterns of DNA methylation: Targets and function of an epigenetic mark. Curr. Opin. Cell Biol. 2007, 19, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Devereux, T.R.; Horikawa, I.; Anna, C.H.; Annab, L.A.; Afshari, C.A.; Barrett, J.C. DNA methylation analysis of the promoter region of the human telomerase reverse transcriptase (hTERT) gene. Cancer Res. 1999, 59, 6087–6090. [Google Scholar] [PubMed]
- Dessain, S.K.; Yu, H.; Reddel, R.R.; Beijersbergen, R.L.; Weinberg, R.A. Methylation of the human telomerase gene CpG island. Cancer Res. 2000, 60, 537–541. [Google Scholar] [PubMed]
- Nomoto, K.; Maekawa, M.; Sugano, K.; Ushiama, M.; Fukayama, N.; Fujita, S.; Kakizoe, T. Methylation Status and Expression of Human Telomerase Reverse Transcriptase mRNA in Relation to Hypermethylation of the p16 gene in Colorectal Cancers as Analyzed by Bisulfite PCR-SSCP. Jpn. J. Clin. Oncol. 2002, 32, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Guilleret, I.; Yan, P.; Grange, F.; Braunschweig, R.; Bosman, F.T.; Benhattar, J. Hypermethylation of the human telomerase catalytic subunit (hTERT) gene correlates with telomerase activity. Int. J. Cancer 2002, 101, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Guilleret, I.; Benhattar, J. Unusual distribution of DNA methylation within the hTERT CpG island in tissues and cell lines. Biochem. Biophys. Res. Commun. 2004, 325, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Saldanha, S.N.; Pate, M.S.; Andrews, L.G.; Tollefsbol, T.O. Epigenetic regulation of human telomerase reverse transcriptase promoter activity during cellular differentiation. Genes Chromosom. Cancer 2004, 41, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, N.G.; Poole, J.C.; Saldanha, S.N.; Hansen, N.J.; Key, J.S.; Pita, M.A.; Andrews, L.G.; Tollefsbol, T.O. Control mechanisms in the regulation of telomerase reverse transcriptase expression in differentiating human teratocarcinoma cells. Biochem. Biophys. Res. Commun. 2003, 306, 650–659. [Google Scholar] [CrossRef]
- Shin, K.; Kang, M.; Dicterow, E.; Park, N.H. Hypermethylation of the hTERT promoter inhibits the expression of telomerase activity in normal oral fibroblasts and senescent normal oral. Br. J. Cancer 2003, 89, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Zinn, R.L.; Pruitt, K.; Eguchi, S.; Baylin, S.B.; Herman, J.G. hTERT Is Expressed in Cancer Cell Lines Despite Promoter DNA Methylation by Preservation of Unmethylated DNA and Active Chromatin around the Transcription Start Site. Cancer Res. 2007, 67, 194–201. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.L.; Paucek, R.D.; Huang, F.W.; Ghandi, M.; Nwumeh, R.; Costello, J.C.; Cech, T.R. Allele-Specific DNA Methylation and Its Interplay with Repressive Histone Marks at Promoter-Mutant TERT Genes. Cell Rep. 2017, 21, 3700–3707. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, J. Presence of Chicken Tumour Virus in the Sarcoma of the Adult Rat inoculated after Birth with Rous Sarcoma Tissue. Nature 1960, 186, 980–981. [Google Scholar] [CrossRef] [PubMed]
- Guntaka, R.V.; Rao, P.Y.; Mitsialis, S.A.; Katz, R. Modification of avian sarcoma proviral DNA sequences in nonpermissive XC cells but not in permissive chicken cells. J. Virol. 1980, 34, 569–572. [Google Scholar] [PubMed]
- Svoboda, J.; Hejnar, J.; Geryk, J.; Elleder, D.; Vernerová, Z. Retroviruses in foreign species and the problem of provirus silencing. Gene 2000, 261, 181–188. [Google Scholar] [CrossRef]
- Hejnar, J.; Plachý, J.; Geryk, J.; Machon, O.; Trejbalová, K.; Guntaka, R.V.; Svoboda, J. Inhibition of the rous sarcoma virus long terminal repeat-driven transcription by in vitro methylation: Different sensitivity in permissive chicken cells versus mammalian cells. Virology 1999, 255, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Shalginskikh, N.; Poleshko, A.; Skalka, A.M.; Katz, R.A. Retroviral DNA methylation and epigenetic repression are mediated by the antiviral host protein Daxx. J. Virol. 2013, 87, 2137–2150. [Google Scholar] [CrossRef] [PubMed]
- Hejnar, J.; Elleder, D.; Hájková, P.; Walter, J.; Blazková, J.; Svoboda, J. Demethylation of host-cell DNA at the site of avian retrovirus integration. Biochem. Biophys. Res. Commun. 2003, 311, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Jähner, D.; Jaenisch, R. Retrovirus-induced de novo methylation of flanking host sequences correlates with gene inactivity. Nature 1985, 315, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Senigl, F.; Auxt, M.; Hejnar, J. Transcriptional provirus silencing as a crosstalk of de novo DNA methylation and epigenomic features at the integration site. Nucleic Acids Res. 2012, 40, 5298–5312. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Yamamoto, H.; Oikawa, R.; Toyota, M.; Yamamoto, M.; Kokudo, N.; Tanaka, S.; Arii, S.; Yotsuyanagi, H.; Koike, K.; et al. DNA methylation at hepatitis B viral integrants is associated with methylation at flanking human genomic sequences. Genome Res. 2015, 25, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Hatano, T.; Sano, D.; Takahashi, H.; Hyakusoku, H.; Isono, Y.; Shimada, S.; Sawakuma, K.; Takada, K.; Oikawa, R.; Watanabe, Y.; et al. Identification of human papillomavirus (HPV) 16 DNA integration and the ensuing patterns of methylation in HPV-associated head and neck squamous cell carcinoma cell lines. Int. J. Cancer 2017, 140, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Forsyth, N.R.; Wright, W.E.; Shay, J.W. Telomerase and differentiation in multicellular organisms: Turn it off, turn it on, and turn it off again. Differentiation 2002, 69, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Swanberg, S.E.; O’Hare, T.H.; Robb, E.A.; Robinson, C.M.; Chang, H.; Delany, M.E. Telomere biology of the chicken: A model for aging research. Exp. Gerontol. 2010, 45, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.A.; Delany, M.E. Ontogeny of telomerase in chicken: Impact of downregulation on pre- and postnatal telomere length in vivo. Dev. Growth Differ. 2000, 42, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Swanberg, S.E.; Delany, M.E. Dynamics of telomere erosion in transformed and non-transformed avian cells in vitro. Cytogenet. Genome Res. 2003, 102, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Polony, T.S.; Bowers, S.J.; Neiman, P.E.; Beemon, K.L. Silent point mutation in an avian retrovirus RNA processing element promotes c-myb-associated short-latency lymphomas. J. Virol. 2003, 77, 9378–9387. [Google Scholar] [CrossRef] [PubMed]
- Pappas, J.J.; Toulouse, A.; Bradley, W.E.C. The bisulfite genomic sequencing protocol. Adv. Lung Cancer 2013, 2, 21–25. [Google Scholar] [CrossRef]
- Stern, J.L.; Theodorescu, D.; Vogelstein, B.; Papadopoulos, N.; Cech, T.R. Mutation of the TERT promoter, switch to active chromatin, and monoallelic TERT expression in multiple cancers. Genes Dev. 2015, 29, 2219–2224. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.W.; Bielski, C.M.; Rinne, M.L.; Hahn, W.C.; Sellers, W.R.; Stegmeier, F.; Garraway, L.A.; Kryukov, G.V. TERT promoter mutations and monoallelic activation of TERT in cancer. Oncogenesis 2015, 4, e176. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, P.; Leão, R.; Lipman, T.; Campbell, B.; Lee, D.; Price, A.; Zhang, C.; Heidari, A.; Stephens, D.; Boerno, S.; et al. A cancer specific hypermethylation signature of the TERT promoter predicts biochemical relapse in prostate cancer: A retrospective cohort study. Oncotarget 2016, 7, 57726–57736. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, P.; Choufani, S.; Mack, S.; Gallagher, D.; Zhang, C.; Lipman, T.; Zhukova, N.; Walker, E.J.; Martin, D.; Merino, D.; et al. Methylation of the TERT promoter and risk stratification of childhood brain tumours: An integrative genomic and molecular study. Lancet Oncol. 2013, 14, 534–542. [Google Scholar] [CrossRef]
- Engdahl, E.; Dunn, N.; Niehusmann, P.; Wideman, S.; Wipfler, P.; Becker, A.J.; Ekström, T.J.; Almgren, M.; Fogdell-Hahn, A. Human Herpesvirus 6B Induces Hypomethylation on Chromosome 17p13.3, Correlating with Increased Gene Expression and Virus Integration. J. Virol. 2017, 91, e02105-16. [Google Scholar] [CrossRef] [PubMed]
- Buendia, M.-A.; Neuveut, C. Hepatocellular Carcinoma. Cold Spring Harb. Perspect. Med. 2015, 5, a021444. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Group | Tissue Type |
|---|---|
| Normal control tissues | normal bursa (NB), normal kidney (NK), normal liver (NL), normal spleen (NS), C2K |
| Chicken cell culture | primary chicken embryo fibroblast (CEF), bursal lymphoma cell line (DT-40) |
| Tumors without telomerase reverse transcriptase (TERT) promoter integration | 205L1, 209L, 214L4, 215K1, 218K, 796L, 223K, 791L1, B8L |
| Tumors with TERT promoter integration | 206L1, 208L2, A1B, C2B, C6L, C7B, C7L, D2L, 4-w-lr-9 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, G.; Beemon, K. ALV Integration-Associated Hypomethylation at the TERT Promoter Locus. Viruses 2018, 10, 74. https://doi.org/10.3390/v10020074
Lam G, Beemon K. ALV Integration-Associated Hypomethylation at the TERT Promoter Locus. Viruses. 2018; 10(2):74. https://doi.org/10.3390/v10020074
Chicago/Turabian StyleLam, Gary, and Karen Beemon. 2018. "ALV Integration-Associated Hypomethylation at the TERT Promoter Locus" Viruses 10, no. 2: 74. https://doi.org/10.3390/v10020074
APA StyleLam, G., & Beemon, K. (2018). ALV Integration-Associated Hypomethylation at the TERT Promoter Locus. Viruses, 10(2), 74. https://doi.org/10.3390/v10020074