Bovine Vaccinia: Insights into the Disease in Cattle

 , and
, and {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Bovine Vaccinia Pathogenesis: Evidence of a Systemic and Persistent Infection
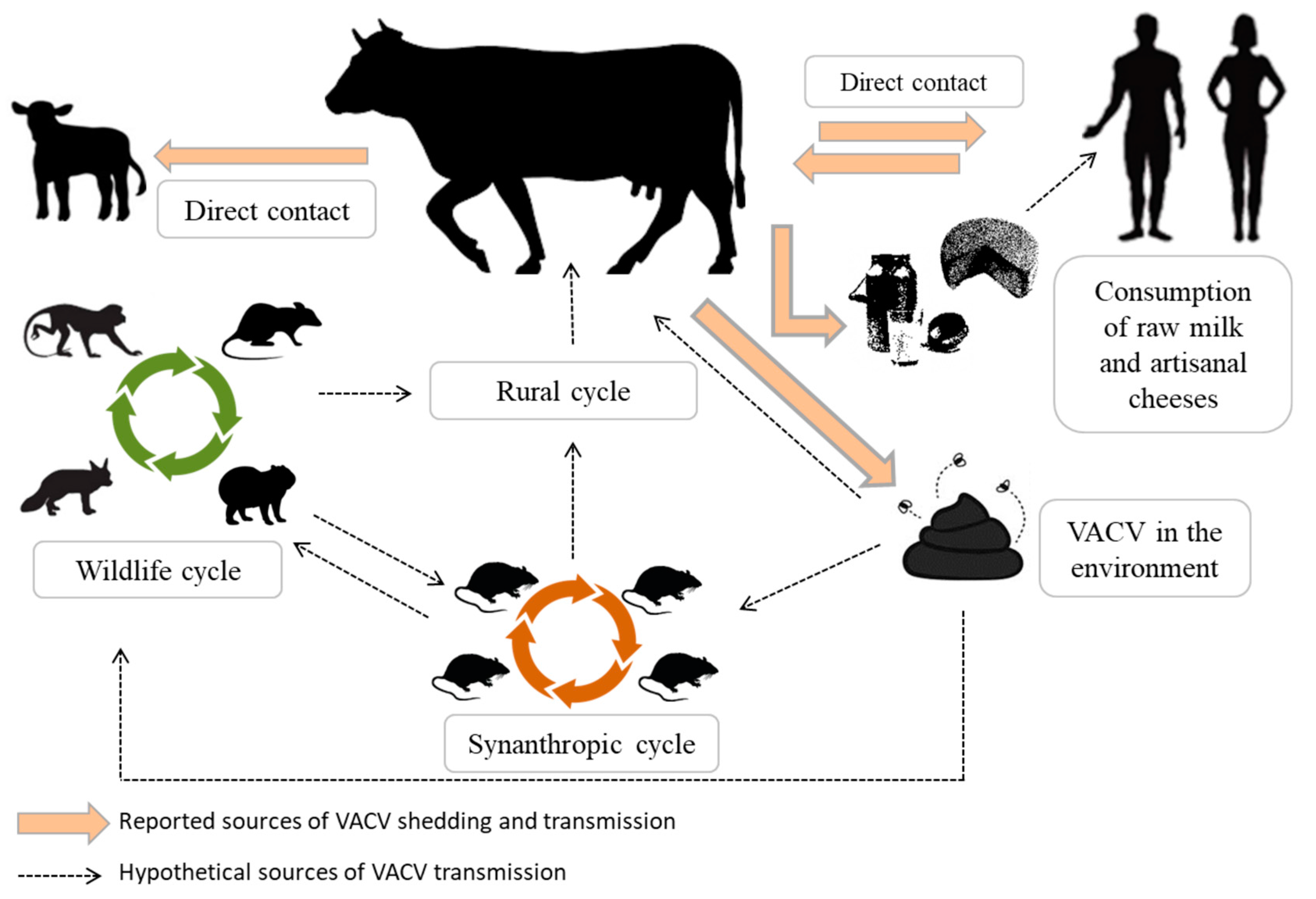
3. Bovine Vaccinia: Viral Shedding Routes and Insights into Its Epidemiology
4. Vaccinia virus: A Possible Foodborne Pathogen
5. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- International Committee on Taxonomy of Viruses (ICTV). International Committee on Taxonomy of Viruses (ICTV). Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 12 October 2017).
- Moss, B. Poxviridae. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 2129–2159. ISBN 9781451105636. [Google Scholar]
- Fenner, F.; Henderson, D.A.; Arita, I.; Jezek, Z.; Ladnyi, I.D. Smallpox and Its Eradication; World Health Organization: Geneva, Switzerland, 1988; ISBN 9241561106. [Google Scholar]
- Henderson, D.A. Principles and lessons from the smallpox eradication programme. Bull. World Health Organ. 1987, 65, 535–546. [Google Scholar] [PubMed]
- Shchelkunov, S.N. An Increasing Danger of Zoonotic Orthopoxvirus Infections. PLoS Pathog. 2013, 9, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Essbauer, S.; Pfeffer, M.; Meyer, H. Zoonotic poxviruses. Vet. Microbiol. 2010, 140, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Kroon, E.G.; Mota, B.E.F.; Abrahão, J.S.; da Fonseca, F.G.; Trindade, G.D.S. Zoonotic Brazilian Vaccinia virus: From field to therapy. Antivir. Res. 2011, 92, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Balamurugan, V.; Bhanuprakash, V.; Venkatesan, G.; Hosamani, M. Emergence and reemergence of vaccinia-like viruses: Global scenario and perspectives. Indian J. Virol. 2012, 23, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bhanuprakash, V.; Venkatesan, G.; Balamurugan, V.; Hosamani, M.; Yogisharadhya, R.; Gandhale, P.; Reddy, K.V.; Damle, A.S.; Kher, H.N.; Chandel, B.S.; et al. Zoonotic infections of buffalopox in India. Zoonoses Public Health 2010, 57. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Hosamani, M.; Balamurugan, V.; Bhanuprakash, V.; Singh, R.K. Partial genetic characterization of viruses isolated from pox-like infection in cattle and buffaloes: Evidence of buffalo pox virus circulation in Indian cows. Arch. Virol. 2010, 155, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.G.; Lanna, M.C.S.; Campos, M.A.S.; Kitajima, E.W.; Peres, J.N.; Golgher, R.R.; Ferreira, P.C.P.; Kroon, E.G. Morphological and molecular characterization of the poxvirus BeAn 58058. Arch. Virol. 1998, 143, 1171–1186. [Google Scholar] [CrossRef] [PubMed]
- Nagasse-Sugahara, T.K.; Kisielius, J.J.; Ueda-Ito, M.; Curti, S.P.; Figueiredo, C.A.; Cruz, Á.S.; Silva, M.M.J.; Ramos, C.H.; Silva, M.C.C.; Sakurai, T.; et al. Human vaccinia-like virus outbreaks in São Paulo and Goiás States, Brazil: Virus detection, isolation and identification. Rev. Inst. Med. Trop. Sao Paulo 2004, 46, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Lobato, Z.I.P.; Trindade, G.S.; Frois, M.C.M.; Ribeiro, E.B.T.; Dias, G.R.C.; Teixeira, B.M.; Lima, F.A.; Almeida, G.M.F.; Kroon, E.G. Outbreak of exantemal disease caused by Vaccinia virus in human and cattle in Zona da Mata region, Minas Gerais. Arq. Bras. Med. Vet. Zootec. 2005, 57, 423–429. [Google Scholar] [CrossRef]
- Damaso, C.R.; Esposito, J.J.; Condit, R.C.; Moussatché, N. An emergent poxvirus from humans and cattle in Rio de Janeiro State: Cantagalo virus may derive from Brazilian smallpox vaccine. Virology 2000, 277, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Schatzmayr, H.G.; Costa, R.V.C.; Gonçalves, M.C.R.; Barreto, D.F.; Batista, V.H.; Silva, M.E.V.; Brust, L.A.C.; Barth, O.M. Infecções humanas causadas por poxvirus relacionados ao vírus vaccinia no Brasil. Rev. Soc. Bras. Med. Trop. 2009, 42, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Trindade, G.S.; da Fonseca, F.G.; Marques, J.T.; Nogueira, M.L.; Mendes, L.C.; Borges, A.S.; Peiro, J.R.; Pituco, E.M.; Bonjardim, C.A.; Ferreira, P.C.; et al. Aracatuba virus: A vaccinialike virus associated with infection in humans and cattle. Emerg. Infect. Dis. 2003, 9, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Megid, J.; Appolinário, C.M.; Langoni, H.; Pituco, E.M.; Okuda, L.H. Short report: Vaccinia virus in humans and cattle in Southwest region of São Paulo State, Brazil. Am. J. Trop. Med. Hyg. 2008, 79, 647–651. [Google Scholar] [PubMed]
- Leite, J.A.; Drumond, B.P.; Trindade, G.S.; Lobato, Z.I.P.; Da Fonseca, F.G.; Dos Santos, J.R.; Madureira, M.C.; Guedes, M.I.M.C.; Ferreira, J.M.S.; Bonjardim, C.A.; et al. Passatempo virus, a Vaccinia virus strain, Brazil. Emerg. Infect. Dis. 2005, 11, 1935–1938. [Google Scholar] [CrossRef] [PubMed]
- Trindade, G.S.; Lobato, Z.I.P.; Drumond, B.P.; Leite, J.A.; Trigueiro, R.C.; Guedes, M.I.M.C.; Da Fonseca, F.G.; Dos Santos, J.R.; Bonjardim, C.A.; Ferreira, P.C.P.; et al. Short report: Isolation of two Vaccinia virus strains from a single bovine vaccinia outbreak in rural area from Brazil: Implications on the emergence of zoonotic Orthopoxviruses. Am. J. Trop. Med. Hyg. 2006, 75, 486–490. [Google Scholar] [PubMed]
- Madureira, M.C. Vaccínia Bovina No Estado De Minas Gerais, 2005–2007. Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil, 2009. [Google Scholar]
- Donatele, D.M.; Travassos, C.E.P.F.; Leite, J.A.; Kroon, E.G. Epidemiologia da poxvirose bovina no Estado do Espírito Santo, Brasil. Braz. J. Vet. Res. Anim. Sci. 2007, 44, 275–282. [Google Scholar] [CrossRef]
- Medaglia, M.L.G.; Pessoa, L.C.G.D.; Sales, E.R.C.; Freitas, T.R.P.; Damaso, C.R. Spread of cantagalo virus to Northern Brazil. Emerg. Infect. Dis. 2009, 15, 1142–1143. [Google Scholar] [CrossRef] [PubMed]
- De Assis, F.L.; Vinhote, W.M.; Barbosa, J.D.; de Oliveira, C.H.S.; de Oliveira, C.M.G.; Campos, K.F.; Silva, N.S.; Trindade, G.D.S.; Abrahão, J.S.; Kroon, E.G. Reemergence of Vaccinia virus during zoonotic outbreak, Pará State, Brazil. Emerg. Infect. Dis. 2013, 19, 2017–2020. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.B.; Assis, F.L.; Ferreira, P.C.P.; Bonjardim, C.A.; de Souza Trindade, G.; Kroon, E.G.; Abrahao, J.S. Group 1 Vaccinia virus Zoonotic Outbreak in Maranhao State, Brazil. Am. J. Trop. Med. Hyg. 2013, 89, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, G.; Assis, F.; Almeida, G.; Albarnaz, J.; Lima, M.; Andrade, A.; Calixto, R.; Oliveira, C.; Diomedes Neto, J.; Trindade, G.; et al. From Lesions to Viral Clones: Biological and Molecular Diversity amongst Autochthonous Brazilian Vaccinia virus. Viruses 2015, 7, 1218–1237. [Google Scholar] [CrossRef] [PubMed]
- Trindade, G.S.; Emerson, G.L.; Carroll, D.S.; Kroon, E.G.; Damon, I.K. Brazilian Vaccinia viruses and their origins. Emerg. Infect. Dis. 2007, 13, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Trindade, G.; Emerson, G.; Sammons, S.; Frace, M.; Govil, D.; Fernandes Mota, B.; Abrahão, J.; de Assis, F.; Olsen-Rasmussen, M.; Goldsmith, C.; et al. Serro 2 Virus Highlights the Fundamental Genomic and Biological Features of a Natural Vaccinia virus Infecting Humans. Viruses 2016, 8, 328. [Google Scholar] [CrossRef] [PubMed]
- Medaglia, M.L.G.; Moussatché, N.; Nitsche, A.; Dabrowski, P.W.; Li, Y.; Damon, I.K.; Lucas, C.G.O.; Arruda, L.B.; Damaso, C.R. Genomic Analysis, Phenotype, and Virulence of the Historical Brazilian Smallpox Vaccine Strain IOC: Implications for the Origins and Evolutionary Relationships of Vaccinia virus. J. Virol. 2015, 89, 11909–11925. [Google Scholar] [CrossRef] [PubMed]
- Damaso, C.R. Revisiting Jenner’s mysteries, the role of the Beaugency lymph in the evolutionary path of ancient smallpox vaccines. Lancet Infect. Dis. 2017, 18, e55–e63. [Google Scholar] [CrossRef]
- Drumond, B.P.; Leite, J.A.; da Fonseca, F.G.; Bonjardim, C.A.; Ferreira, P.C.P.; Kroon, E.G. Brazilian Vaccinia virus strains are genetically divergent and differ from the Lister vaccine strain. Microbes Infect. 2008, 10, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.M.S.; Drumond, B.P.; Guedes, M.I.M.C.; Pascoal-Xavier, M.A.; Almeida-Leite, C.M.; Arantes, R.M.E.; Mota, B.E.F.; Abrahão, J.S.; Alves, P.A.; Oliveira, F.M.; et al. Virulence in murine model shows the existence of two distinct populations of Brazilian Vaccinia virus strains. PLoS ONE 2008, 3, e3043. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.K.; Brum, M.C.S.; Nogueira, C.E.W.; Drumond, B.P.; Alves, P.A.; Siqueira-Lima, L.; Assis, F.L.; Trindade, G.S.; Bonjardim, C.A.; Ferreira, P.C.; et al. Assessing the variability of Brazilian Vaccinia virus isolates from a horse exanthematic lesion: Coinfection with distinct viruses. Arch. Virol. 2011, 156, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Cargnelutti, J.F.; Schmidt, C.; Masuda, E.K.; Braum, L.D.; Weiblen, R.; Furtado Flores, E. Vaccinia viruses isolated from cutaneous disease in horses are highly virulent for rabbits. Microb. Pathog. 2012, 52, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.M.S.; Abrahão, J.S.; Drumond, B.P.; Oliveira, F.M.; Alves, P.A.; Pascoal-Xavier, M.A.; Lobato, Z.I.P.; Bonjardim, C.A.; Peregrino Ferreira, P.C.; Kroon, E.G. Vaccinia virus: Shedding and horizontal transmission in a murine model. J. Gen. Virol. 2008, 89, 2986–2991. [Google Scholar] [CrossRef] [PubMed]
- Usme-Ciro, J.A.; Paredes, A.; Walteros, D.M.; Tolosa-Pérez, E.N.; Laiton-Donato, K.; Pinzón, M.D.; Petersen, B.W.; Gallardo-Romero, N.F.; Li, Y.; Wilkins, K.; et al. Detection and Molecular Characterization of Zoonotic Poxviruses Circulating in the Amazon Region of Colombia, 2014. Emerg. Infect. Dis. 2017, 23, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Franco-Luiz, A.P.M.; Fagundes-Pereira, A.; Costa, G.B.; Alves, P.A.; Oliveira, D.B.; Bonjardim, C.A.; Ferreira, P.C.P.; de Souza Trindade, G.; Panei, C.J.; Galosi, C.M.; et al. Spread of Vaccinia virus to Cattle Herds, Argentina, 2011. Emerg. Infect. Dis. 2014, 20, 1576–1578. [Google Scholar] [CrossRef] [PubMed]
- Franco-Luiz, A.P.M.; Oliveira, D.B.; Pereira, A.F.; Gasparini, M.C.S.; Bonjardim, C.A.; Ferreira, P.C.P.; Trindade, G.D.S.; Puentes, R.; Furtado, A.; Abrahão, J.S.; et al. Detection of Vaccinia virus in Dairy Cattle Serum Samples from 2009, Uruguay. Emerg. Infect. Dis. 2016, 22, 2174–2177. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of United Nations (FAO). World Livestock 2011: Livestock in Food Security World; FAO: Rome, Italy, 2011; ISBN 9789251070130. [Google Scholar]
- Silva-Fernandes, A.T.; Travassos, C.E.P.F.; Ferreira, J.M.S.; Abrahão, J.S.; Rocha, E.S.D.O.; Viana-Ferreira, F.; dos Santos, J.R.; Bonjardim, C.A.; Ferreira, P.C.P.; Kroon, E.G. Natural human infections with Vaccinia virus during bovine vaccinia outbreaks. J. Clin. Virol. 2009, 44, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Schatzmayr, H.G.; Costa, R.V.; Gonçalves, M.C.; D'Andréa, P.S.; Barth, O.M. Human and animal infections by vaccinia-like viruses in the state of Rio de Janeiro: A novel expanding zoonosis. Vaccine 2011, 29. [Google Scholar] [CrossRef] [PubMed]
- Megid, J.; Borges, I.A.; Abrahão, J.S.; Trindade, G.S.; Appolinário, C.M.; Ribeiro, M.G.; Allendorf, S.D.; Antunes, J.M.A.P.; Silva-Fernandes, A.T.; Kroon, E.G. Vaccinia virus Zoonotic Infection, São Paulo State, Brazil. Emerg. Infect. Dis. 2011, 18, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Batista, V.H.; Scremin, J.; Aguiar, L.M.; Schatzmayr, H.G. Vulvar infection and possible human-to-human transmission of bovine poxvirus disease. Virus Rev. Res. 2009, 14, 30–49. [Google Scholar] [CrossRef]
- Assis, F.L.; Borges, I.A.; Mesquita, V.S.; Ferreira, P.C.; Trindade, G.S.; Kroon, E.G.; Abrahão, J.S. Vaccinia virus in household environment during bovine vaccinia outbreak, Brazil. Emerg. Infect. Dis. 2013, 19, 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Assis, F.L.; Franco-Luiz, A.P.M.; Paim, L.M.; Oliveira, G.P.; Pereira, A.F.; de Almeida, G.M.F.; Figueiredo, L.B.; Tanus, A.; Trindade, G.S.; Ferreira, P.P.; et al. Horizontal study of Vaccinia virus infections in an endemic area: Epidemiologic, phylogenetic and economic aspects. Arch. Virol. 2015, 160, 2703–2708. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.C.; Reis, B.B.; Ricci, J.E.R.; Fernandes, F.S.; Corrêa, J.F.; Schatzmayr, H.G. Infecção em humanos por varíola bovina na microrregião de Itajubá, Estado de Minas Gerais: Relato de caso. Rev. Soc. Bras. Med. Trop. 2008, 41, 507–511. [Google Scholar] [CrossRef]
- Rehfeld, I.S.; Guedes, M.I.M.C.; Matos, A.C.D.; de Oliveira, T.M.L.; Rivetti Junior, A.V.; Moura, A.C.J.; Paes, P.R.O.; do Lago, L.A.; Kroon, E.G.; Lobato, Z.I.P. Clinical, hematological and biochemical parameters of dairy cows experimentally infected with Vaccinia virus. Res. Vet. Sci. 2013, 95, 752–757. [Google Scholar] [CrossRef] [PubMed]
- Guedes, M.I.M.C.; Rehfeld, I.S.; De Oliveira, T.M.L.; Assis, F.L.; Matos, A.C.D.; Abrahão, J.S.; Kroon, E.G.; Lobato, Z.I.P. Detection of Vaccinia virus in Blood and Faeces of Experimentally Infected Cows. Transbound. Emerg. Dis. 2012, 60, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Lauder, I.M.; Martin, W.B.; Murray, M.; Pirie, H.M. Experimental vaccinia infection of cattle: A comparison with other virus infections of cows’ teats. Vet. Rec. 1971, 89, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Rivetti, A.V.; Guedes, M.I.M.C.; Rehfeld, I.S.; Oliveira, T.M.L.; Matos, A.C.D.; Abrahão, J.S.; Kroon, E.G.; Lobato, Z.I.P. Bovine vaccinia, a systemic infection: Evidence of fecal shedding, viremia and detection in lymphoid organs. Vet. Microbiol. 2013, 162. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, I.S.; Guedes, M.I.M.C.; da Silva, T.M.R.; Matos, A.C.D.; Campos, F.S.; Aline, M.; Guedes, R.M.C.; Lobato, Z.I.P. Pathogenesis of Vaccinia virus in experimentally infected cows. PLoS ONE. under review.
- Abrahão, J.S.; Oliveira, T.M.L.; Campos, R.K.; Madureira, M.C.; Kroon, E.G.; Lobato, Z.I.P. Bovine vaccinia outbreaks: Detection and isolation of Vaccinia virus in milk samples. Foodborne Pathog. Dis. 2009, 6, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.M.L.; Guedes, M.I.M.C.; Rehfeld, I.S.; Matos, A.C.D.; Rivetti, A.V.; Alves, P.A.; Galinari, G.C.F.; Cerqueira, M.M.O.P.; Abrahão, J.S.; Lobato, Z.I.P. Detection of Vaccinia Virus in Milk: Evidence of a Systemic and Persistent Infection in Experimentally Infected Cows. Foodborne Pathog. Dis. 2015, 12, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Panchanathan, V.; Chaudhri, G.; Karupiah, G. Correlates of protective immunity in poxvirus infection: Where does antibody stand? Immunol. Cell Biol. 2008, 86, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Spriggs, M.K.; Kollert, B.H.; Satot, T.; Morrisseyt, P.J.; Fanslowt, W.C.; Smithiest, O.; Voices, R.F.; Widmert, M.B.; Maliszewski, C.R. b2-Microglobulin-, CD8+ T-cell-deficient mice survive inoculation with high doses of Vaccinia virus and exhibit altered IgG responses. Immunology 1992, 89, 6070–6074. [Google Scholar]
- Coulibaly, S.; Brühl, P.; Mayrhofer, J.; Schmid, K.; Gerencer, M.; Falkner, F.G. The nonreplicating smallpox candidate vaccines defective vaccinia Lister (dVV-L) and modified vaccinia Ankara (MVA) elicit robust long-term protection. Virology 2005, 341, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Johnson, A.; Liggitt, D.; Bevan, M.J. Cellular and humoral immunity against Vaccinia virus infection of mice. J. Immunol. 2004, 172, 6265–6271. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.C.D. Vaccinia virus: Padronização de Técnica Para Imunodiagnóstico e Estudo da Resposta Imune Humoral de Bovinos. Master’s Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil, 2012. [Google Scholar]
- Guedes, M.I.M.C.; Villani, F.N.A.; Gerber, P.F.; Bozzi, A.M.; Matos, A.C.D.; Rehfeld, I.S.; Damasio, M.P.S.; Dorneles, E.M.S.; Carvalho, A.T.; Côrrea-Oliveira, R.; et al. Cellular immune response in cows experimentally infected with Vaccinia virus. Virus Rev. Res. 2012, 17, 398–399. [Google Scholar] [CrossRef]
- Rehfeld, I.S.; Matos, A.C.D.; Guedes, M.I.M.C.; Costa, A.G.; Fraiha, A.L.S.; Lobato, Z.I.P. Subclinical bovine vaccinia: An important risk factor in the epidemiology of this zoonosis in cattle. Res. Vet. Sci. 2017, 114, 233–235. [Google Scholar] [CrossRef] [PubMed]
- Quixabeira-Santos, J.C.; Medaglia, M.L.G.; Pescador, C.A.; Damaso, C.R. Animal movement and establishment of Vaccinia virus Cantagalo strain in Amazon biome, Brazil. Emerg. Infect. Dis. 2011, 17, 726. [Google Scholar] [CrossRef] [PubMed]
- Abrahão, J.S.; de Souza Trindade, G.; Ferreira, J.M.S.; Campos, R.K.; Bonjardim, C.A.; Ferreira, P.C.P.; Kroon, E.G. Long-lasting stability of Vaccinia virus strains in murine feces: Implications for virus circulation and environmental maintenance. Arch. Virol. 2009, 154, 1551–1553. [Google Scholar] [CrossRef] [PubMed]
- D’Anunciação, L.; Guedes, M.I.M.; Oliveira, T.L.; Rehfeld, I.; Bonjardim, C.A.; Ferreira, P.P.; de Souza Trindade, G.; Lobato, Z.P.; Kroon, E.G.; Abrahão, J.S. Filling One More Gap: Experimental Evidence of Horizontal Transmission of Vaccinia virus Between Bovines and Rodents. Vector-Borne Zoonotic Dis. 2012, 12, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, I.S.; Guedes, M.I.M.C.; Fraiha, A.L.S.; Costa, A.G.; Matos, A.C.D.; Fiúza, A.T.L.; Lobato, Z.I.P. Vaccinia virus transmission through experimentally contaminated milk using a murine model. PLoS ONE 2015, 10, e127350. [Google Scholar] [CrossRef] [PubMed]
- Abrahão, J.S.; Guedes, M.I.M.; Trindade, G.S.; Fonseca, F.G.; Campos, R.K.; Mota, B.F.; Lobato, Z.I.P.; Silva-Fernandes, A.T.; Rodrigues, G.O.L.; Lima, L.S.; et al. One more piece in the VACV ecological puzzle: Could peridomestic rodents be the link between wildlife and bovine vaccinia outbreaks in Brazil? PLoS ONE 2009, 4, e7428. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.V.; Medaglia, M.L.G.; Soares, H.S.; Quixabeira-Santos, J.C.; Gennari, S.M.; Damaso, C.R. Presence of neutralizing antibodies to Orthopoxvirus in Capybaras (Hydrochoerus hydrochaeris) in Brazil. J. Infect. Dev. Ctries. 2014, 8, 1646–1649. [Google Scholar] [CrossRef] [PubMed]
- Peres, M.G.; Bacchiega, T.S.; Appolinario, C.M.; Vicente, A.F.; Allendorf, S.D.; Antunes, J.M.A.P.; Moreira, S.A.; Legatti, E.; Fonseca, C.R.; Pituco, E.M.; et al. Serological study of Vaccinia virus reservoirs in areas with and without official reports of outbreaks in cattle and humans in Sao Paulo, Brazil. Arch. Virol. 2013, 158, 2433–2441. [Google Scholar] [CrossRef] [PubMed]
- Abrahão, J.S.; Silva-Fernandes, A.T.; Lima, L.S.; Campos, R.K.; Guedes, M.I.M.C.; Cota, M.M.G.; Assis, F.L.; Borges, I.A.; Souza-Júnior, M.F.; Lobato, Z.I.P.; et al. Vaccinia virus Infection in Monkeys, Brazilian Amazon. Emerg. Infect. Dis. 2010, 16, 976–979. [Google Scholar] [CrossRef] [PubMed]
- Peres, M.G.; Barros, C.B.; Appolinario, C.M.; Antunes, J.M.A.P.; Mioni, M.S.R.; Bacchiega, T.S.; Allendorf, S.D.; Vicente, A.F.; Fonseca, C.R.; Megid, J. Dogs and Opossums Positive for Vaccinia virus during Outbreak Affecting Cattle and Humans, Sao Paulo State, Brazil. Emerg. Infect. Dis. 2016, 22, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.B.; Miranda, J.B.; Almeida, G.G.; Silva de Oliveira, J.; Pinheiro, M.S.; Goncalves, S.A.; Pimenta Dos Reis, J.K.; Goncalves, R.; Ferreira, P.C.P.; Bonjardim, C.A.; et al. Detection of Vaccinia virus in Urban Domestic Cats, Brazil. Emerg. Infect. Dis. 2017, 23, 360–362. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.B.; Borges, I.A.; Campos, S.P.S.; Vieira, F.N.; de Azara, T.M.F.; Marques, F.A.; Costa, G.B.; Luis, A.P.M.F.; de Oliveira, J.S.; Ferreira, P.C.P.; et al. Serologic and Molecular Evidence of Vaccinia virus Circulation among Small Mammals from Different Biomes, Brazil. Emerg. Infect. Dis. 2017, 23, 931–938. [Google Scholar] [CrossRef] [PubMed]
- Dutra, L.A.L.; de Freitas Almeida, G.M.; Oliveira, G.P.; Abrahao, J.S.; Kroon, E.G.; de Souza Trindade, G. Molecular evidence of Orthopoxvirus DNA in capybara (Hydrochoerus hydrochaeris) stool samples. Arch. Virol. 2017, 162, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Essbauer, S.; Meyer, H.; Porsch-Ozcurumez, M.; Pfeffer, M. Long-lasting stability of Vaccinia virus (Orthopoxvirus) in food and environmental samples. Zoonoses Public Health 2007, 54, 118–124. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.M.L.; Rehfeld, I.S.; Guedes, M.I.M.C.; Ferreira, J.M.S.; Kroon, E.G.; Lobato, Z.I.P. Susceptibility of Vaccinia virus to chemical disinfectants. Am. J. Trop. Med. Hyg. 2011, 85, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.C.D.; Guedes, M.I.M.C.; Rehfeld, I.S.; Costa, E.A.; Costa, A.G.; Silva, N.L.D.; Lage, A.P.; Lobato, Z.I.P. Bovine vaccinia: Inactivated Vaccinia virus vaccine induces protection in murine model. Vet. Microbiol. 2017, 204, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Matos, A.C.D. Development, potency and efficacy test of an inactivated vaccine using the Vaccinia virus strain GP2. Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil, 2016. [Google Scholar]
- De Oliveira, T.M.L.; Rehfeld, I.S.; Siqueira, J.M.F.; Abrahão, J.S.; Campos, R.K.; dos Santos, A.K.R.; Cerqueira, M.M.O.P.; Kroon, E.G.; Lobato, Z.I.P. Vaccinia virus is not inactivated after thermal treatment and cheese production using experimentally contaminated milk. Foodborne Pathog. Dis. 2010, 7, 1491–1496. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.M.L.; Guedes, M.I.M.C.; Rehfeld, I.S.; Matos, A.C.D.; Rivetti Júnior, A.V.; da Cunha, A.F.; Cerqueira, M.M.O.P.; Abrahão, J.S.; Lobato, Z.I.P. Vaccinia virus detection in dairy products made with milk from experimentally infected cows. Transbound. Emerg. Dis. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, I.S.; Fraiha, A.L.S.; Matos, A.C.D.; Guedes, M.I.M.C.; Costa, E.A.; de Souza, M.R.; Cavalcante, L.F.L.; Lobato, Z.I.P. Short communication: Survival of Vaccinia virus in inoculated cheeses during 60-day ripening. J. Dairy Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rehfeld, I.S. Transmissão de Vaccinia virus pelo leite em modelo murino; detecção e viabilidade de poxvirus no queijo e leite. Ph.D. Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Minas Gerais, Brazil, 2016. [Google Scholar]
- Gurav, Y.K.; Raut, C.G.; Yadav, P.D.; Tandale, B.V.; Sivaram, A.; Pore, M.D.; Basu, A.; Mourya, D.T.; Mishra, A.C. Buffalopox outbreak in humans and animals in Western Maharashtra, India. Prev. Vet. Med. 2011, 100, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.B.; Teodoro, L.; Augusto, S.; Leite, J.A.; César, P.; Ferreira, P.; Bonjardim, C.A.; Abrahão, J.S.; Kroon, E.G.; Moreno, E.C. Seroprevalence of Orthopoxvirus in rural Brazil : Insights into anti-OPV immunity status and its implications for emergent zoonotic OPV. Virol. J. 2016, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Instituto Brasileiro de Geografia e Estatística (IBGE) Produção da Pecuária Municipal—PPM. Available online: https://www.ibge.gov.br/estatisticas-novoportal/economicas/agricultura-e-pecuaria/9107-producao-da-pecuaria-municipal.html?&t=destaques (accessed on 15 December 2017).


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matos, A.C.D.; Rehfeld, I.S.; Guedes, M.I.M.C.; Lobato, Z.I.P. Bovine Vaccinia: Insights into the Disease in Cattle. Viruses 2018, 10, 120. https://doi.org/10.3390/v10030120
Matos ACD, Rehfeld IS, Guedes MIMC, Lobato ZIP. Bovine Vaccinia: Insights into the Disease in Cattle. Viruses. 2018; 10(3):120. https://doi.org/10.3390/v10030120
Chicago/Turabian StyleMatos, Ana Carolina Diniz, Izabelle Silva Rehfeld, Maria Isabel Maldonado Coelho Guedes, and Zélia Inês Portela Lobato. 2018. "Bovine Vaccinia: Insights into the Disease in Cattle" Viruses 10, no. 3: 120. https://doi.org/10.3390/v10030120
APA StyleMatos, A. C. D., Rehfeld, I. S., Guedes, M. I. M. C., & Lobato, Z. I. P. (2018). Bovine Vaccinia: Insights into the Disease in Cattle. Viruses, 10(3), 120. https://doi.org/10.3390/v10030120