Vitamin E Supplementation Ameliorates Newcastle Disease Virus-Induced Oxidative Stress and Alleviates Tissue Damage in the Brains of Chickens

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Reagents and Viruses
2.2. Ethics Statement
2.3. Animal Grouping and Treatments
2.4. Sample Collection
2.5. Preparation of Tissue Homogenates
2.6. Determination of Protein Content
2.7. Determination of NO Level
2.8. Determination of MDA Level
2.9. Determination of GSH Level
2.10. Determination of TAOC
2.11. Determination of CAT Activity
2.12. Determination of SOD activity
2.13. Determination of GST Activity
2.14. NDV Antibody Titers
2.15. Detection of Viral Loads in the Brain
2.16. Histopathology
2.17. Statistical Analysis
3. Results
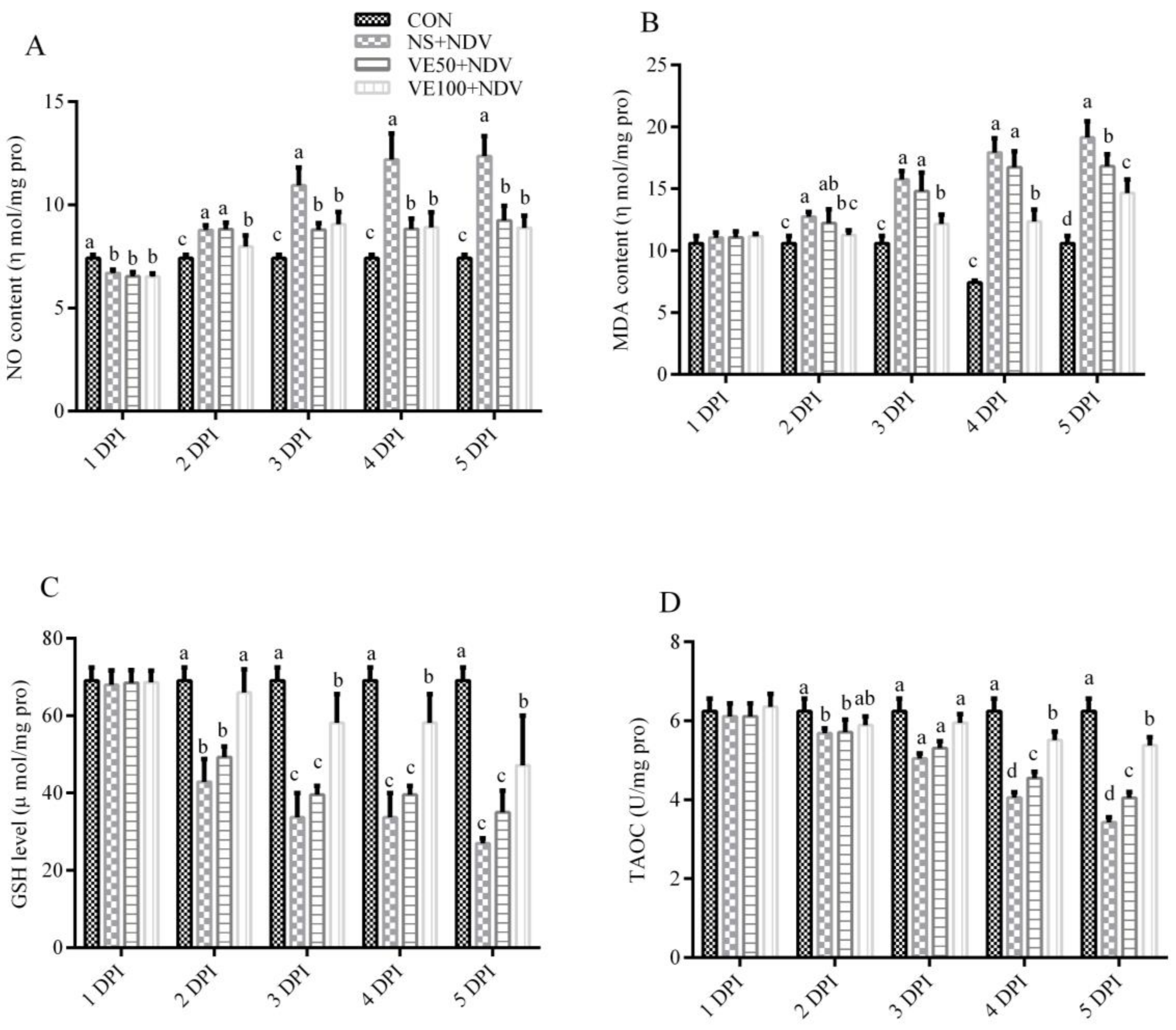
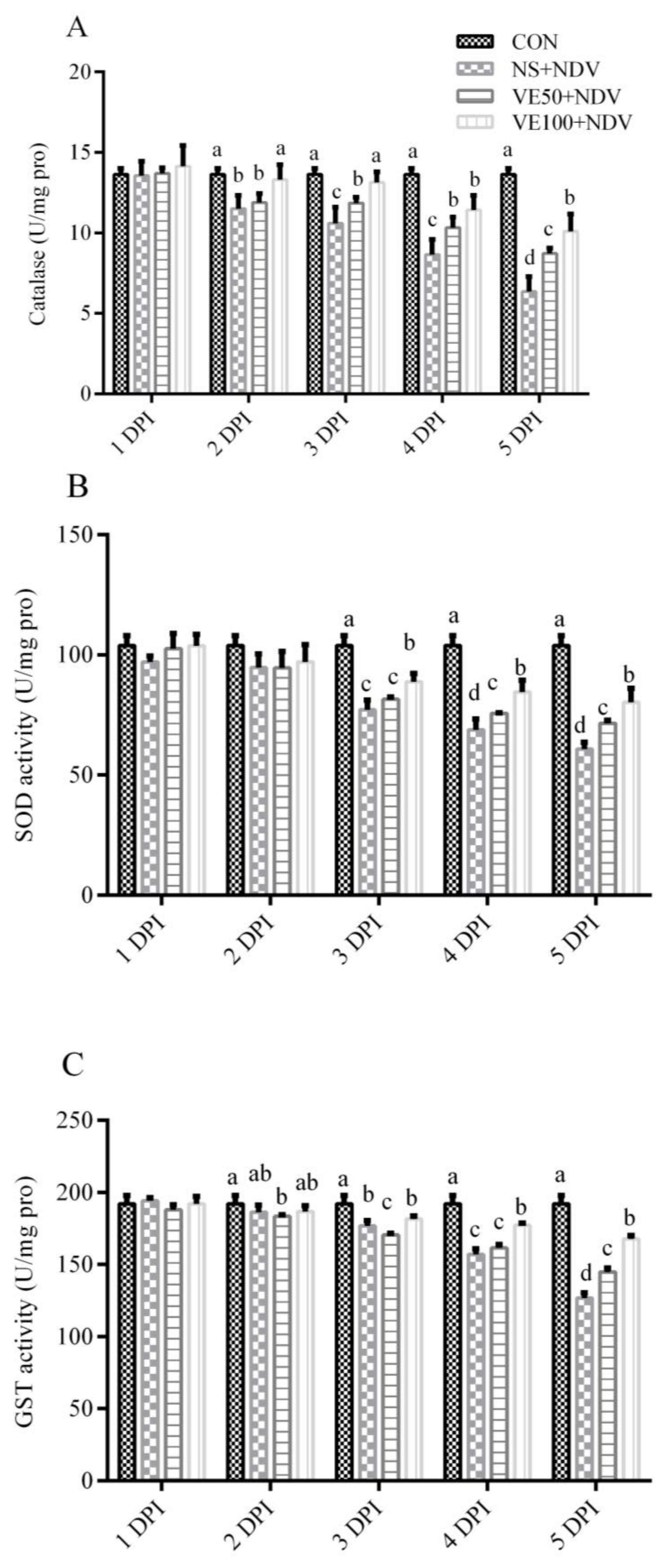
3.1. Antioxidants and Associated Enzymes
3.1.1. Nonenzymatic Antioxidant Parameters
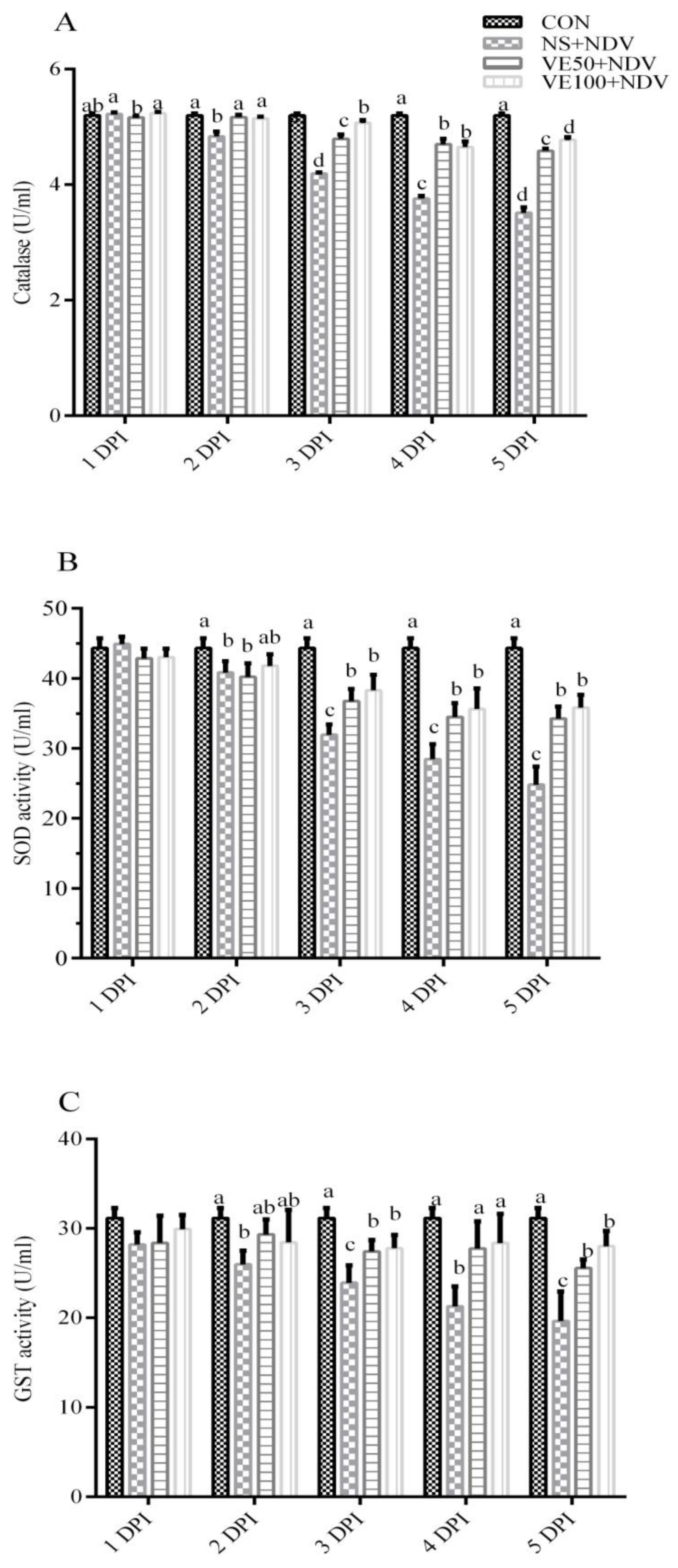
3.1.2. Enzymatic Antioxidant Parameters
3.2. Virus Load and Antibody Titers
3.3. Brain Histopathology
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-κB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New roles in redox signaling for an old antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative stress during HIV infection: Mechanisms and consequences. Oxid. Med. Cell. Longev. 2016, 2016, 8910396. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhang, H.; Wang, Q.; Wang, J. The NS1 protein of avian influenza virus H9N2 induces oxidative-stress-mediated chicken oviduct epithelial cells apoptosis. J. Gen. Virol. 2016, 97, 3183–3192. [Google Scholar] [CrossRef] [PubMed]
- Hosakote, Y.M.; Brasier, A.R.; Casola, A.; Garofalo, R.P.; Kurosky, A.; Lyles, D.S. Respiratory syncytial virus infection triggers epithelial HMGB1 release as a damage-associated molecular pattern promoting a monocytic inflammatory response. J. Virol. 2016, 90, 9618–9631. [Google Scholar] [CrossRef] [PubMed]
- Camini, F.C.; da Silva Caetano, C.C.; Almeida, L.T.; de Brito Magalhaes, C.L. Implications of oxidative stress on viral pathogenesis. Arch. Virol. 2017, 162, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Cemek, M.; Dede, S.; Bayiroglu, F.; Caksen, H.; Cemek, F.; Mert, N. Oxidant and non-enzymatic antioxidant status in measles. J. Trop. Pediatr. 2007, 53, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.W.; Su, Y.C.; Her, G.M.; Ken, C.F.; Hong, J.R. Betanodavirus induces oxidative stress-mediated cell death that prevented by anti-oxidants and zfcatalase in fish cells. PLoS ONE 2011, 6, e25853. [Google Scholar] [CrossRef] [PubMed]
- Ciriolo, M.R.; Palamara, A.T.; Incerpi, S.; Lafavia, E.; Bue, M.C.; DeVito, P.; Garaci, E.; Rotilio, G. Loss of GSH, oxidative stress, and decrease of intracellular pH as sequential steps in viral infection. J. Biol. Chem. 1997, 272, 2700–2708. [Google Scholar] [CrossRef] [PubMed]
- Escaffre, O.; Halliday, H.; Borisevich, V.; Casola, A.; Rockx, B. Oxidative stress in NIPAH virus-infected human small airway epithelial cells. J. Gen. Virol. 2015, 96, 2961–2970. [Google Scholar] [CrossRef] [PubMed]
- Subbaiah, K.C.; Raniprameela, D.; Visweswari, G.; Rajendra, W.; Lokanatha, V. Perturbations in the antioxidant metabolism during newcastle disease virus (NDV) infection in chicken: Protective role of vitamin E. Naturwissenschaften 2011, 98, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Subbaiah, K.C.; Valluru, L.; Rajendra, W.; Ramamurthy, C.; Thirunavukkarusu, C.; Subramanyam, R. Newcastle disease virus (NDV) induces protein oxidation and nitration in brain and liver of chicken: Ameliorative effect of vitamin E. Int. J. Biochem. Cell Biol. 2015, 64, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Cattoli, G.; Susta, L.; Terregino, C.; Brown, C. Newcastle disease: A review of field recognition and current methods of laboratory detection. J. Vet. Diagn. Investig. 2011, 23, 637–656. [Google Scholar] [CrossRef] [PubMed]
- Moura, V.M.; Susta, L.; Cardenas-Garcia, S.; Stanton, J.B.; Miller, P.J.; Afonso, C.L.; Brown, C.C. Neuropathogenic capacity of lentogenic, mesogenic, and velogenic newcastle disease virus strains in day-old chickens. Vet. Pathol. 2016, 53, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.; Koch, G.; Suarez, D.; Mundt, E.; Jones, R.; Rautenschlein, S. Newcastle disease, other paramyxoviruses, and avian metapneumovirus infections. In Diseases of Poultry, 13th ed.; Blackwell Publishing: Ames, IA, USA, 2013; pp. 250–377. [Google Scholar]
- Ecco, R.; Susta, L.; Afonso, C.L.; Miller, P.J.; Brown, C. Neurological lesions in chickens experimentally infected with virulent newcastle disease virus isolates. Avian Pathol. 2011, 40, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Kommers, G.D.; King, D.J.; Seal, B.S.; Brown, C.C. Virulence of pigeon-origin newcastle disease virus isolates for domestic chickens. Avian Dis. 2001, 45, 906–921. [Google Scholar] [CrossRef] [PubMed]
- Susta, L.; Miller, P.J.; Afonso, C.L.; Brown, C.C. Clinicopathological characterization in poultry of three strains of newcastle disease virus isolated from recent outbreaks. Vet. Pathol. 2011, 48, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.J.; Afonso, C.L.; El Attrache, J.; Dorsey, K.M.; Courtney, S.C.; Guo, Z.; Kapczynski, D.R. Effects of newcastle disease virus vaccine antibodies on the shedding and transmission of challenge viruses. Dev. Comp. Immunol. 2013, 41, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Ulatowski, L.; Manor, D. Vitamin E trafficking in neurologic health and disease. Annu. Rev. Nutr. 2013, 33, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, P.A.; Beck, M.A. The immune response to herpes simplex virus encephalitis in mice is modulated by dietary vitamin E. J. Nutr. 2008, 138, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Han, S.N.; Wu, D.; Ha, W.K.; Beharka, A.; Smith, D.E.; Bender, B.S.; Meydani, S.N. Vitamin E supplementation increases T helper 1 cytokine production in old mice infected with influenza virus. Immunology 2000, 100, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Chen, L.; Wu, L.; Liu, X. Newcastle disease in geese: Natural occurrence and experimental infection. Avian Pathol. 2004, 33, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Ma, H.; Wu, Y.; Liu, W.; Wang, X.; Liu, Y.; Liu, X. A vaccine candidate of attenuated genotype VII newcastle disease virus generated by reverse genetics. Vaccine 2009, 27, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Cornax, I.; Miller, P.J.; Afonso, C.L. Characterization of live lasota vaccine strain-induced protection in chickens upon early challenge with a virulent newcastle disease virus of heterologous genotype. Avian Dis. 2012, 56, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Liu, Y.; Liu, C.; Cheng, J.; Fan, W.; Zhang, X.; Liu, J. Growth performance and oxidative damage in kidney induced by oral administration of CR(III) in chicken. Chemosphere 2015, 139, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Floyd, R.A. Oxygen radical-mediated damage to brain tissue. In Oxygen Radicals in Biology and Medicine; Simic, M.G., Taylor, K.A., Ward, J.F., von Sonntag, C., Eds.; Springer: Boston, MA, USA, 1988; pp. 1015–1023. [Google Scholar]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef] [PubMed]
- Naziroglu, M. New molecular mechanisms on the activation of TRPM2 channels by oxidative stress and ADP-ribose. Neurochem. Res. 2007, 32, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Zaki, M.H.; Akuta, T.; Akaike, T. Nitric oxide-induced nitrative stress involved in microbial pathogenesis. J. Pharmacol. Sci. 2005, 98, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Akaike, T.; Noguchi, Y.; Ijiri, S.; Setoguchi, K.; Suga, M.; Zheng, Y.M.; Dietzschold, B.; Maeda, H. Pathogenesis of influenza virus-induced pneumonia: Involvement of both nitric oxide and oxygen radicals. Proc. Natl. Acad. Sci. USA 1996, 93, 2448–2453. [Google Scholar] [CrossRef] [PubMed]
- Perrone, L.A.; Belser, J.A.; Wadford, D.A.; Katz, J.M.; Tumpey, T.M. Inducible nitric oxide contributes to viral pathogenesis following highly pathogenic influenza virus infection in mice. J. Infect. Dis. 2013, 207, 1576–1584. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, H.; Takeuchi, R.; Ohsaki, M.; Seki, K.; Chiba, S. Respiratory syncytial virus infection of human respiratory epithelial cells enhances inducible nitric oxide synthase gene expression. J. Leukoc. Biol. 1999, 66, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.A.; Saxena, V.K.; Ara, A.; Singh, K.B.; Sundaresan, N.R.; Saxena, M.; Rasool, T.J. Immune response to newcastle disease virus in chicken lines divergently selected for cutaneous hypersensitivity. Int. J. Immunogenet. 2007, 34, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Sick, C.; Schultz, U.; Munster, U.; Meier, J.; Kaspers, B.; Staeheli, P. Promoter structures and differential responses to viral and nonviral inducers of chicken type i interferon genes. J. Biol. Chem. 1998, 273, 9749–9754. [Google Scholar] [CrossRef] [PubMed]
- Rue, C.A.; Susta, L.; Cornax, I.; Brown, C.C.; Kapczynski, D.R.; Suarez, D.L.; King, D.J.; Miller, P.J.; Afonso, C.L. Virulent newcastle disease virus elicits a strong innate immune response in chickens. J. Gen. Virol. 2011, 92, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Misra, U.K.; Kalita, J.; Khanna, V.K.; Khan, M.Y. Imbalance in oxidant/antioxidant system in different brain regions of rat after the infection of Japanese encephalitis virus. Neurochem. Int. 2009, 55, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Abba, Y.; Hassim, H.; Hamzah, H.; Ibrahim, O.E.; Mohd Lila, M.A.; Noordin, M.M. Pathological vicissitudes and oxidative stress enzyme responses in mice experimentally infected with Reptarenavirus (isolate UPM/MY01). Microb. Pathog. 2017, 104, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Schachtele, S.J.; Hu, S.; Little, M.R.; Lokensgard, J.R. Herpes simplex virus induces neural oxidative damage via microglial cell toll-like receptor-2. J. Neuroinflammation 2010, 7, 35. [Google Scholar] [CrossRef] [PubMed]
- Sultana, R.; Perluigi, M.; Allan Butterfield, D. Lipid peroxidation triggers neurodegeneration: A redox proteomics view into the Alzheimer disease brain. Free Radic. Biol. Med. 2013, 62, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; Chauhan, V.; Chauhan, A. Glutathione redox imbalance in brain disorders. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Escaffre, O.; Saito, T.B.; Juelich, T.L.; Ikegami, T.; Smith, J.K.; Perez, D.D.; Atkins, C.; Levine, C.B.; Huante, M.B.; Nusbaum, R.J.; et al. Contribution of human lung parenchyma and leukocyte influx to oxidative stress and immune system-mediated pathology following nipah virus infection. J. Virol. 2017, 91, e00275-17. [Google Scholar] [CrossRef] [PubMed]
- Matveeva, O.V.; Kochneva, G.V.; Netesov, S.V.; Onikienko, S.B.; Chumakov, P.M. Mechanisms of oncolysis by paramyxovirus sendai. Acta Nat. 2015, 7, 6–16. [Google Scholar]
- Sen, C.K.; Khanna, S.; Roy, S. Tocotrienol: The natural vitamin e to defend the nervous system? Ann. N. Y. Acad Sci. 2004, 1031, 127–142. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rehman, Z.U.; Qiu, X.; Sun, Y.; Liao, Y.; Tan, L.; Song, C.; Yu, S.; Ding, Z.; Munir, M.; Nair, V.; et al. Vitamin E Supplementation Ameliorates Newcastle Disease Virus-Induced Oxidative Stress and Alleviates Tissue Damage in the Brains of Chickens. Viruses 2018, 10, 173. https://doi.org/10.3390/v10040173
Rehman ZU, Qiu X, Sun Y, Liao Y, Tan L, Song C, Yu S, Ding Z, Munir M, Nair V, et al. Vitamin E Supplementation Ameliorates Newcastle Disease Virus-Induced Oxidative Stress and Alleviates Tissue Damage in the Brains of Chickens. Viruses. 2018; 10(4):173. https://doi.org/10.3390/v10040173
Chicago/Turabian StyleRehman, Zaib Ur, Xusheng Qiu, Yingjie Sun, Ying Liao, Lei Tan, Cuiping Song, Shengqing Yu, Zhuang Ding, Muhammad Munir, Venugopal Nair, and et al. 2018. "Vitamin E Supplementation Ameliorates Newcastle Disease Virus-Induced Oxidative Stress and Alleviates Tissue Damage in the Brains of Chickens" Viruses 10, no. 4: 173. https://doi.org/10.3390/v10040173
APA StyleRehman, Z. U., Qiu, X., Sun, Y., Liao, Y., Tan, L., Song, C., Yu, S., Ding, Z., Munir, M., Nair, V., Meng, C., & Ding, C. (2018). Vitamin E Supplementation Ameliorates Newcastle Disease Virus-Induced Oxidative Stress and Alleviates Tissue Damage in the Brains of Chickens. Viruses, 10(4), 173. https://doi.org/10.3390/v10040173