Discovery and Biochemical Characterization of PlyP56, PlyN74, and PlyTB40—Bacillus Specific Endolysins

 , , ,
, , , 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacteriophage Sequence Analysis
2.2. Bacterial Strains and Growth Conditions
2.3. Cloning of Vector Constructs
2.4. Recombinant Protein Expression
2.5. Recombinant Protein Purification
2.6. Turbidity Reduction Assay
2.7. Plate Lysis (Spot) Assay
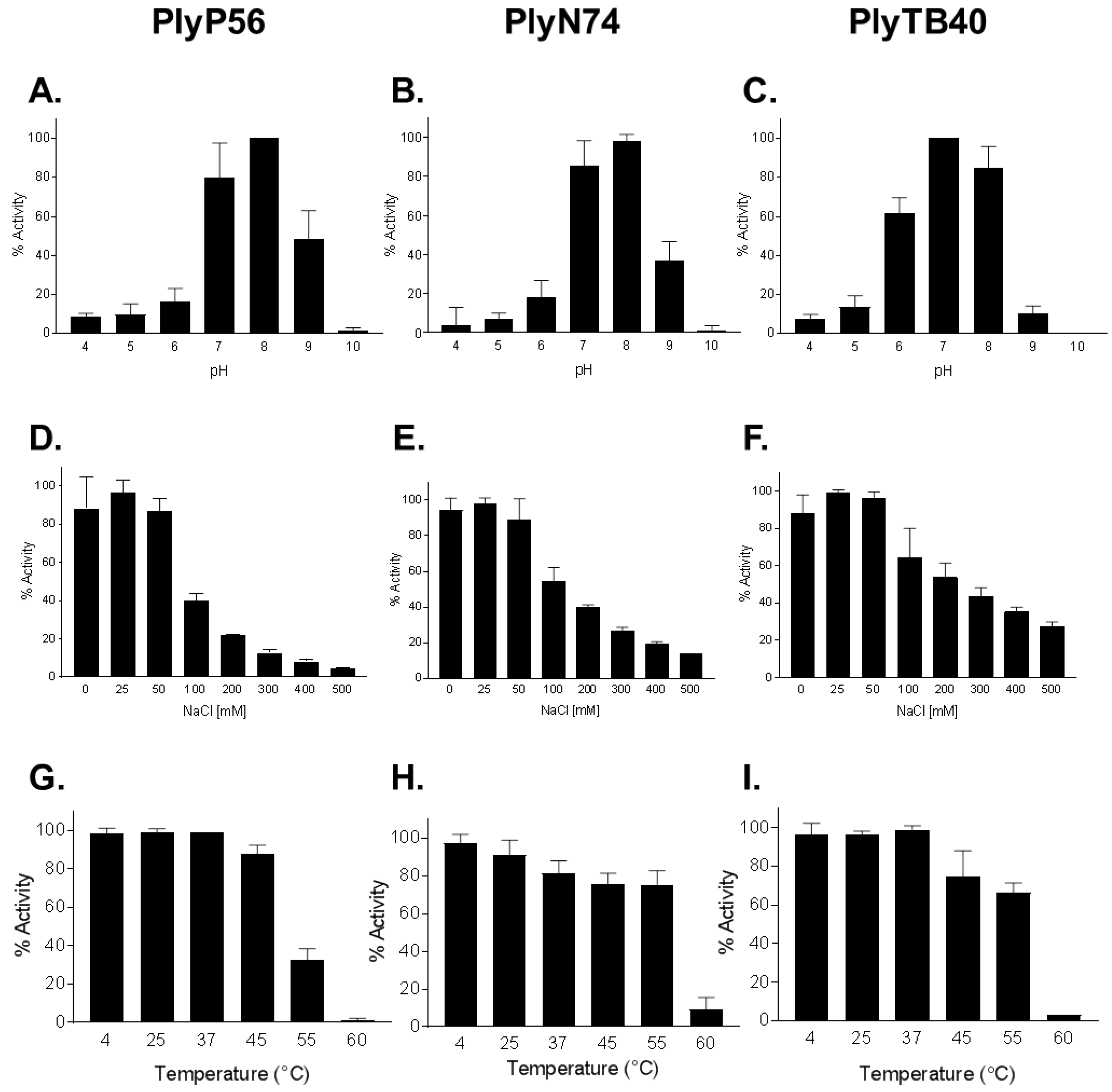
2.8. Characterization of PlyP56, PlyN74, and PlyTB40
2.9. Spectrum of Lytic Activity
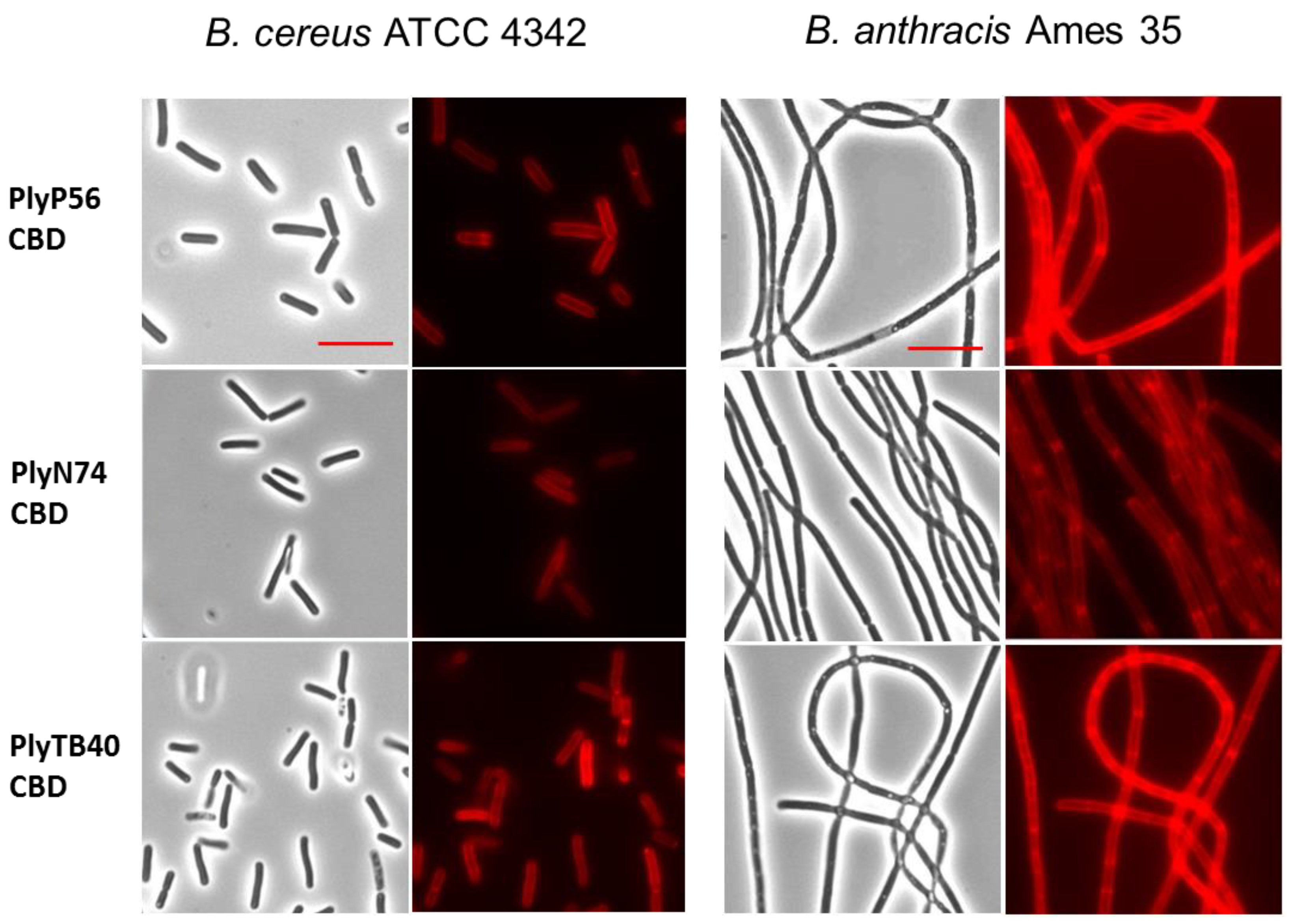
2.10. Fluorescent Labeling of CBDs
2.11. CBD-Binding Assay
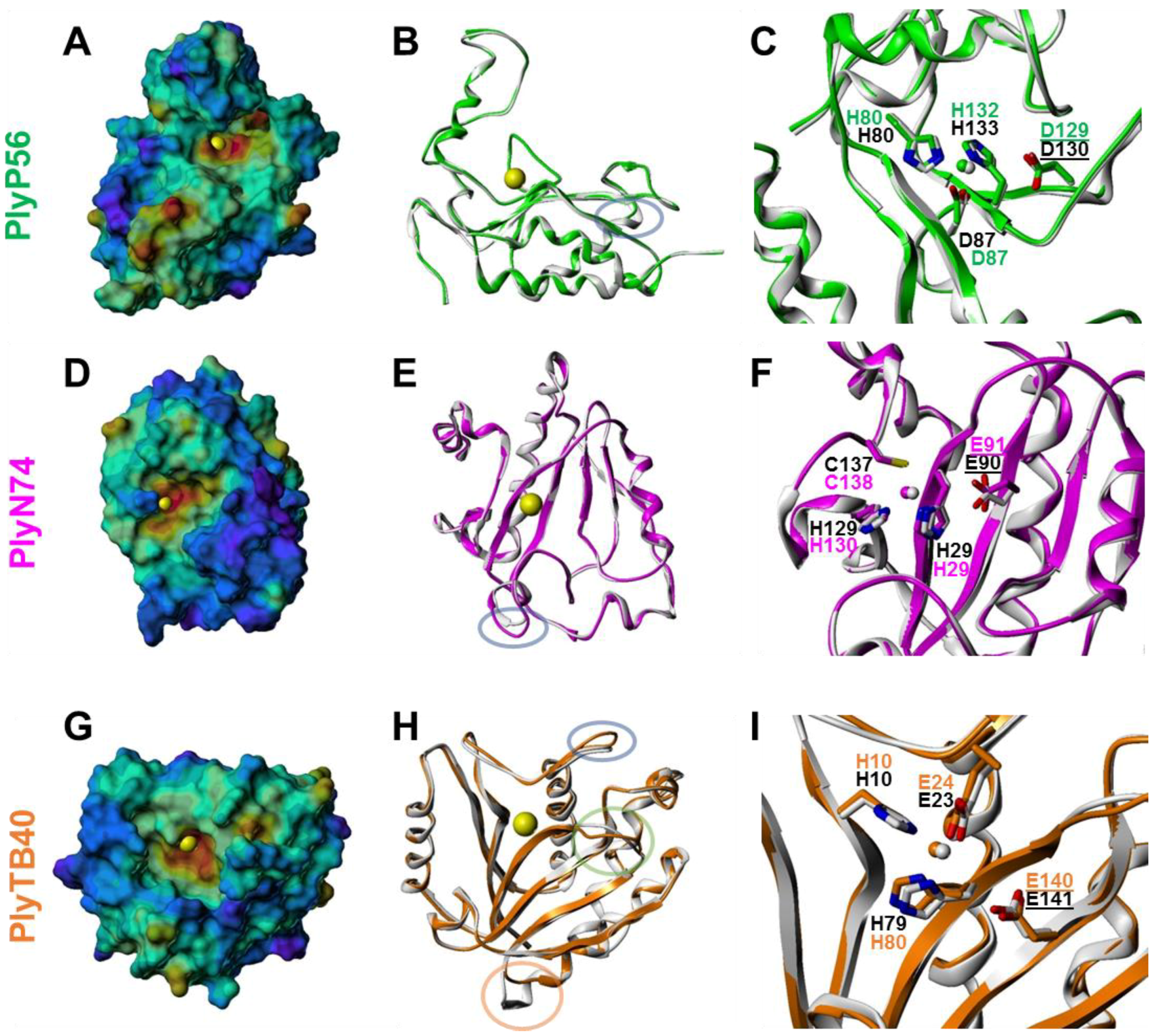
2.12. Structural Modeling of BACILLUS Bacteriophage Endolysin EADs
3. Results
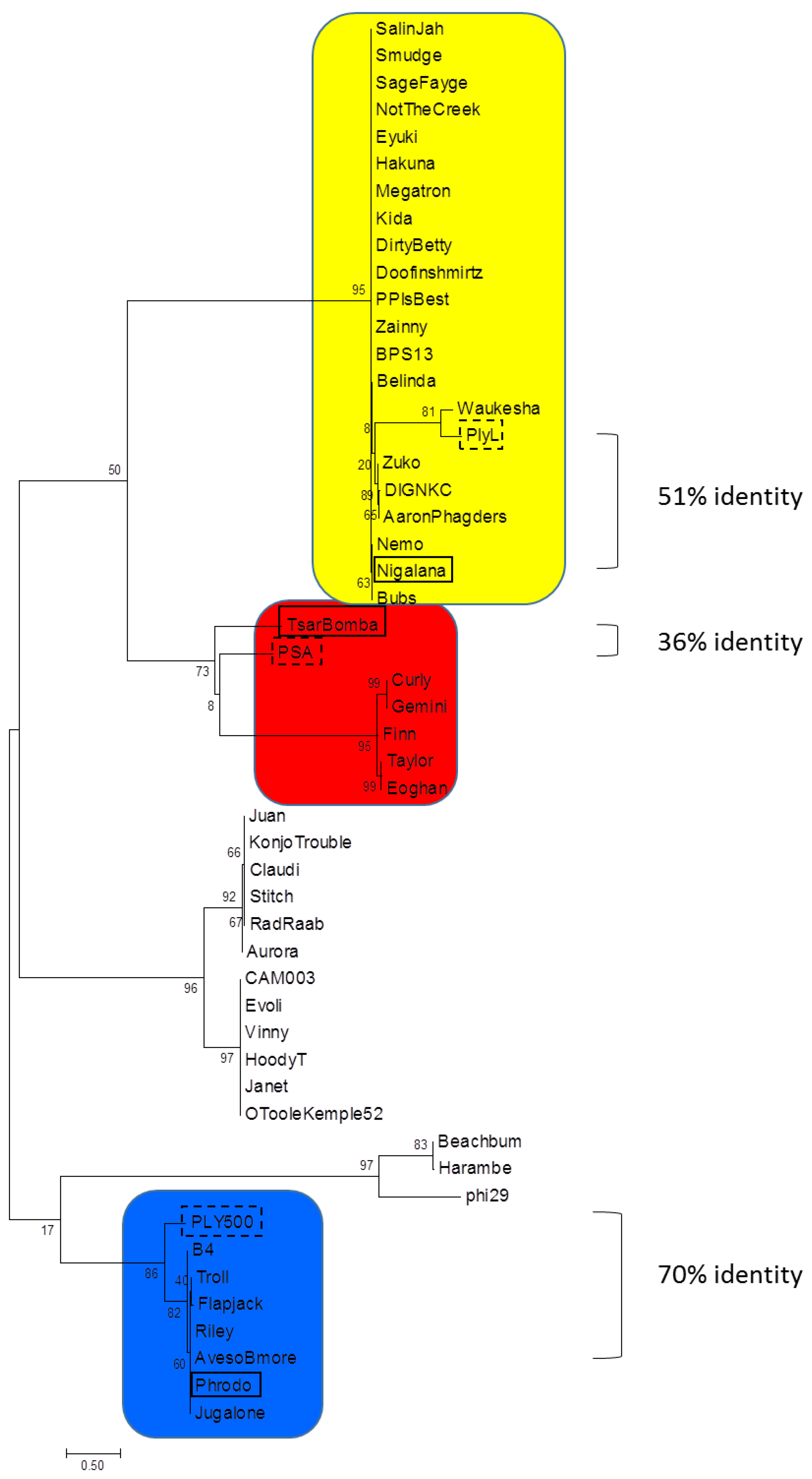
3.1. Phylogenetic Analysis
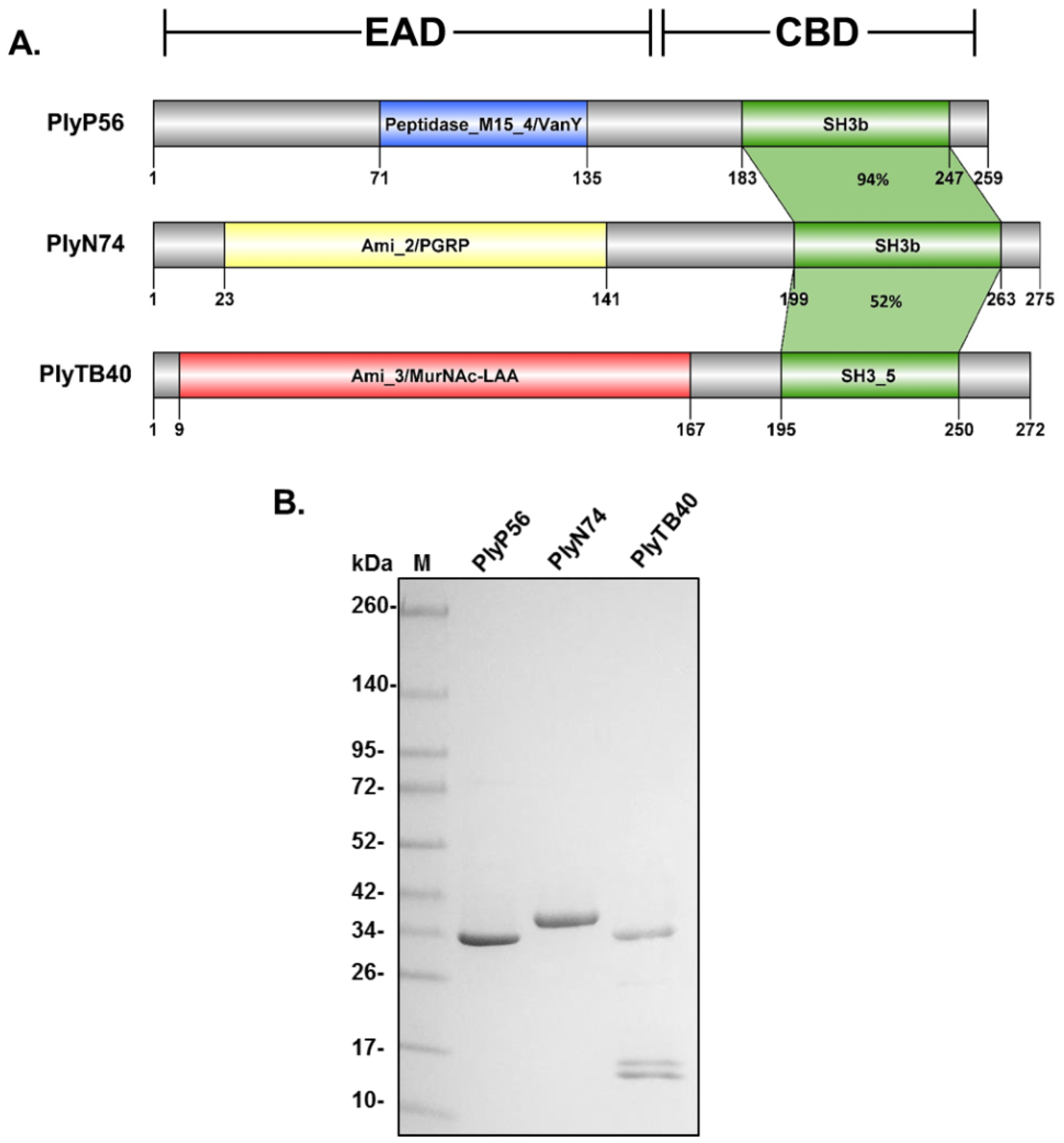
3.2. Endolysin Domain Architecture and Homology
3.3. Purification and Biochemical Characterization
3.4. Activity and Biochemical Characterization of Endolysins
3.5. Structural Modeling of Bacillus Bacteriophage Endolysin EADs
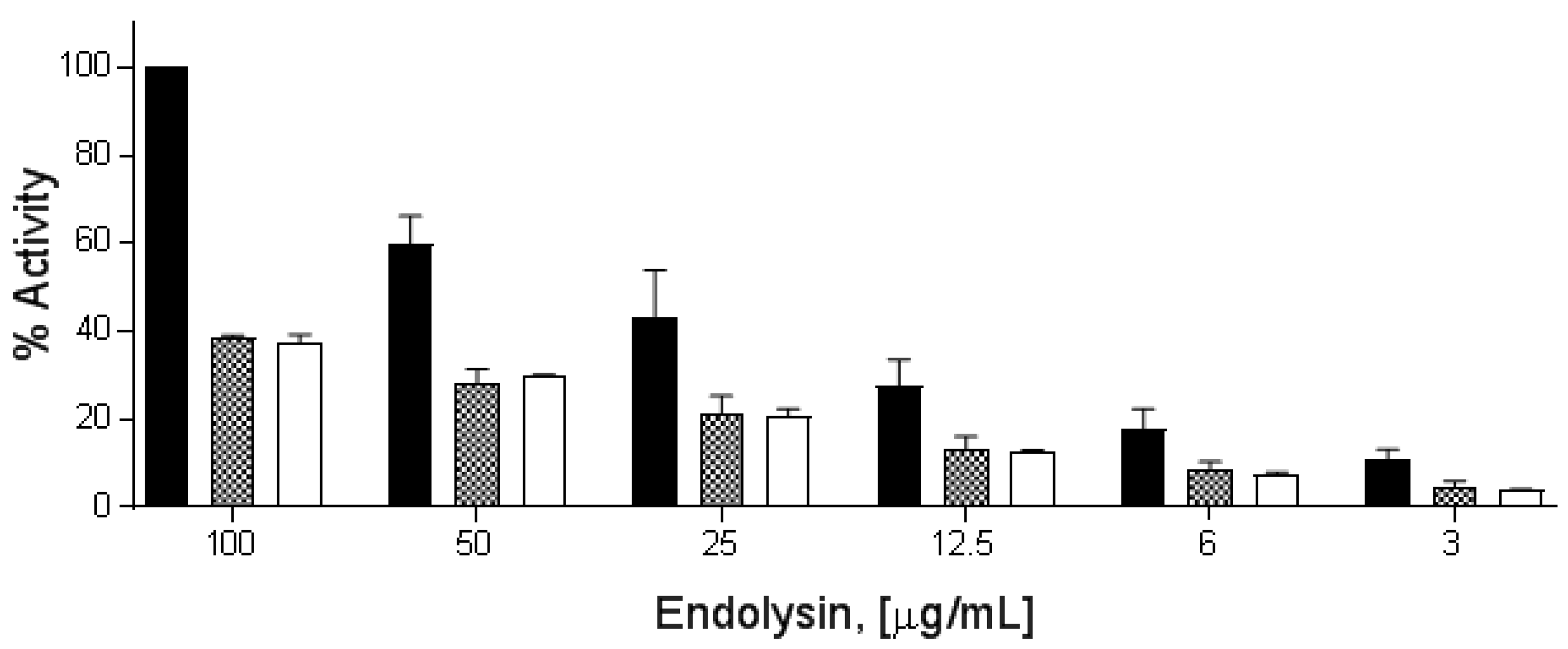
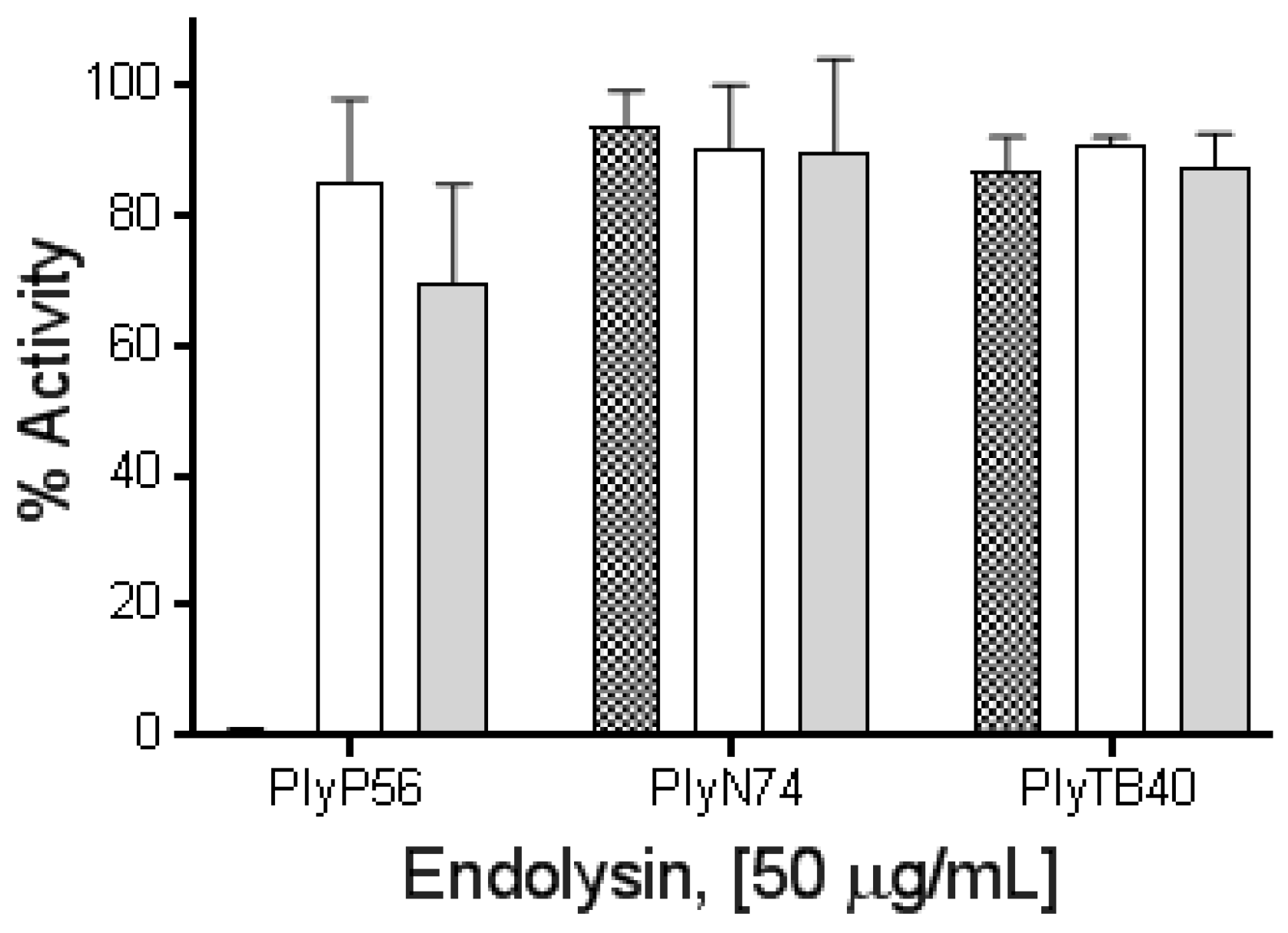
3.6. Effect of Divalent Metal Ions
3.7. Host Specificity
3.8. Cell Wall Binding
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leggett, M.J.; McDonnell, G.; Denyer, S.P.; Setlow, P.; Maillard, J.Y. Bacterial spore structures and their protective role in biocide resistance. J. Appl. Microbiol. 2012, 113, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, S.; Boon, N.; Uyttendaele, M. Diversity of Bacillus cereus group strains is reflected in their broad range of pathogenicity and diverse ecological lifestyles. FEMS Microbiol. Ecol. 2013, 84, 433–450. [Google Scholar] [CrossRef] [PubMed]
- Okinaka, R.T.; Keim, P. The Phylogeny of Bacillus cereus sensu lato. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Leggett, M.J.; Schwarz, J.S.; Burke, P.A.; McDonnell, G.; Denyer, S.P.; Maillard, J.Y. Resistance to and killing by the sporicidal microbicide peracetic acid. J. Antimicrob. Chemother. 2015, 70, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Gherardi, G. Bacillus cereus disease other than food-borne poisoning. In The Diverse Faces of Bacillus cereus; Savini, V., Ed.; Elsevier, Inc.: London, UK, 2016; pp. 93–116. [Google Scholar]
- Kutsuna, S.; Hayakawa, K.; Kita, K.; Katanami, Y.; Imakita, N.; Kasahara, K.; Seto, M.; Akazawa, K.; Shimizu, M.; Kano, T.; et al. Risk factors of catheter-related bloodstream infection caused by Bacillus cereus: Case-control study in 8 teaching hospitals in Japan. Am. J. Infect. Control 2017, 45, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Aronson, A.I. Insecticidal toxins. In Bacillus subtilis and Other Gram-Positive Bacteria; Sonenshein, A.L., Hoch, J.A., Losick, R., Eds.; American Society for Microbiology: Washington, DC, USA, 1993; pp. 953–963. [Google Scholar]
- Hoffmaster, A.R.; Hill, K.K.; Gee, J.E.; Marston, C.K.; De, B.K.; Popovic, T.; Sue, D.; Wilkins, P.P.; Avashia, S.B.; Drumgoole, R.; et al. Characterization of Bacillus cereus isolates associated with fatal pneumonias: Strains are closely related to Bacillus anthracis and harbor B. anthracis virulence genes. J. Clin. Microbiol. 2006, 44, 3352–3360. [Google Scholar] [CrossRef] [PubMed]
- Khasnabis, J.; Adhikari, P.; Chowdhury, D.; Rai, C.; Roy, A. Incidence of multiple drug resistant Bacillus cereus in some popular snacks and sweets sold in Kolkata city, India. Indian J. Microbiol. Res. 2017, 4, 14–19. [Google Scholar]
- Kim, C.W.; Cho, S.H.; Kang, S.H.; Park, Y.B.; Yoon, M.H.; Lee, J.B.; No, W.S.; Kim, J.B. Prevalence, genetic diversity, and antibiotic resistance of Bacillus cereus isolated from Korean fermented soybean products. J. Food Sci. 2015, 80, M123–M128. [Google Scholar] [CrossRef] [PubMed]
- Merzougui, S.; Lkhider, M.; Grosset, N.; Gautier, M.; Cohen, N. Prevalence, PFGE typing, and antibiotic resistance of Bacillus cereus group isolated from food in Morocco. Foodborne Pathog. Dis. 2014, 11, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Bacteriophage lytic enzymes: Novel anti-infectives. Trends Microbiol. 2005, 13, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Loessner, M.J. Bacteriophage endolysins—Current state of research and applications. Curr. Opin. Microbiol. 2005, 8, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Bacteriophage endolysins: A novel anti-infective to control Gram-positive pathogens. Int. J. Med. Microbiol. 2010, 300, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, J.M.; Nelson, D.; Fischetti, V.A. Rapid killing of Streptococcus pneumoniae with a bacteriophage cell wall hydrolase. Science 2001, 294, 2170–2172. [Google Scholar] [CrossRef] [PubMed]
- Royet, J.; Dziarski, R. Peptidoglycan recognition proteins: Pleiotropic sensors and effectors of antimicrobial defences. Nat. Rev. Microbiol. 2007, 5, 264–277. [Google Scholar] [CrossRef] [PubMed]
- Schuch, R.; Nelson, D.; Fischetti, V.A. A bacteriolytic agent that detects and kills Bacillus anthracis. Nature 2002, 418, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Barros, M.; Vennemann, T.; Gallagher, D.T.; Yin, Y.; Linden, S.B.; Heselpoth, R.D.; Spencer, D.J.; Donovan, D.M.; Moult, J.; et al. A bacteriophage endolysin that eliminates intracellular streptococci. Elife 2016, 5, e13152. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.C.; Schmelcher, M.; Rodriguez-Rubio, L.; Klumpp, J.; Pritchard, D.G.; Dong, S.; Donovan, D.M. Endolysins as antimicrobials. Adv. Virus Res. 2012, 83, 299–365. [Google Scholar] [PubMed]
- Schuch, R.; Pelzek, A.J.; Raz, A.; Euler, C.W.; Ryan, P.A.; Winer, B.Y.; Farnsworth, A.; Bhaskaran, S.S.; Stebbins, C.E.; Xu, Y.; et al. Use of a bacteriophage lysin to identify a novel target for antimicrobial development. PLoS ONE 2013, 8, e60754. [Google Scholar] [CrossRef] [PubMed]
- Korndörfer, I.P.; Danzer, J.; Schmelcher, M.; Zimmer, M.; Skerra, A.; Loessner, M.J. The crystal structure of the bacteriophage PSA endolysin reveals a unique fold responsible for specific recognition of Listeria cell walls. J. Mol. Biol. 2006, 364, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Korndörfer, I.P.; Kanitz, A.; Danzer, J.; Zimmer, M.; Loessner, M.J.; Skerra, A. Structural analysis of the l-alanoyl-d-glutamate endopeptidase domain of Listeria bacteriophage endolysin Ply500 reveals a new member of the LAS peptidase family. Acta Crystallogr. Sect. D Biol. Crystallogr. 2008, 64, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Low, L.Y.; Yang, C.; Perego, M.; Osterman, A.; Liddington, R.C. Structure and lytic activity of a Bacillus anthracis prophage endolysin. J. Biol. Chem. 2005, 280, 35433–35439. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Yun, J.; Lim, J.A.; Kang, D.H.; Ryu, S. Characterization of an endolysin, LysBPS13, from a Bacillus cereus bacteriophage. FEMS Microbiol. Lett. 2012, 332, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Saedi, M.S.; Garvey, K.J.; Ito, J. Cloning and purification of a unique lysozyme produced by Bacillus phage phi 29. Proc. Natl. Acad. Sci. USA 1987, 84, 955–958. [Google Scholar] [CrossRef] [PubMed]
- Son, B.; Yun, J.; Lim, J.A.; Shin, H.; Heu, S.; Ryu, S. Characterization of LysB4, an endolysin from the Bacillus cereus-infecting bacteriophage B4. BMC Microbiol. 2012, 12, 33. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Söding, J.; Biegert, A.; Lupas, A.N. The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res. 2005, 33, W244–W248. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Fiser, A.; Do, R.K.G.; Šali, A. Modeling of loops in protein structures. Protein Sci. 2000, 9, 1753–1773. [Google Scholar] [CrossRef] [PubMed]
- Šali, A.; Blundell, T.L. Comparative protein modelling by satisfaction of spatial restraints. J. Mol. Biol. 1993, 234, 779–815. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.-Y.; Sali, A. Statistical potential for assessment and prediction of protein structures. Protein Sci. 2006, 15, 2507–2524. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Jordan, T.C.; Burnett, S.H.; Carson, S.; Caruso, S.M.; Clase, K.; DeJong, R.J.; Dennehy, J.J.; Denver, D.R.; Dunbar, D.; Elgin, S.C.; et al. A broadly implementable research course in phage discovery and genomics for first-year undergraduate students. MBio 2014, 5, e01051-13. [Google Scholar] [CrossRef] [PubMed]
- Büttner, F.M.; Renner-Schneck, M.; Stehle, T. X-Ray crystallography and its impact on understanding bacterial cell wall remodeling processes. Int. J. Med. Microbiol. 2015, 305, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Loessner, M.J.; Maier, S.K.; Daubek-Puza, H.; Wendlinger, G.; Scherer, S. Three Bacillus cereus bacteriophage endolysins are unrelated but reveal high homology to cell wall hydrolases from different bacilli. J. Bacteriol. 1997, 179, 2845–2851. [Google Scholar] [CrossRef] [PubMed]
- Dunne, M.; Leicht, S.; Krichel, B.; Mertens, H.D.; Thompson, A.; Krijgsveld, J.; Svergun, D.I.; Gomez-Torres, N.; Garde, S.; Uetrecht, C.; et al. Crystal structure of the CTP1L endolysin reveals how its activity is regulated by a secondary translation product. J. Biol. Chem. 2016, 291, 4882–4893. [Google Scholar] [CrossRef] [PubMed]
- Proenca, D.; Velours, C.; Leandro, C.; Garcia, M.; Pimentel, M.; Sao-Jose, C. A two-component, multimeric endolysin encoded by a single gene. Mol. Microbiol. 2015, 95, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.; Martinez, B.; Rodriguez, L.; Rodriguez, A. Synergy between the phage endolysin LysH5 and nisin to kill Staphylococcus aureus in pasteurized milk. Int. J. Food Microbiol. 2010, 141, 151–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Peng, Q.; Gao, M. Characteristics of a broad lytic spectrum endolysin from phage BtCS33 of Bacillus thuringiensis. BMC Microbiol. 2012, 12, 297. [Google Scholar] [CrossRef] [PubMed]
- Yoong, P.; Schuch, R.; Nelson, D.; Fischetti, V.A. PlyPH, a bacteriolytic enzyme with a broad pH range of activity and lytic action against Bacillus anthracis. J. Bacteriol. 2006, 188, 2711–2714. [Google Scholar] [CrossRef] [PubMed]
- Linden, S.B.; Zhang, H.; Heselpoth, R.D.; Shen, Y.; Schmelcher, M.; Eichenseher, F.; Nelson, D.C. Biochemical and biophysical characterization of PlyGRCS, a bacteriophage endolysin active against methicillin-resistant Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2015, 99, 741–752. [Google Scholar] [CrossRef] [PubMed]
- Firczuk, M.; Bochtler, M. Folds and activities of peptidoglycan amidases. FEMS Microbiol. Lett. 2007, 31, 676–691. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed]
- Cerda-Costa, N.; Gomis-Ruth, F.X. Architecture and function of metallopeptidase catalytic domains. Protein Sci. 2014, 23, 123–144. [Google Scholar] [CrossRef] [PubMed]
- Helgason, E.; Okstad, O.A.; Caugant, D.A.; Johansen, H.A.; Fouet, A.; Mock, M.; Hegna, I.; Kolsto, A.B. Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis—One species on the basis of genetic evidence. Appl. Environ. Microbiol. 2000, 66, 2627–2630. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Oliveira, H.; Melo, L.D.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: Their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuch, R.; Fischetti, V.A.; Nelson, D.C. A genetic screen to identify bacteriophage lysins. Methods Mol. Biol. 2009, 502, 307–319. [Google Scholar] [PubMed]
- Schmelcher, M.; Loessner, M.J. Application of bacteriophages for detection of foodborne pathogens. Bacteriophage 2014, 4, e28137. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, D.G.; Lessard, I.A.; Walsh, C.T. Mutational analysis of potential zinc-binding residues in the active site of the enterococcal D-Ala-D-Ala dipeptidase VanX. Biochemistry 1997, 36, 10498–10505. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Melo, L.D.; Santos, S.B.; Nobrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [PubMed]
- Low, L.Y.; Yang, C.; Perego, M.; Osterman, A.; Liddington, R. Role of net charge on catalytic domain and influence of cell wall binding domain on bactericidal activity, specificity, and host range of phage lysins. J. Biol. Chem. 2011, 286, 34391–34403. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.J.; Garefalaki, V.; Spoerl, R.; Narbad, A.; Meijers, R. Structure-based modification of a Clostridium difficile-targeting endolysin affects activity and host range. J. Bacteriol. 2011, 193, 5477–5486. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | EAD | CBD | Examples |
|---|---|---|---|
| I. | G25 muramidase | Amidase02_C | Vinny ORF63 |
| II. | G25 PlyB-like | Amidase02_C | Stitch ORF31 |
| III. | MurNAc-LAA | 2X LysM | Taylor ORF31 |
| IV. | MurNAc-LAA | SH3 | TsarBomba ORF40 (PlyTB40) |
| V. | VanY | 2X PG_binding_1 | SPO1 ORF107 |
| VI. | Peptidase M15_4/VanY | SH3 | Phrodo ORF56 (PlyP56) |
| VII. | GH24 muramidase | SH3 | Beachbum ORF23 |
| VIII. | PGRP | Amidase02_C | Waukesha ORF68 |
| IX. | PGRP | SH3 | Nigalana ORF74 (PlyN74) |
| Species | Strain 1 | Bacteriophage Endolysins 2 | ||
|---|---|---|---|---|
| PlyP56 | PlyN74 | PlyTB40 | ||
| B. cereus | ATCC 4342 | 84.9 ± 6.0 | 69.2 ± 9.8 | 71.9 ± 12.9 |
| B. cereus | ATCC 14579 | 73.4 ± 1.3 | 59.7 ± 7.2 | 40.9 ± 20.5 |
| B. cereus | ATCC 11778 | 79.6 ± 4.4 | 60.6 ± 4.4 | 58.2 ± 13.2 |
| B. cereus | ATCC 13061 | 45.8 ± 2.4 | 37.9 ± 3.8 | 24.0 ± 8.1 |
| B. thuringiensis | ATCC 10792 | 38.7 ± 5.2 | 35.8 ± 9.1 | 25.6 ± 6.6 |
| B. amyloliquefaciens | ATCC 23842 | 36.5 ± 20.5 | 23.9 ± 13.8 | 6.2 ± 2.4 |
| B. circulans | ATCC 4513 | 53.2 ± 3.2 | 17.2 ± 5.1 | 5.5 ± 5.1 |
| B. coagulans | ATCC 7050 | 8.0 ± 1.7 | 5.7 ± 7.9 | 4.6 ± 4.5 |
| B. licheniformis | ATCC 14580 | 4.6 ± 6.4 | 30.2 ± 3.6 | 9.6 ± 3.7 |
| B. megaterium | ATCC 14581 | 83.9 ± 12.3 | 30.2 ± 15.5 | 9.6 ± 1.6 |
| B. pumilus | BJ0050 | 58.8 ± 15.2 | 46.1 ± 11.1 | 32.6 ± 14.4 |
| B. pumilus | ATCC 700814 | 16.7 ± 18.4 | 10.9 ± 12.5 | 2.6 ± 4.3 |
| B. subtilis | ATCC 6051 | 3.5 ± 1.9 | 1.4 ± 0.5 | 0.1 ± 0.2 |
| B. subtilis | ATCC 33608 | 2.9 ± 2.3 | 1.6 ± 2.7 | 0.6 ± 0.8 |
| Lysinb. Sphaericus | ATCC 4525 | 36.9 ± 19.8 | 18.9 ± 9.4 | 10.8 ± 3.4 |
| Paenib. Polymyxa | ATCC 7070 | 6.0 ± 4.7 | 3.7 ± 4.3 | 3.1 ± 1.5 |
| Species | Strain 1 | Bacteriophage Endolysins 2 | ||
|---|---|---|---|---|
| PlyP56 | PlyN74 | PlyTB40 | ||
| B. cereus | ATCC 4342 | +++ | +++ | ++ |
| B. anthracis | Ames 35 | ++ | +++ | + |
| B. anthracis | UM23 | + | +/− | +/− |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Etobayeva, I.; Linden, S.B.; Alem, F.; Harb, L.; Rizkalla, L.; Mosier, P.D.; Johnson, A.A.; Temple, L.; Hakami, R.M.; Nelson, D.C. Discovery and Biochemical Characterization of PlyP56, PlyN74, and PlyTB40—Bacillus Specific Endolysins. Viruses 2018, 10, 276. https://doi.org/10.3390/v10050276
Etobayeva I, Linden SB, Alem F, Harb L, Rizkalla L, Mosier PD, Johnson AA, Temple L, Hakami RM, Nelson DC. Discovery and Biochemical Characterization of PlyP56, PlyN74, and PlyTB40—Bacillus Specific Endolysins. Viruses. 2018; 10(5):276. https://doi.org/10.3390/v10050276
Chicago/Turabian StyleEtobayeva, Irina, Sara B. Linden, Farhang Alem, Laith Harb, Lucas Rizkalla, Philip D. Mosier, Allison A. Johnson, Louise Temple, Ramin M. Hakami, and Daniel C. Nelson. 2018. "Discovery and Biochemical Characterization of PlyP56, PlyN74, and PlyTB40—Bacillus Specific Endolysins" Viruses 10, no. 5: 276. https://doi.org/10.3390/v10050276
APA StyleEtobayeva, I., Linden, S. B., Alem, F., Harb, L., Rizkalla, L., Mosier, P. D., Johnson, A. A., Temple, L., Hakami, R. M., & Nelson, D. C. (2018). Discovery and Biochemical Characterization of PlyP56, PlyN74, and PlyTB40—Bacillus Specific Endolysins. Viruses, 10(5), 276. https://doi.org/10.3390/v10050276