An Ointment Consisting of the Phage Lysin LysGH15 and Apigenin for Decolonization of Methicillin-Resistant Staphylococcus aureus from Skin Wounds



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Strains
2.3. LysGH15-Api-Aquaphor Ointment Preparation
2.4. In Vitro Activity of LysGH15-Api-Aquaphor Ointment
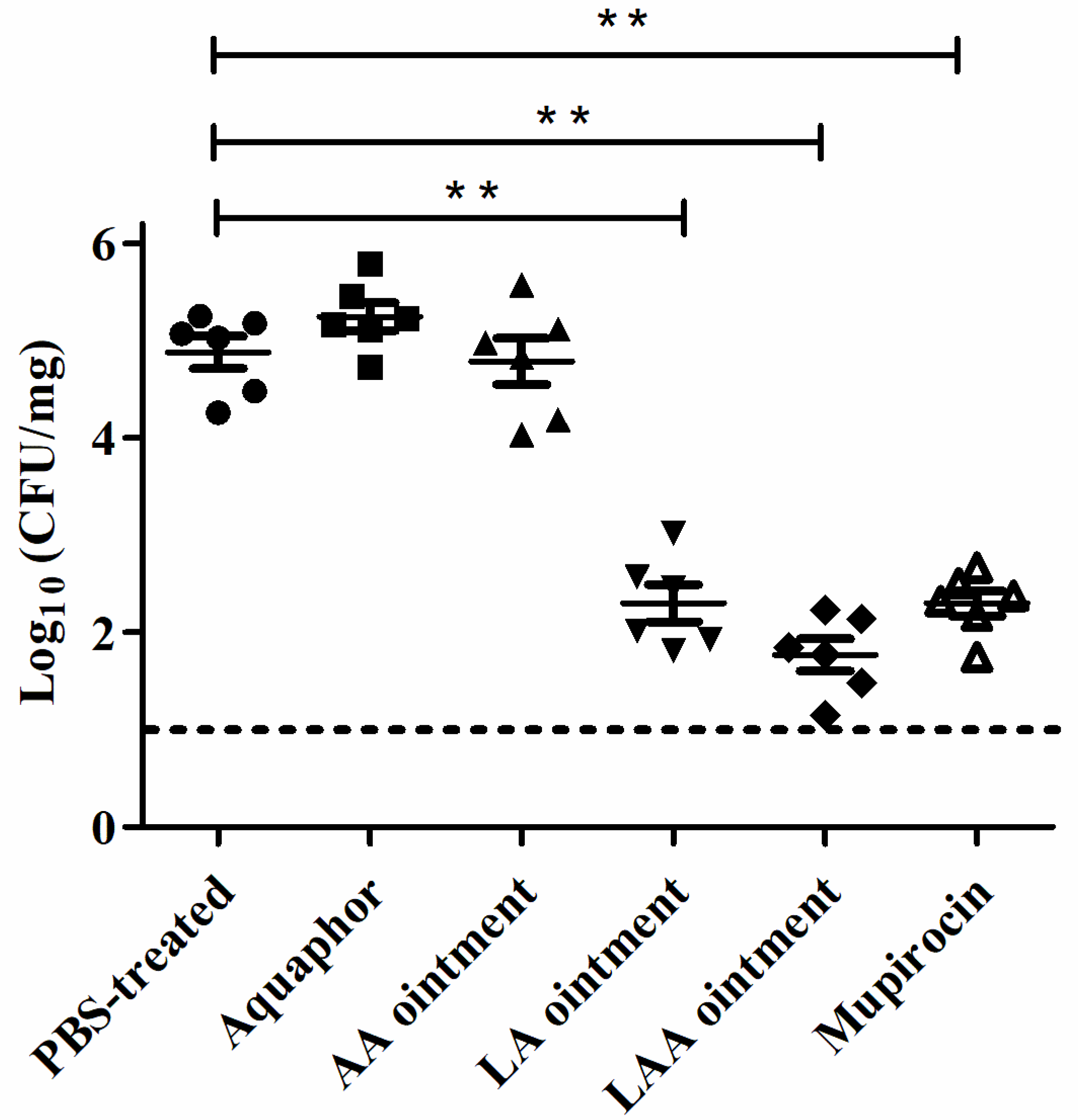
2.5. Mouse Model of Skin Damage and Topical Treatment
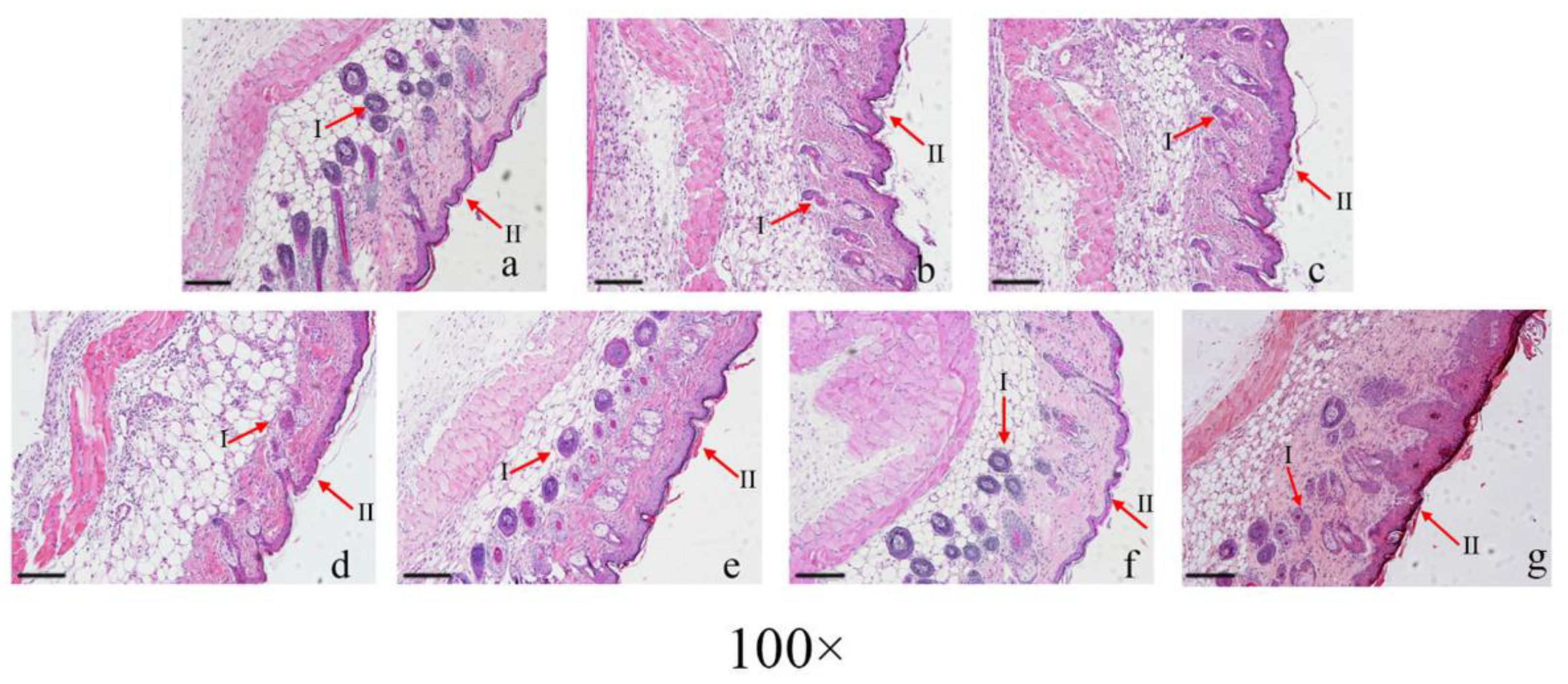
2.6. Histology
2.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
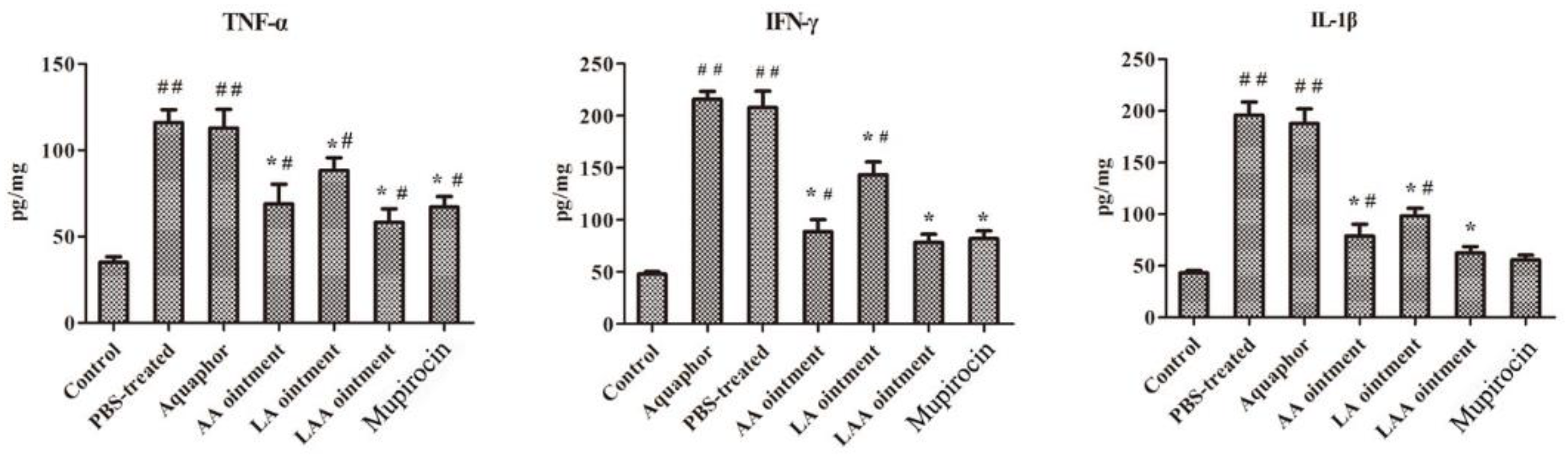
2.8. Immunological Assays
2.9. Data Analysis
3. Results
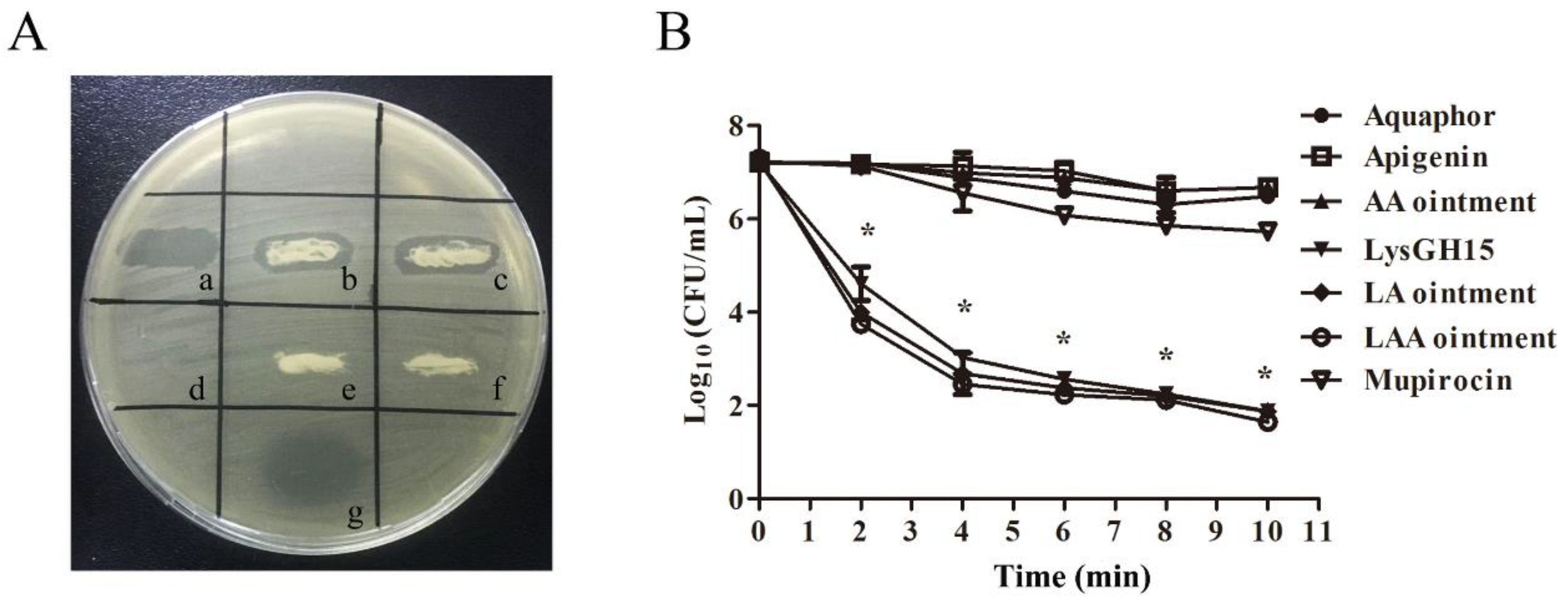
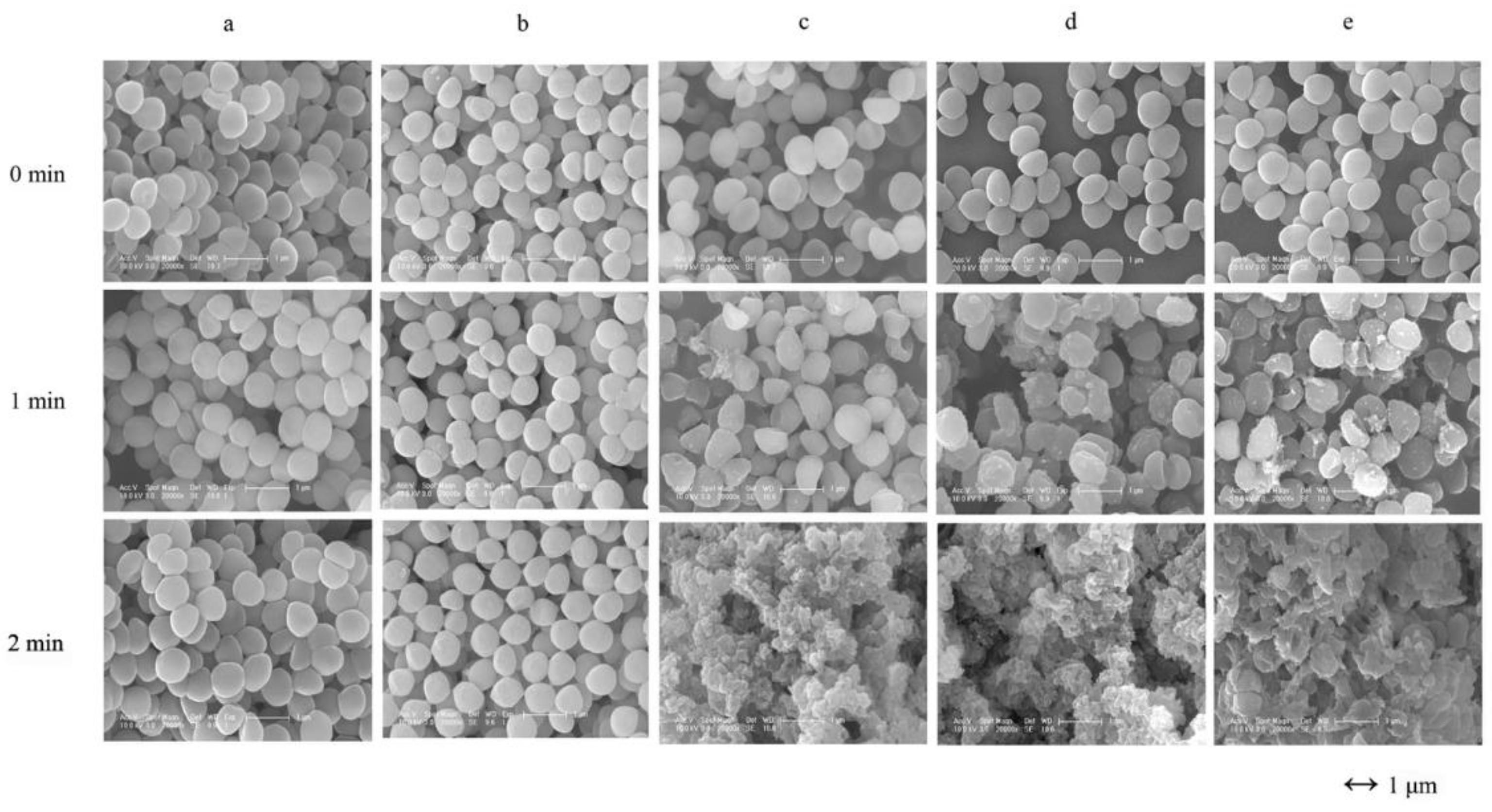
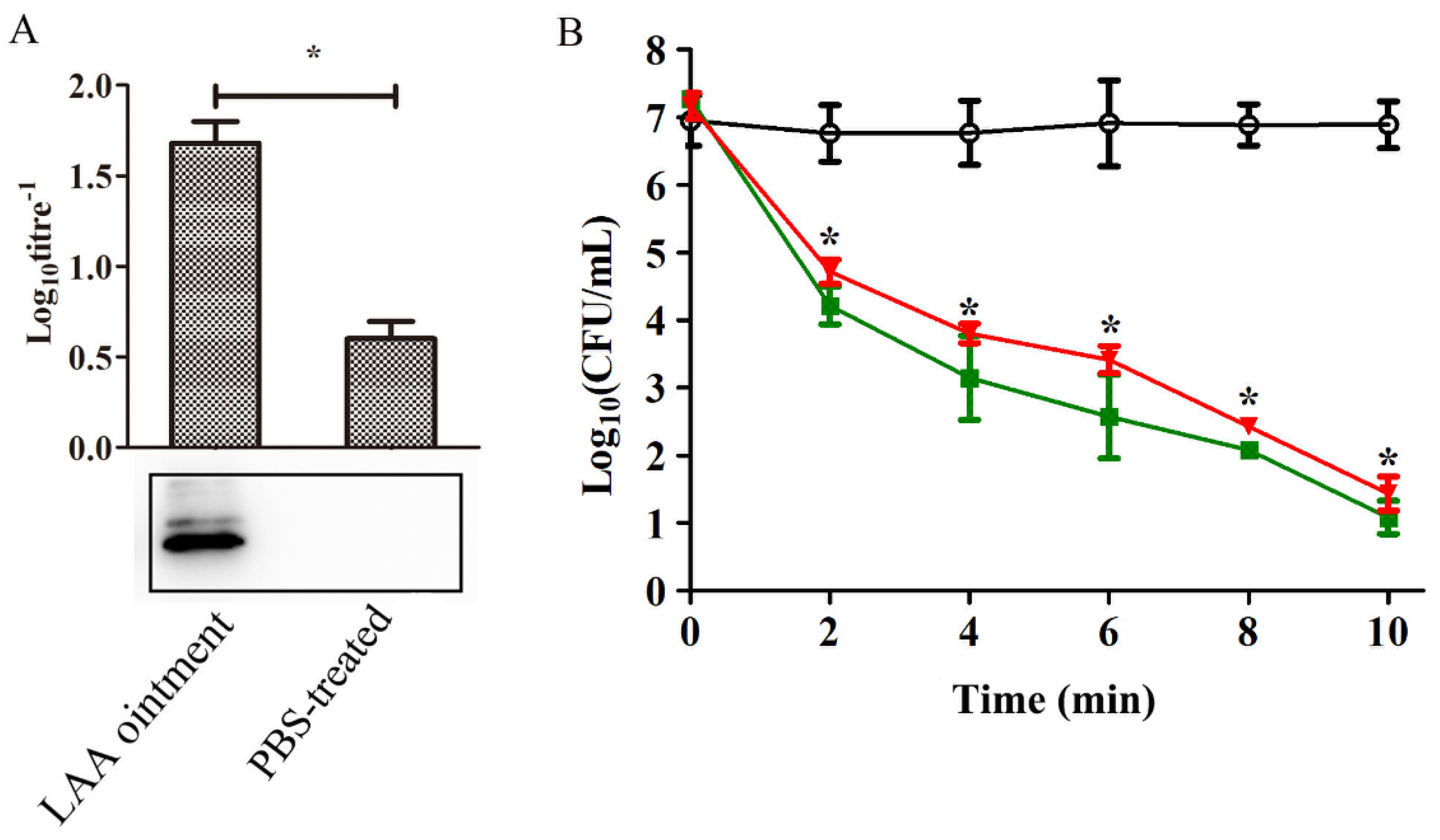
3.1. In Vitro Lytic Activity and Haemolysis Inhibition by LAA
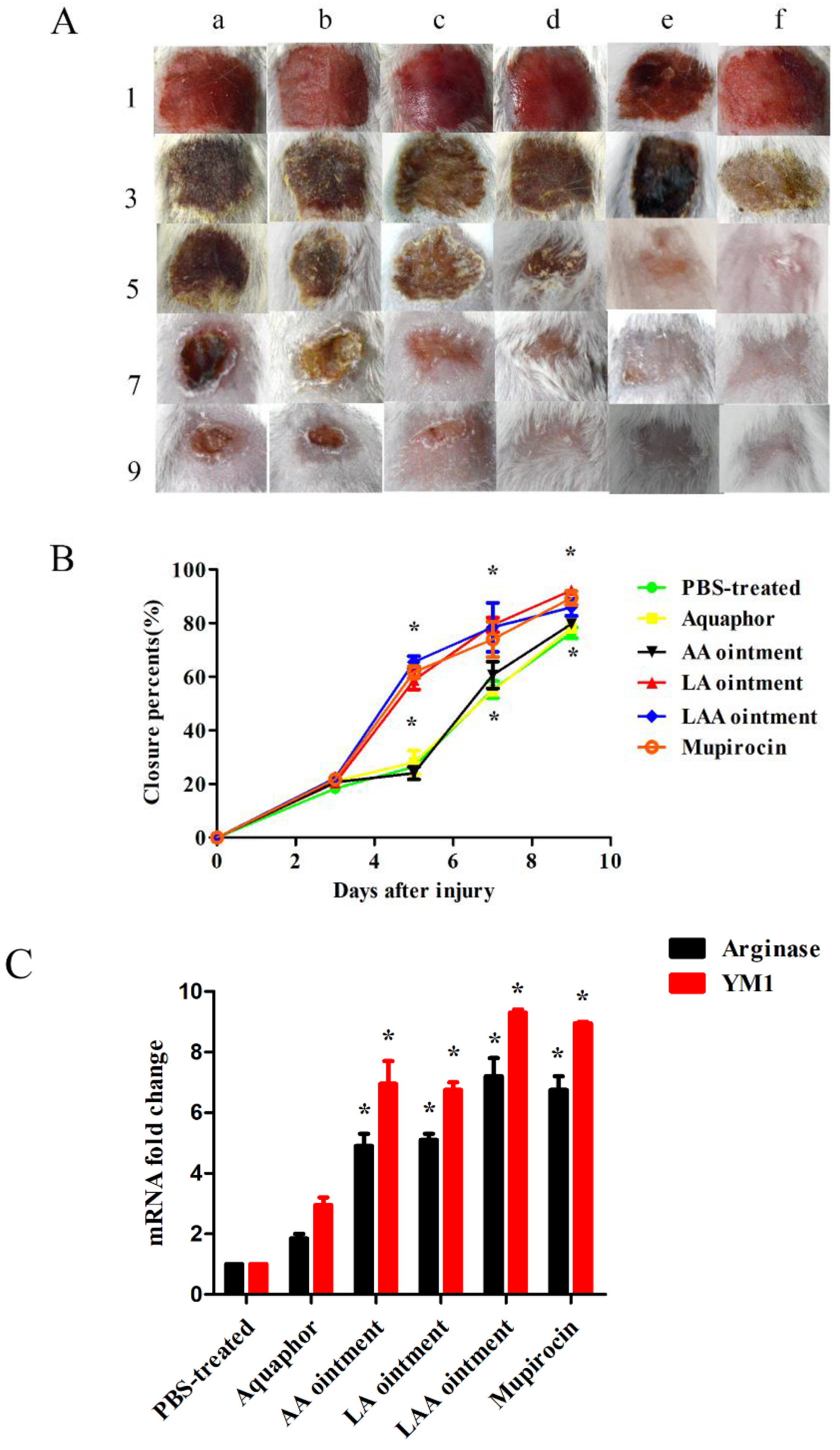
3.2. LAA Accelerates Skin Wound Healing
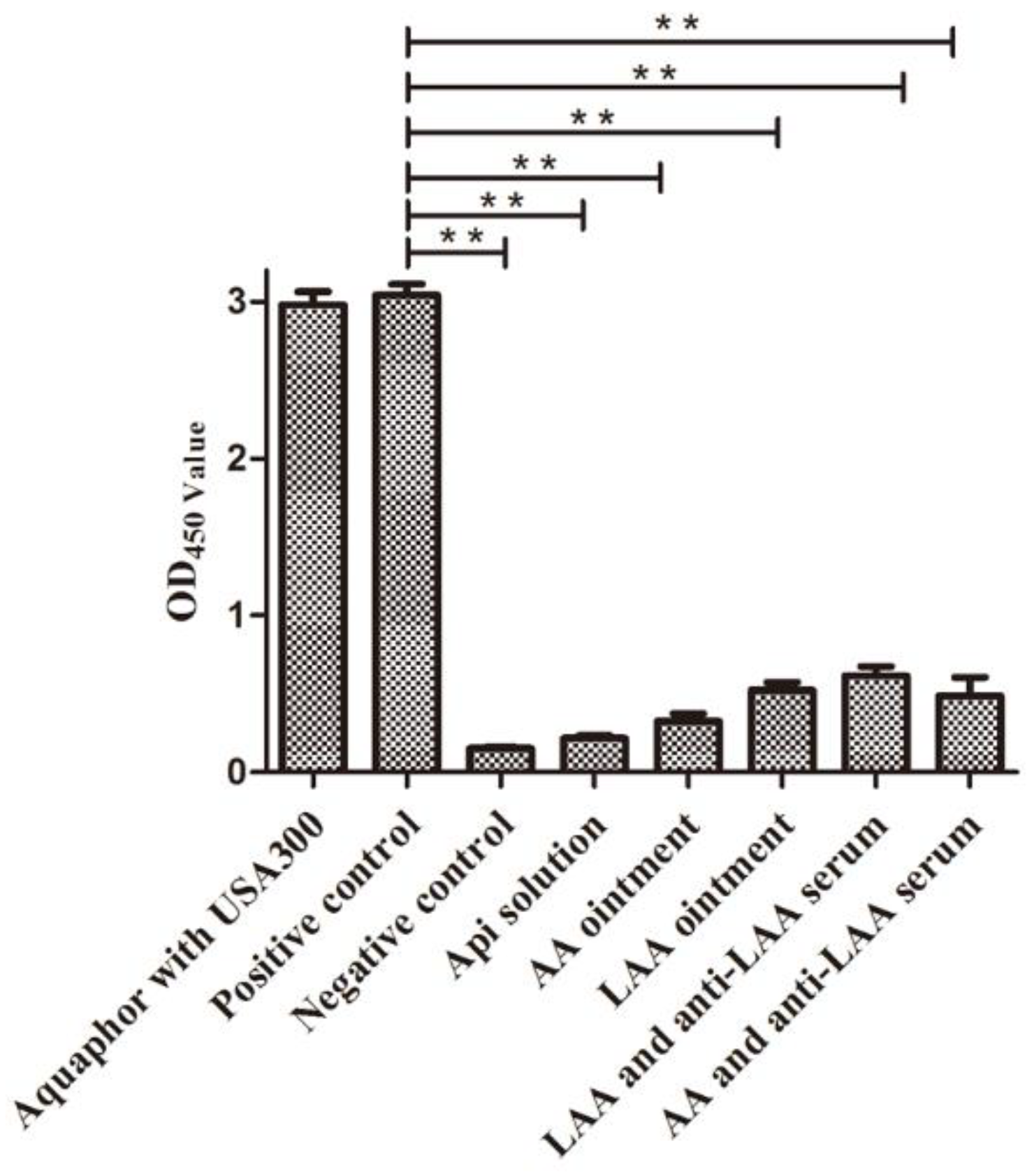
3.3. Anti-LAA Serum Did not Affect the Activity of LAA
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| S. aureus | Staphylococcus aureus |
| MRSA | methicillin-resistant Staphylococcus aureus |
| MSSA | methicillin-susceptible Staphylococcus aureus |
| CHAP | cysteine, histidine-dependent amidohydrolases/peptidases |
| SH3b | Src homology 3b |
| Api | Apigenin |
| ATCC | American Type Culture Collection |
| BHI | brain heart infusion |
| PBS | phosphate-buffered saline |
| LAA | LysGH15-api-Aquaphor |
| LA | LysGH15-Aquaphor |
| AA | Api-Aquaphor |
| SEM | Scanning electron microscopy |
| AAM | alternatively activated macrophages |
| ELISA | enzyme-linked immunosorbent assay |
| H&E | hematoxylin and eosin |
References
- Peacock, J.J.; Marsik, F.J.; Wenzel, R.P. Methicillin-resistant Staphylococcus aureus: Introduction and spread within a hospital. Ann. Intern. Med. 1980, 93, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.J. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Iruloh, C.G.; Bonner, S.; Ma, K. Methicillin-sensitive Staphylococcus aureus chorioamnionitis and foetal death after mechanical induction of labour: A case report. J. Obstet. Gynaecol. 2017, 38, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Sasson, G.; Bai, A.D.; Showler, A.; Burry, L.; Steinberg, M.; Ricciuto, D.R.; Fernandes, T.; Chiu, A.; Raybardhan, S.; Science, M.; et al. Staphylococcus aureus bacteremia in immunosuppressed patients: A multicenter, retrospective cohort study. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Novel Method to Control Pathogenic Bacteria on Human Mucous Membranes. Ann. N. Y. Acad. Sci. 2003, 987, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Kugelberg, E.; Norstrom, T.; Petersen, T.K.; Duvold, T.; Andersson, D.I.; Hughes, D. Establishment of a superficial skin infection model in mice by using Staphylococcus aureus and Streptococcus pyogenes. Antimicrob. Agents Chemother. 2005, 49, 3435–3441. [Google Scholar] [CrossRef] [PubMed]
- David, M.Z.; Daum, R.S. Treatment of Staphylococcus aureus Infections. Curr. Top. Microbiol. Immunol. 2017, 409, 325–383. [Google Scholar] [PubMed]
- Peterson, L.R.; Woods, C.W.; Davis, T.J.; Wang, Z.X.; Young, S.A.; Osiecki, J.C.; Lewinski, M.A.; Liesenfeld, O. Performance of the cobas MRSA/SA Test for Simultaneous Detection of Methicillin-Susceptible and Methicillin-Resistant Staphylococcus aureus From Nasal Swabs. Am. J. Clin. Pathol. 2017, 148, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Davis, K.A.; Stewart, J.J.; Crouch, H.K.; Florez, C.E.; Hospenthal, D.R. Methicillin-resistant Staphylococcus aureus (MRSA) nares colonization at hospital admission and its effect on subsequent MRSA infection. Clin. Infect. Dis. 2004, 39, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.Q.; Lin, L.; Lin, T.; Hao, F.; Zeng, F.Q.; Bi, Z.G.; Yi, D.; Zhao, B. Skin colonization by Staphylococcus aureus in patients with eczema and atopic dermatitis and relevant combined topical therapy: A double-blind multicentre randomized controlled trial. Br. J. Dermatol. 2006, 155, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Kahanov, L.; Kim, Y.K.; Eberman, L.; Dannelly, K.; Kaur, H.; Ramalinga, A. Staphylococcus aureus and community-associated methicillin-resistant Staphylococcus aureus (CA-MRSA) in and around therapeutic whirlpools in college athletic training rooms. J. Athl. Train. 2015, 50, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Platt, R. Risk of methicillin-resistant Staphylococcus aureus infection after previous infection or colonization. Clin. Infect. Dis. 2003, 36, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Van Hal, S.J.; Jensen, S.O.; Vaska, V.L.; Espedido, B.A.; Paterson, D.L.; Gosbell, I.B. Predictors of mortality in Staphylococcus aureus Bacteremia. Clin. Microbiol. Rev. 2012, 25, 362–386. [Google Scholar] [CrossRef] [PubMed]
- Lentino, J.R.; Baddour, L.M.; Wray, M.; Wong, E.S.; Yu, V.L. Staphylococcus aureus and other bacteremias in hemodialysis patients: Antibiotic therapy and surgical removal of access site. Infection 2000, 28, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Van Rijen, M.; Bonten, M.; Wenzel, R.; Kluytmans, J. Mupirocin ointment for preventing Staphylococcus aureus infections in nasal carriers. Cochrane Database Syst. Rev. 2008, 4, CD006216. [Google Scholar] [CrossRef] [PubMed]
- Greenhalgh, D.G. Topical Antimicrobial Agents for Burn Wounds. Clin. Plast. Surg. 2009, 36, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.M.; Xu, W.; Lei, L.C.; Huang, J.; Feng, X.; Sun, C.J.; Du, C.T.; Zuo, J.; Li, Y.; Du, T.; et al. LysGH15, a novel bacteriophage lysin, protects a murine bacteremia model efficiently against lethal methicillin-resistant Staphylococcus aureus infection. J. Clin. Microbiol. 2011, 49, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.F.; Li, X.; Wang, B.; Gong, P.J.; Xiao, F.; Yang, M.; Zhang, L.; Song, J.; Hu, L.Y.; Cheng, M.J.; et al. Combination Therapy of LysGH15 and Apigenin as a New Strategy for Treating Pneumonia Caused by Staphylococcus aureus. Appl. Environ. Microbiol. 2015, 82, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, D.; Li, X.W.; Hu, L.Y.; Cheng, M.J.; Xia, F.F.; Gong, P.J.; Wang, B.; Ge, J.L.; Zhang, H.; et al. LysGH15 kills Staphylococcus aureus without being affected by the humoral immune response or inducing inflammation. Sci. Rep. 2016, 6, 29344. [Google Scholar] [CrossRef] [PubMed]
- Rashel, M.; Uchiyama, J.; Ujihara, T.; Uehara, Y.; Kuramoto, S.; Sugihara, S.; Yagyu, K.I.; Muraoka, A.; Sugai, M.; Hiramatsu, K.; et al. Efficient Elimination of Multidrug-Resistant Staphylococcus aureus by Cloned Lysin Derived from Bacteriophage φMR11. J. Infect. Dis. 2007, 196, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Idelevich, E.A.; von Eiff, C.; Friedrich, A.W.; Iannelli, D.; Xia, G.; Peters, G.; Peschel, A.; Wanninger, I.; Becker, K. In vitro activity against Staphylococcus aureus of a novel antimicrobial agent, PRF-119, a recombinant chimeric bacteriophage endolysin. Antimicrob. Agents Chemother. 2011, 55, 4416–4419. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yu, J.P.; Wei, H.P. Engineered bacteriophage lysins as novel anti-infectives. Front. Microbiol. 2014, 5, 542. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.M.; Feng, Y.; Feng, X.; Sun, C.J.; Lei, L.C.; Ding, W.; Niu, F.; Jiao, L.; Yang, M.; Li, Y.; et al. Structural and biochemical characterization reveals LysGH15 as an unprecedented “EF-hand-like” calcium-binding phage lysin. PLoS Pathog. 2014, 10, e1004109. [Google Scholar] [CrossRef] [PubMed]
- Duthie, G.; Crozier, A. Plant-derived phenolic antioxidants. Curr. Opin. Lipidol. 2000, 11, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Li, R.W.; David, L.G.; Myers, S.P.; Leach, D.N. Anti-inflammatory activity of Chinese medicinal vine plants. J. Ethnopharmacol. 2003, 85, 61–67. [Google Scholar] [CrossRef]
- Cesca, T.G.; Faqueti, L.G.; Rocha, L.W.; Meira, N.A.; Meyre-Silva, C.; de Souza, M.M.; Quintão, N.L.M.; Silva, R.M.L.; Filho, V.C.; Bresolin, T.M.B. Antinociceptive, anti-inflammatory and wound healing features in animal models treated with a semisolid herbal medicine based on Aleurites moluccana L. Willd. Euforbiaceae standardized leaf extract. J. Ethnopharmacol. 2012, 143, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.; Park, J.; Lee, E.; Lim, S.; Yu, J.G.; Lee, S.J.; Chen, H.; Dong, Z.; Lee, K.W.; Lee, H.J. Src kinase is a direct target of apigenin against UVB-induced skin inflammation. Carcinogenesis 2013, 34, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Jornet, P.; Camacho-Alonso, F.; Gomez-Garcia, F.; Molina, M.F.; Canas, X.; Serafin, A.; Castillo, J.; Vicente-Ortega, V. Effects of potassium apigenin and verbena extract on the wound healing process of SKH-1 mouse skin. Int. Wound J. 2014, 11, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Ho, Y.; Hsieh, W.; Huang, S.; Chang, Y.; Huang, G. Apigenin-7-Glycoside Prevents LPS-Induced Acute Lung Injury via Downregulation of Oxidative Enzyme Expression and Protein Activation through Inhibition of MAPK Phosphorylation. Int. J. Mol. Sci. 2015, 16, 1736–1754. [Google Scholar] [CrossRef] [PubMed]
- Patricia, M.P.; Francisco, G.G.; Francisco, M.D.; Rui, A.M.; Pia, L.J. The therapeutic effects of apigenin and dexamethasone on 5-fluorouracil-induced oral mucositis—A pilot study using a Syrian hamster model. J. Oral Pathol. Med. 2016, 46, 142–147. [Google Scholar]
- Dong, J.; Qiu, J.Z.; Wang, J.F.; Li, H.G.; Dai, X.H.; Zhang, Y.; Wang, X.; Tan, W.; Niu, X.D.; Deng, X.M.; et al. Apigenin alleviates the symptoms of Staphylococcus aureus pneumonia by inhibiting the production of alpha-hemolysin. FEMS Microbiol. Lett. 2013, 338, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Pastagia, M.; Euler, C.; Chahales, P.; Fuentes-Duculan, J.; Krueger, J.G.; Fischetti, V.A. A Novel Chimeric Lysin Shows Superiority to Mupirocin for Skin Decolonization of Methicillin-Resistant and—Sensitive Staphylococcus aureus Strains. Antimicrob. Agents Chemother. 2011, 55, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.C.; Zhang, Y.J.; Cai, Q.; Li, B.S.; Yang, H.; Wang, H.; Qi, H.; Zhou, Y.M.; Meng, W.Y. Effects of the micro-nano surface topography of titanium alloy on the biological responses of osteoblast. J. Biomed. Mater. Res. Part A 2016, 105, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Worlitzsch, D.; Kaygin, H.; Steinhuber, A.; Dalhoff, A.; Botzenhart, K.; Doring, G. Effects of amoxicillin, gentamicin, and moxifloxacin on the hemolytic activity of Staphylococcus aureus in vitro and in vivo. Antimicrob. Agents Chemother. 2001, 45, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Li, X.Y.; Hu, S.L.; Liu, T.; Yuan, B.H.; Gu, H.B.; Ni, Q.; Zhang, X.F.; Zheng, F. IL-33 accelerates cutaneous wound healing involved in upregulation of alternatively activated macrophages. Mol. Immunol. 2013, 56, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Rak, G.D.; Osborne, L.C.; Siracusa, M.C.; Kim, B.S.; Wang, K.; Bayat, A.; Artis, D.; Volk, S.W. IL-33-Dependent Group 2 Innate Lymphoid Cells Promote Cutaneous Wound Healing. J. Investig. Dermatol. 2016, 136, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.Y.; Han, D.; Sun, X.Y.; Zhang, M.L.; Feng, X.; Sun, C.J.; Gu, J.M.; Tong, C.Y.; Lei, L.C.; Han, W.Y. Mannose-modified chitosan microspheres enhance OprF-OprI-mediated protection of mice against Pseudomonas aeruginosa infection via induction of mucosal immunity. Appl. Microbiol. Biotechnol. 2015, 99, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Mirza, R.; DiPietro, L.A.; Koh, T.J. Selective and specific macrophage ablation is detrimental to wound healing in mice. Am. J. Pathol. 2009, 175, 2454–2462. [Google Scholar] [CrossRef] [PubMed]
- Hetem, D.J.; Vogely, H.C.; Severs, T.T.; Troelstra, A.; Kusters, J.G.; Bonten, M.J. Acquisition of high-level mupirocin resistance in CoNS following nasal decolonization with mupirocin. J. Antimicrob. Chemother. 2015, 70, 1182–1184. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Tu, W.; Xu, C. Mupirocin for preventing exit-site infection and peritonitis in patients undergoing peritoneal dialysis. Nephrol. Dial. Transplant. 2010, 25, 587–592. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.; Moore, Z.E. Interventions for preventing infectious complications in haemodialysis patients with central venous catheters. Cochrane Database Syst. Rev. 2010, 20, CD006894. [Google Scholar] [CrossRef] [PubMed]
- Behm, B.; Babilas, P.; Landthaler, M.; Schreml, S. Cytokines, chemokines and growth factors in wound healing. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 812–820. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Gulbins, E.; Edwards, M.; Caldwell, C.C.; Fraunholz, M.; Becker, K.A. Staphylococcus aureus α-Toxin Induces Inflammatory Cytokines via Lysosomal Acid Sphingomyelinase and Ceramides. Cell Physiol. Biochem. 2017, 43, 2170–2184. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Pelling, J.C. Targeting the PI3K/Akt/mTOR axis by apigenin for cancer prevention. Anti-Cancer Agents Med. Chem. 2013, 13, 971–978. [Google Scholar] [CrossRef]
- Qiu, J.Z.; Luo, M.J.; Wang, J.F.; Dong, J.; Li, H.G.; Leng, B.F.; Zhang, Q.; Dai, X.H.; Zhang, Y.; Niu, X.D.; et al. Isoalantolactone protects against Staphylococcus aureus pneumonia. FEMS Microbiol. Lett. 2011, 324, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Wolf, A.J.; Iliev, I.D.; Berg, B.L.; Underhill, D.M.; Liu, G.Y. Poorly Cross-Linked Peptidoglycan in MRSA Due to mecA Induction Activates the Inflammasome and Exacerbates Immunopathology. Cell Host Microbe 2015, 18, 604–612. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, M.; Zhang, L.; Zhang, H.; Li, X.; Wang, Y.; Xia, F.; Wang, B.; Cai, R.; Guo, Z.; Zhang, Y.; et al. An Ointment Consisting of the Phage Lysin LysGH15 and Apigenin for Decolonization of Methicillin-Resistant Staphylococcus aureus from Skin Wounds. Viruses 2018, 10, 244. https://doi.org/10.3390/v10050244
Cheng M, Zhang L, Zhang H, Li X, Wang Y, Xia F, Wang B, Cai R, Guo Z, Zhang Y, et al. An Ointment Consisting of the Phage Lysin LysGH15 and Apigenin for Decolonization of Methicillin-Resistant Staphylococcus aureus from Skin Wounds. Viruses. 2018; 10(5):244. https://doi.org/10.3390/v10050244
Chicago/Turabian StyleCheng, Mengjun, Lei Zhang, Hao Zhang, Xinwei Li, Yanmei Wang, Feifei Xia, Bin Wang, Ruopeng Cai, Zhimin Guo, Yufeng Zhang, and et al. 2018. "An Ointment Consisting of the Phage Lysin LysGH15 and Apigenin for Decolonization of Methicillin-Resistant Staphylococcus aureus from Skin Wounds" Viruses 10, no. 5: 244. https://doi.org/10.3390/v10050244
APA StyleCheng, M., Zhang, L., Zhang, H., Li, X., Wang, Y., Xia, F., Wang, B., Cai, R., Guo, Z., Zhang, Y., Ji, Y., Sun, C., Feng, X., Lei, L., Yang, Y., Han, W., & Gu, J. (2018). An Ointment Consisting of the Phage Lysin LysGH15 and Apigenin for Decolonization of Methicillin-Resistant Staphylococcus aureus from Skin Wounds. Viruses, 10(5), 244. https://doi.org/10.3390/v10050244