Potential Application of the CRISPR/Cas9 System against Herpesvirus Infections



Abstract
:1. Introduction
1.1. Properties of Herpesviruses
1.2. Life Cycle of Herpesviruses
1.3. Latency and Reactivation of Herpesviruses
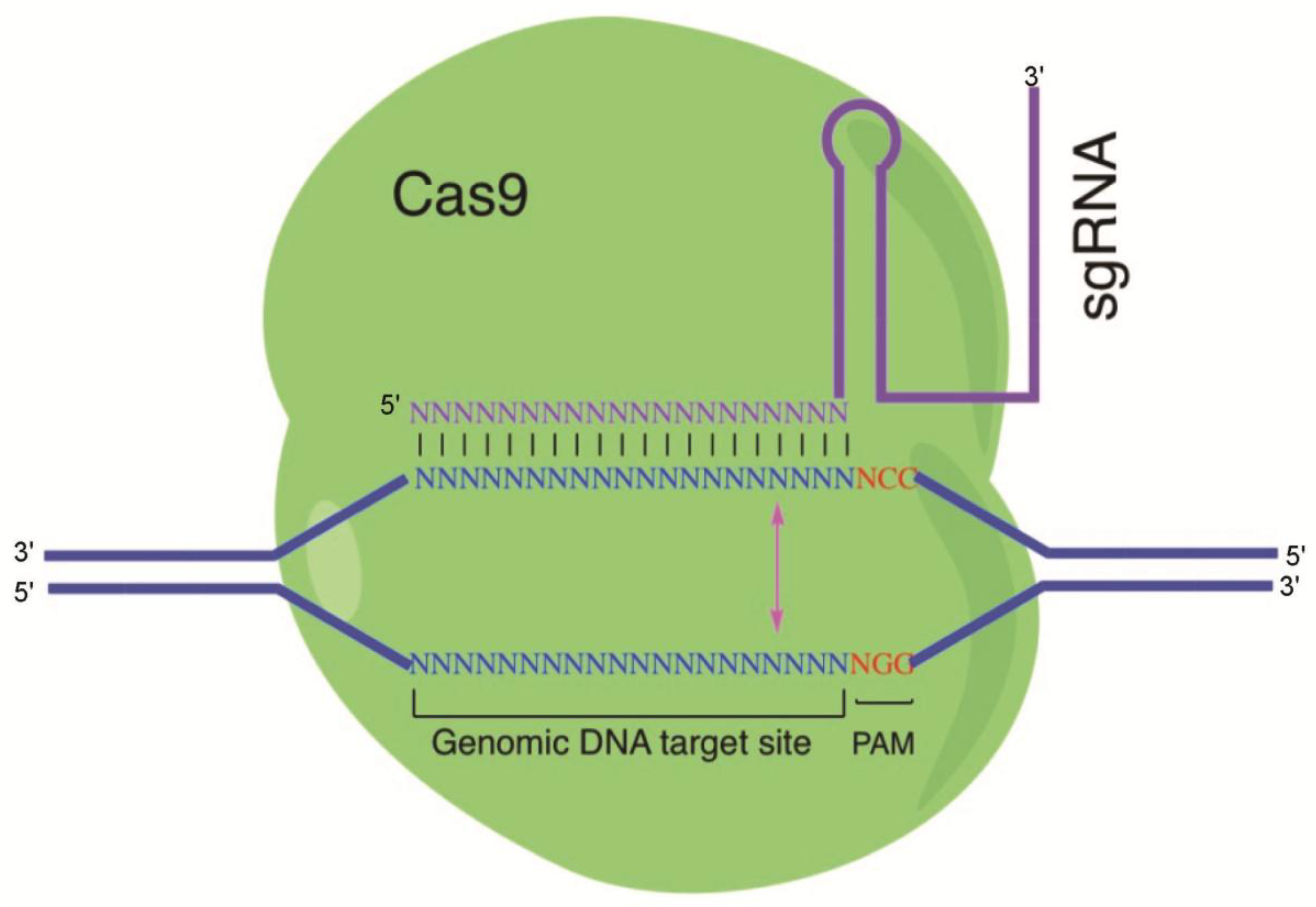
1.4. The Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/Cas9 Nuclease System
2. CRISPR/Cas9 as a Tool for Studying Herpesvirus Host Interaction
2.1. HSV
2.2. EBV
2.3. CMV
2.4. KSHV
3. Treatment of Herpesvirus-Associated Diseases Based on CRISPR/Cas9
4. Challenges of CRISPR/Cas9 Delivery
- The delivery tool is not specific enough: Some delivery tools are not very specific and may deliver nucleic acids to nontarget cells. It is important to reduce the risk of nonspecific delivery, but the evaluation of their benefits and risks is complex.
- The delivery tool is not very efficient: Not all delivery tools are efficient; some of them are low in efficiency and require multiple rounds of transfections. Additionally, it is hard to improve and evaluate their efficiency, especially in animals and clinics.
- The delivery tool is deficient in biosafety: Some delivery tools are toxic, biohazardous, or even destructive to normal cells or recipient hosts. Some delivery tools such as lipoids, viruses, bacteria, and nanoparticles may induce vector-associated immune responses in hosts, and immune barriers must be overcome [36]. Thus, verifying their safety in preliminary testing is needed.
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Pellett, P.E.; Roizman, B. Herpesviridae. In Fields Virology; Lippincott-William & Wilkins: Philadelphia, PA, USA, 2013; pp. 1802–1822. [Google Scholar]
- Charpentier, E.; Doudna, J.A. Biotechnology: Rewriting a genome. Nature 2013, 495, 50–51. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Makarova, K.S.; Zhang, F. Diversity, classification and evolution of CRISPR-Cas systems. Curr. Opin. Microbiol. 2017, 37, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Van Diemen, F.R.; Kruse, E.M.; Hooykaas, M.J.; Bruggeling, C.E.; Schurch, A.C.; van Ham, P.M.; Imhof, S.M.; Nijhuis, M.; Wiertz, E.J.; Lebbink, R.J. CRISPR/Cas9-mediated genome editing of herpesviruses limits productive and latent infections. PLoS Pathog. 2016, 12, e1005701. [Google Scholar] [CrossRef] [PubMed]
- Damania, B.; Cesarman, E. Kaposi’s sarcoma-associated herpesvirus. In Fields Virology; Lippincott-William & Wilkins: Philadelphia, PA, USA, 2013; pp. 2080–2128. [Google Scholar]
- Longnecker, R.; Kieff, E.; Cohen, J.I. Epstein-Barr Virus/Replication and Epstein-Barr Virus. In Fields Virology; Lippincott-William & Wilkins: Philadelphia, PA, USA, 2013; pp. 1898–1959. [Google Scholar]
- Mocarski, E.S.; Shenk, T.; Griffiths, P.D.; Pass, R.F. Cytomegaloviruses. In Fields Virology; Lippincott-William & Wilkins: Philadelphia, PA, USA, 2013; pp. 1960–2014. [Google Scholar]
- Roizman, B.; Knipe, D.M.; Whitley, R.J. Herpes Simplex Viruses. In Fields Virology; Lippincott-William & Wilkins: Philadelphia, PA, USA, 2013; pp. 1823–1897. [Google Scholar]
- Jiang, F.; Doudna, J.A. CRISPR-Cas9 structures and mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef] [PubMed]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Nishimasu, H.; Kodera, N.; Hirano, S.; Ando, T.; Uchihashi, T.; Nureki, O. Real-space and real-time dynamics of CRISPR-Cas9 visualized by high-speed atomic force microscopy. Nat. Commun. 2017, 8, 1430. [Google Scholar] [CrossRef] [PubMed]
- Knight, S.C.; Xie, L.; Deng, W.; Guglielmi, B.; Witkowsky, L.B.; Bosanac, L.; Zhang, E.T.; El Beheiry, M.; Masson, J.B.; Dahan, M.; et al. Dynamics of CRISPR-Cas9 genome interrogation in living cells. Science 2015, 350, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Anders, C.; Niewoehner, O.; Duerst, A.; Jinek, M. Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 2014, 513, 569–773. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, J.E.; Norville, J.E.; Mali, P.; Rios, X.; Aach, J.; Church, G.M. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res. 2013, 41, 4336–4343. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 227–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedland, A.E.; Tzur, Y.B.; Esvelt, K.M.; Colaiacovo, M.P.; Church, G.M.; Calarco, J.A. Heritable genome editing in C. elegans via a CRISPR-Cas9 system. Nat. Methods 2013, 10, 741–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, H.; Shivalila, C.S.; Dawlaty, M.M.; Cheng, A.W.; Zhang, F.; Jaenisch, R. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 2013, 153, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Van Diemen, F.R.; Lebbink, R.J. CRISPR/Cas9, a powerful tool to target human herpesviruses. Cell. Microbiol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Diner, B.A.; Lum, K.K.; Toettcher, J.E.; Cristea, I.M. Viral DNA sensors IFI16 and cyclic GMP-AMP synthase possess distinct functions in regulating viral gene expression, immune defenses, and apoptotic responses during herpesvirus infection. MBio 2016, 7, e01553-16. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.E.; Bottero, V.; Flaherty, S.; Dutta, S.; Singh, V.V.; Chandran, B. IFI16 restricts HSV-1 replication by accumulating on the hsv-1 genome, repressing HSV-1 gene expression, and directly or indirectly modulating histone modifications. PLoS Pathog. 2014, 10, e1004503. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Fan, S.; Zhou, J.; Zhang, Y.; Che, Y.; Cai, H.; Wang, L.; Guo, L.; Liu, L.; Li, Q. The mutated tegument protein UL7 attenuates the virulence of herpes simplex virus 1 by reducing the modulation of alpha-4 gene transcription. Virol. J. 2016, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Furuse, Y.; Oshitani, H.; Kiyono, T. Highly efficient CRISPR/Cas9-mediated cloning and functional characterization of gastric cancer-derived Epstein-Barr virus strains. J. Virol. 2016, 90, 4383–4393. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Xuan, B.; Liu, H.; Zhong, J.; Yu, D.; Qian, Z. Tuberous Sclerosis Complex Protein 2-Independent Activation of mTORC1 by Human Cytomegalovirus pUL38. J. Virol. 2015, 89, 7625–7635. [Google Scholar] [CrossRef] [PubMed]
- Bierle, C.J.; Anderholm, K.M.; Wang, J.B.; McVoy, M.A.; Schleiss, M.R. Targeted mutagenesis of guinea pig cytomegalovirus using CRISPR/Cas9-mediated gene editing. J. Virol. 2016, 90, 6989–6998. [Google Scholar] [CrossRef] [PubMed]
- Avey, D.; Tepper, S.; Li, W.; Turpin, Z.; Zhu, F. Phosphoproteomic Analysis of KSHV-Infected Cells Reveals Roles of ORF45-Activated RSK during Lytic Replication. PLoS Pathog. 2015, 11, e1004993. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Sun, L.; Yu, T.; Pan, Y.; Wang, D.; Hu, X.; Fu, Z.; He, Q.; Cao, G. A CRISPR/Cas9 and Cre/Lox system-based express vaccine development strategy against re-emerging Pseudorabies virus. Sci. Rep. 2016, 6, 19176. [Google Scholar] [CrossRef] [PubMed]
- Moos, W.H.; Pinkert, C.A.; Irwin, M.H.; Faller, D.V.; Kodukula, K.; Glavas, I.P.; Steliou, K. Epigenetic treatment of persistent viral Infections. Drug Dev. Res. 2017, 78, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Pennington, M.R.; Van de Walle, G.R. Electric cell-substrateiImpedance sensing to monitor viral growth and study cellular responses to infection with alphaherpesviruses in real time. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Roehm, P.C.; Shekarabi, M.; Wollebo, H.S.; Bellizzi, A.; He, L.; Salkind, J.; Khalili, K. Inhibition of HSV-1 replication by gene editing strategy. Sci. Rep. 2016, 6, 23146. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Quake, S.R. RNA-guided endonuclease provides a therapeutic strategy to cure latent herpesviridae infection. Proc. Natl. Acad. Sci. USA 2014, 111, 13157–13162. [Google Scholar] [CrossRef] [PubMed]
- Yuen, K.S.; Wang, Z.M.; Wong, N.M.; Zhang, Z.Q.; Cheng, T.F.; Lui, W.Y.; Chan, C.P.; Jin, D.Y. Suppression of Epstein-Barr virus DNA load in latently infected nasopharyngeal carcinoma cells by CRISPR/Cas9. Virus Res. 2018, 244, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.R.; Yang, H.C.; Kuo, Y.T.; Liu, C.J.; Yang, T.Y.; Sung, K.C.; Lin, Y.Y.; Wang, H.Y.; Wang, C.C.; Shen, Y.C.; et al. The CRISPR/Cas9 system facilitates clearance of the intrahepatic HBV templates in vivo. Mol. Ther. Nucleic Acids 2014, 19, e186. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Zhang, T.; Qu, X.; Zhang, Y.; Putatunda, R.; Xiao, X.; Li, F.; Xiao, W.; Zhao, H.; Dai, S.; et al. In vivo excision of HIV-1 provirus by saCas9 and multiplex single-guide RNAs in animal models. Mol. Ther. 2017, 25, 1168–1186. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Menezes, M.E.; Bhatia, S.; Wang, X.Y.; Emdad, L.; Sarkar, D.; Fisher, P.B. Gene therapies for cancer: Strategies, challenges and successes. J. Cell. Physiol. 2015, 230, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, G.A.R.; Paiva, R.M.A. Gene therapy: Advances, challenges and perspectives. Einstein (Sao Paulo) 2017, 15, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Gong, H.; Li, H.; Vu, G.P.; Lu, S.; Liu, F. Oral delivery of RNase P ribozymes by Salmonella inhibits viral infection in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 3222–3227. [Google Scholar] [CrossRef] [PubMed]
- Gan, W.B.; Grutzendler, J.; Wong, W.T.; Wong, R.O.; Lichtman, J.W. Multicolor “DiOlistic” labeling of the nervous system using lipophilic dye combinations. Neuron 2000, 27, 219–225. [Google Scholar] [CrossRef]
- Guo, W.; Guo, Y.; Tang, S.; Qu, H.; Zhao, H. Dendritic cell-Ewing’s sarcoma cell hybrids enhance antitumor immunity. Clin. Orthop. Relat. Res. 2008, 466, 2176–2183. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.P.; Sharei, A.; Ding, X.; Sahay, G.; Langer, R.; Jensen, K.F. In vitro and ex vivo strategies for intracellular delivery. Nature 2016, 538, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Oggu, G.S.; Sasikumar, S.; Reddy, N.; Ella, K.K.R.; Rao, C.M.; Bokara, K.K. Gene delivery approaches for mesenchymal stem cell therapy: Strategies to increase efficiency and specificity. Stem. Cell Rev. 2017, 13, 725–740. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Min, T.; Ma, S.; Hu, L.; Chen, H.; Lu, D. Intracellular generation of single-strand template increases the knock-in efficiency by combining CRISPR/Cas9 with AAV. Mol. Genet. Genom. 2018; in press. [Google Scholar]
- Finn, J.D.; Smith, A.R.; Patel, M.C.; Shaw, L.; Youniss, M.R.; van Heteren, J.; Dirstine, T.; Ciullo, C.; Lescarbeau, R.; Seitzer, J.; et al. A single administration of CRISPR/Cas9 lipid nanoparticles achieves robust and persistent in vivo genome editing. Cell Rep. 2018, 22, 2227–2235. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.H.; Suh, Y. In vivo genome editing in animals using AAV-CRISPR system: Applications to translational research of human disease. F1000Res 2017, 6, 2153. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type (Synonym) | Subfamily | Primary Target Cells | Latency Cells | Pathophysiology |
|---|---|---|---|---|
| HHV-1 (HSV-1) | α (alpha) | Mucoepithelial cells | Sensory neurons | Oral or genital herpes (predominantly orofacial), cold sores, keratitis, etc. |
| HHV-2 (HSV-2) | α (alpha) | Mucoepithelial cells | Sensory neurons | Oral or genital herpes (predominantly genital), etc. |
| HHV-4 (EBV) | γ (gamma) | B cells, Epithelial cells | B cells, Epithelial cells | Infectious mononucleosis, Burkitt’s lymphoma, nasopharyngeal carcinoma, Hodgkin’s disease, post-transplant lymphomas, CNS lymphoma in AIDS patients, etc. |
| HHV-5 (HCMV) | β (beta) | Monocytes, Lymphocytes, Epithelial cells | Peripheral monocytes, CD34+ progenitor cells | Infectious mononucleosis-like syndrome, retinitis, pneumonitis, gastrointestinal diseases, mental retardation, vascular disorders, etc. |
| HHV-8 (KSHV) | γ | Lymphocytes and other cells | B cells, Mononucleocytes | Kaposi’s sarcoma, primary effusion lymphoma, some types of multicentric Castleman’s disease, etc. |
| Delivery Tools | Example | Characteristic |
|---|---|---|
| Lipoid | Lipofectamine, Liposome | The lipid subunits which form liposomes entrap the transfection materials, allowing themselves to overcome the electrostatic repulsion of the cell membrane to let DNA or RNA cross into the cytoplasm to access the nuclei or organelles. |
| Virus | Lentivirus, Adenovirus, Adeno-associated virus (AAV), Baculovirus | A specific virus is engineered to deliver DNA or RNA to target cells and used as a vector for gene transfer. |
| Nanoparticle | Mesoporous silica nanoparticles (MSNs), Dendrimers, Carbon Nanotubes, Cationic polymers | Nanoparticles (1–100 nanometers in size), consist of a variety of compounds and materials, can be complexed with DNA or RNA for gene delivery. |
| Bacterium | Salmonella | An attenuated strain of Salmonella which is invasive but nonpathogenic shows DNA transfer activity with little cytotoxicity and pathogenicity in hosts. |
| Gene gun | PDS-1000/He Particle Delivery System | The device, a biolistic particle delivery system, is used for delivering exogenous DNA to cells; the payload is an elemental particle of a heavy metal coated with DNA. |
| Electroporation | Electroporator | An electric field is applied to cells to increase the cell permeability, allowing DNA to be introduced into the cell. |
| Nanostraw | Navan | The device is used for creating a direct physical conduit to cells for DNA delivery, mimicking the gap junction between cells. |
| Challenge | Strategy |
|---|---|
| Specificity | Discovery of a specific virus such as adeno-associated viruses (AAV). |
| Efficiency | Application of a combination system such as AAV-CRISPR. |
| Biosafety | Combination with several factors such as smaller Cas9 orthologues, tissue-specific minimal promoters, AAV serotypes, and different routes of administration; Development of novel and safe delivery tools such as lipid nanoparticles (LNP), AAV, and baculoviruses. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Sheng, J.; Trang, P.; Liu, F. Potential Application of the CRISPR/Cas9 System against Herpesvirus Infections. Viruses 2018, 10, 291. https://doi.org/10.3390/v10060291
Chen Y-C, Sheng J, Trang P, Liu F. Potential Application of the CRISPR/Cas9 System against Herpesvirus Infections. Viruses. 2018; 10(6):291. https://doi.org/10.3390/v10060291
Chicago/Turabian StyleChen, Yuan-Chuan, Jingxue Sheng, Phong Trang, and Fenyong Liu. 2018. "Potential Application of the CRISPR/Cas9 System against Herpesvirus Infections" Viruses 10, no. 6: 291. https://doi.org/10.3390/v10060291
APA StyleChen, Y. -C., Sheng, J., Trang, P., & Liu, F. (2018). Potential Application of the CRISPR/Cas9 System against Herpesvirus Infections. Viruses, 10(6), 291. https://doi.org/10.3390/v10060291