Identification and Characterization of Type IV Pili as the Cellular Receptor of Broad Host Range Stenotrophomonas maltophilia Bacteriophages DLP1 and DLP2



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Phage and Growth Conditions
2.2. Transposon Mutant Library Receptor Screen
2.3. Phage Plaquing Assays
2.4. Construction of ΔpilA S. maltophilia D1585 and 280 Mutants
2.5. Complementation of Pilus Mutants
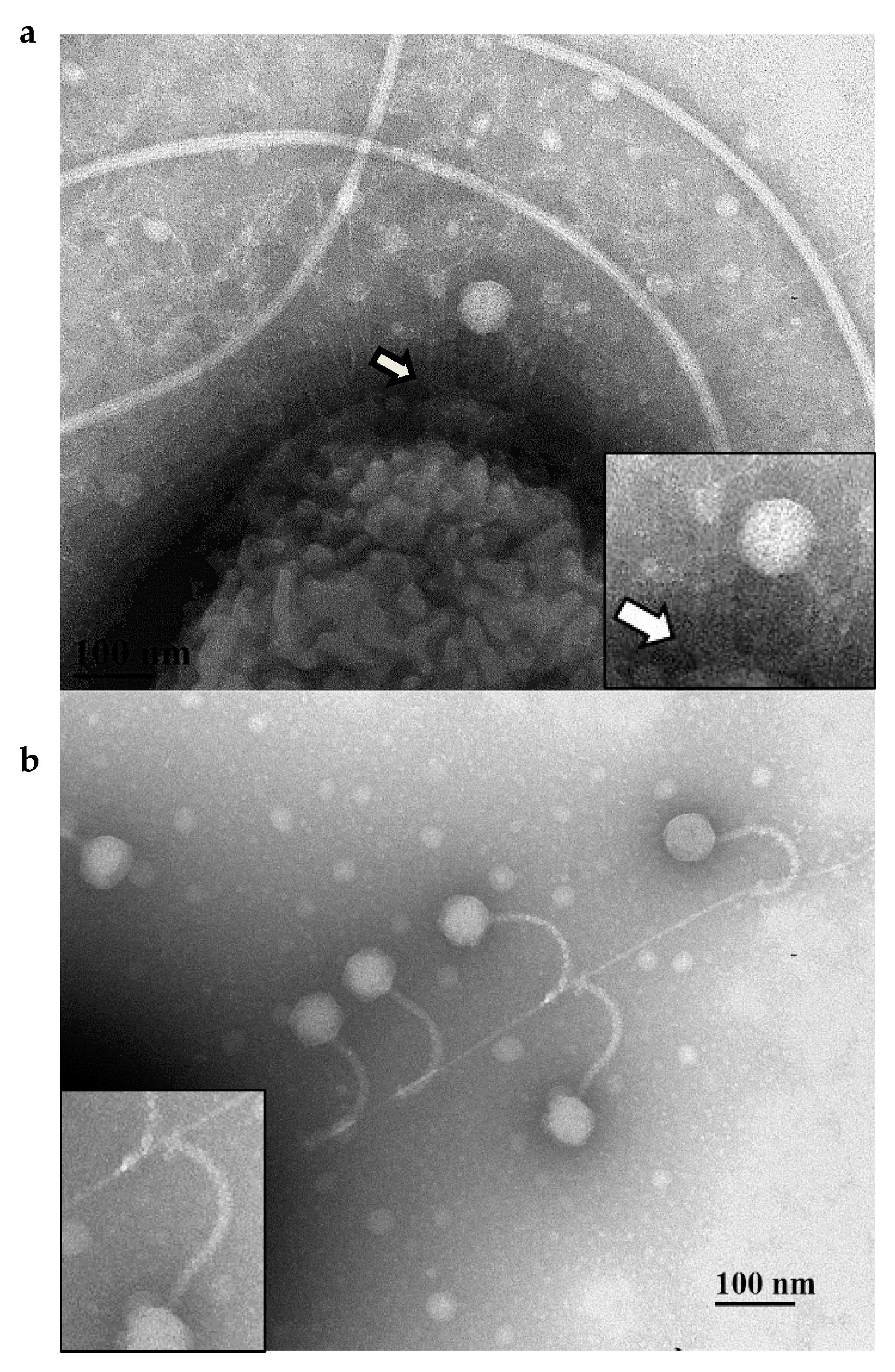
2.6. Transmission Electron Microscopy
2.7. Twitching Motility Assay
2.8. Bioinformatic Analysis
3. Results and Discussion
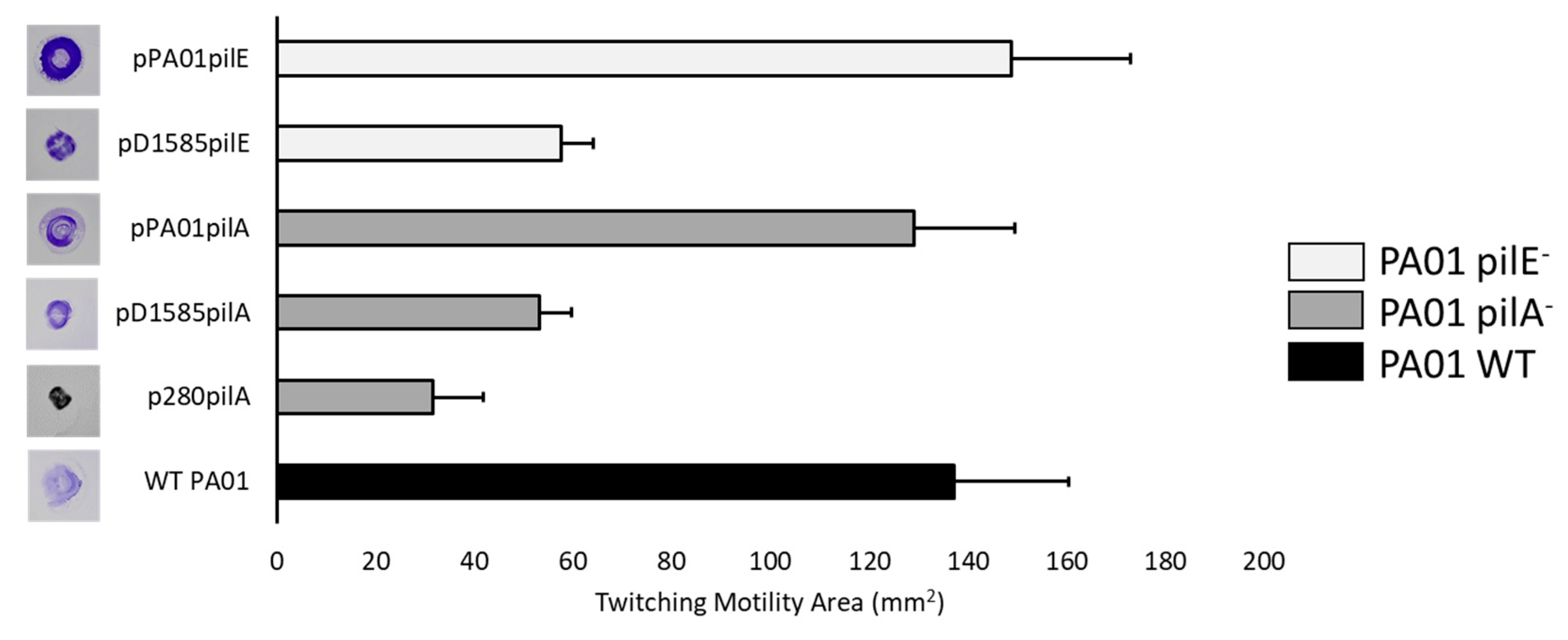
3.1. P. aeruginosa PA01 Type IV Pilus Mutants Are Resistant to DLP1 Infection
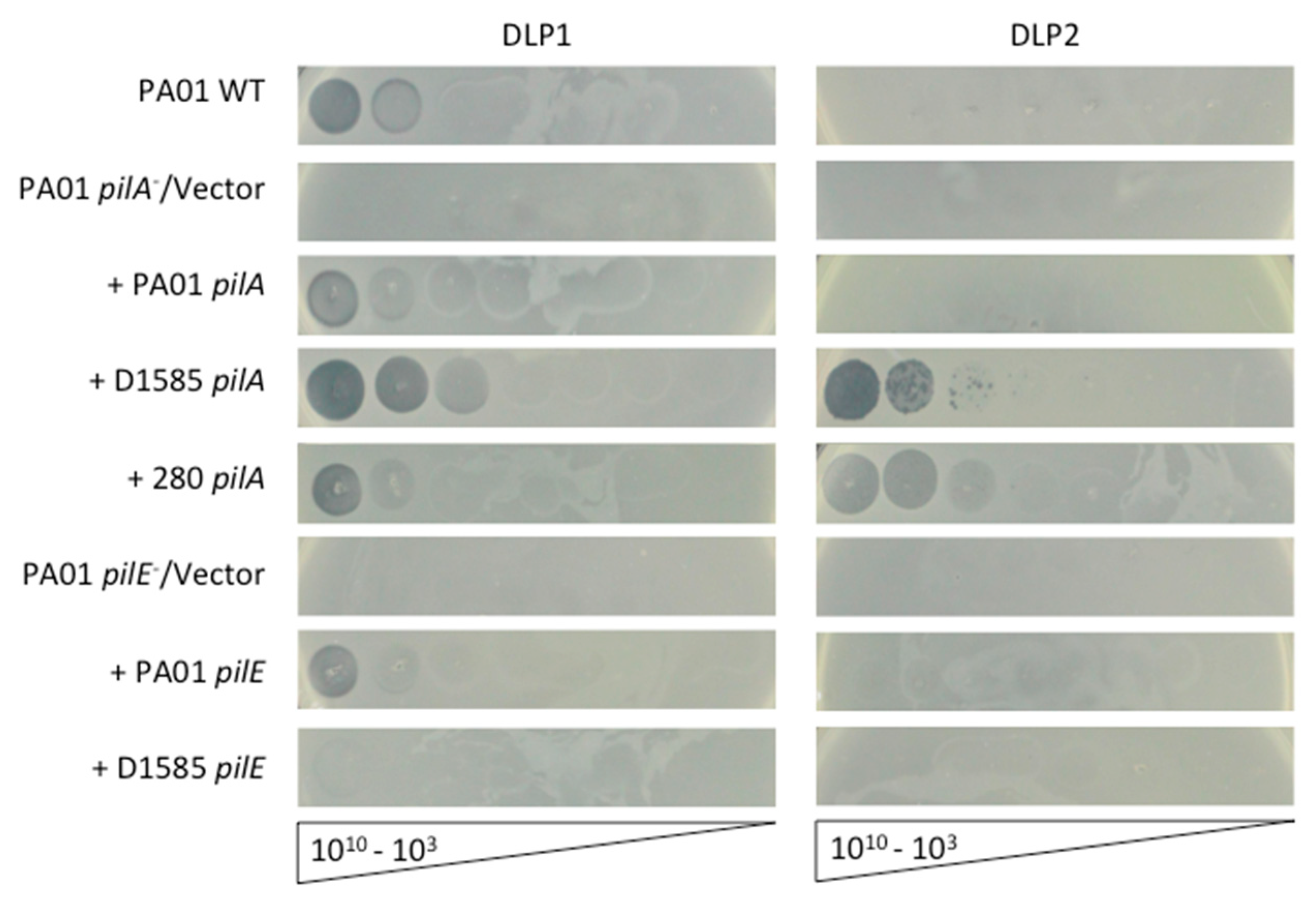
3.2. Complementation in P. aeruginosa Restores DLP1 Infectivity
3.3. Deletion of pilA in S. maltophilia D1585 Prevents DLP1 and DLP2 Infection
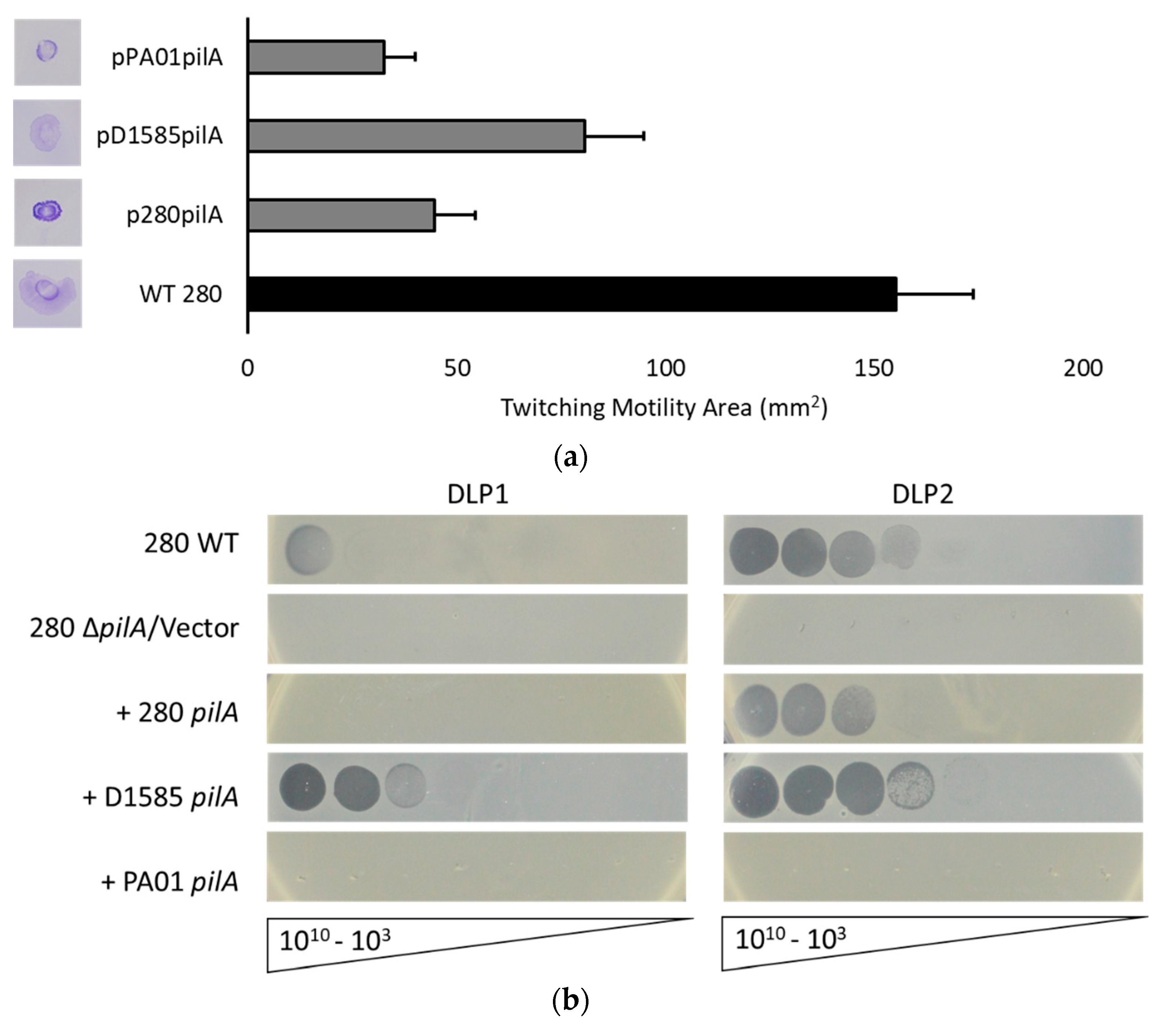
3.4. Deletion of pilA in S. maltophilia 280 Prevents DLP2 Infection
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. The Review on Antimicrobial Resistance. Available online: https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 7 May 2018).
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Martinez, J.L. Friends or foes: Can we make a distinction between beneficial and harmful strains of the Stenotrophomonas maltophilia complex? Front. Microbiol. 2015, 6, 241. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Monchy, S.; Cardinale, M.; Taghavi, S.; Crossman, L.; Avison, M.B.; Berg, G.; van der Lelie, D.; Dow, J.M. The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 2009, 7, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Pabary, R.; Singh, C.; Morales, S.; Bush, A.; Alshafi, K.; Bilton, D.; Alton, E.W.F.W.; Smithyman, A.; Davies, J.C. Antipseudomonal bacteriophage reduces infective burden and inflammatory response in murine lung. Antimicrob. Agents Chemother. 2016, 60, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Vilas Boas, D.; Sillankorva, S.; Azeredo, J. Phage therapy: A step forward in the treatment of Pseudomonas aeruginosa infections. J. Virol. 2015, 89, 7449–7456. [Google Scholar] [CrossRef] [PubMed]
- Pompilio, A.; Pomponio, S.; Crocetta, V.; Gherardi, G.; Verginelli, F.; Fiscarelli, E.; Dicuonzo, G.; Savini, V.; D’Antonio, D.; Di Bonaventura, G. Phenotypic and genotypic characterization of Stenotrophomonas maltophilia isolates from patients with cystic fibrosis: Genome diversity, biofilm formation, and virulence. BMC Microbiol. 2011, 11, 159. [Google Scholar] [CrossRef] [PubMed]
- McVay, C.S.; Velásquez, M.; Fralick, J.A. Phage therapy of Pseudomonas aeruginosa in a mouse burn wound model. Antimicrob. Agents Chemother. 2007, 51, 1934–1938. [Google Scholar] [CrossRef] [PubMed]
- Seed, K.D.; Dennis, J.J. Experimental bacteriophage therapy increases survival of Galleria mellonella larvae infected with clinically relevant strains of the Burkholderia cepacia complex. Antimicrob. Agents Chemother. 2009, 53, 2205–2208. [Google Scholar] [CrossRef] [PubMed]
- Semler, D.D.; Goudie, A.D.; Finlay, W.H.; Dennis, J.J. Aerosol phage therapy efficacy in Burkholderia cepacia complex respiratory infections. Antimicrob. Agents Chemother. 2014, 58, 4005–4013. [Google Scholar] [CrossRef] [PubMed]
- Merabishvili, M.; Pirnay, J.P.; Verbeken, G.; Chanishvili, N.; Tediashvili, M.; Lashkhi, N.; Glonti, T.; Krylov, V.; Mast, J.; Van Parys, L.; et al. Quality-controlled small-scale production of a well-defined bacteriophage cocktail for use in human clinical trials. PLoS ONE 2009, 4, e4944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, A.; Hawkins, C.H.; Änggård, E.E.; Harper, D.R. A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa; A preliminary report of efficacy. Clin. Otolaryngol. 2009, 34, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Vandenheuvel, D.; Lavigne, R.; Brüssow, H. Bacteriophage therapy: Advances in formulation strategies and human clinical trials. Annu. Rev. Virol. 2015, 2, 599–618. [Google Scholar] [CrossRef] [PubMed]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.K.; Turner, P.E.; Kim, S.; Mojibian, H.R.; Elefteriades, J.A.; Narayan, D. Phage treatment of an aortic graft infected with Pseudomonas aeruginosa. Evol. Med. Public Health 2018, 2018, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Leon, M.; Bastias, R. Virulence reduction in bacteriophage resistant bacteria. Front. Microbiol. 2015, 6, 343. [Google Scholar] [CrossRef] [PubMed]
- Peters, D.L.; Lynch, K.H.; Stothard, P.; Dennis, J.J. The isolation and characterization of two Stenotrophomonas maltophilia bacteriophages capable of cross-taxonomic order infectivity. BMC Genom. 2015, 16, 664. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; Kuhl, S.J.; Blasdel, B.G.; Kutter, E.M. Phage treatment of human infections. Bacteriophage 2011, 1, 66–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, D.L.; Stothard, P.; Dennis, J.J. The isolation and characterization of Stenotrophomonas maltophilia T4-like bacteriophage DLP6. PLoS ONE 2017, 12, e0173341. [Google Scholar] [CrossRef] [PubMed]
- Lewenza, S.; Falsafi, R.K.; Winsor, G.; Gooderham, W.J.; Mcphee, J.B.; Brinkman, F.S.L.; Hancock, R.E.W. Construction of a mini-Tn5-luxCDABE mutant library in Pseudomonas aeruginosa PAO1: A tool for identifying differentially regulated genes. Genome Res. 2005, 15, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, M.A.; Alwood, A.; Thaipisuttikul, I.; Spencer, D.; Haugen, E.; Ernst, S.; Will, O.; Kaul, R.; Raymond, C.; Levy, R.; et al. Comprehensive transposon mutant library of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2003, 100, 14339–14344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Held, K.; Ramage, E.; Jacobs, M.; Gallagher, L.; Manoil, C. Sequence-verified two-allele transposon mutant library for Pseudomonas aeruginosa PAO1. J. Bacteriol. 2012, 194, 6387–6389. [Google Scholar] [CrossRef] [PubMed]
- Krylov, V.N.; Zhazykov, I. Pseudomonas bacteriophage phiKZ—Possible model for studying the genetic control of morphogenesis. Genetika 1978, 14, 678–685. [Google Scholar] [PubMed]
- Kropinski, A.M.; Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of bacteriophages by double agar overlay plaque assay. In Bacteriophages: Methods and Protocols, Volume 1: Isolation, Charcterization, and Interactions; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: New Delhi, India, 2009; pp. 69–76. [Google Scholar]
- Holloway, B.W. Genetic Recombination in Pseudomonas aeruginosa. J. Gen. Microbiol. 1955, 13, 572–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, R.; Priefer, U.; Puhler, A. A broad host range mobilization system for in vivo genetic engineering: Transposon mutagenesis in gram negative bacteria. Nat. Biotechnol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Hanahan, D.; Jessee, J.; Bloom, F.R. Plasmid transformation of Escherichia coli and other bacteria. Methods Enzymol. 1991, 204, 63–113. [Google Scholar] [PubMed]
- Mesyanzhinov, V.V.; Robben, J.; Grymonprez, B.; Kostyuchenko, V.A.; Bourkaltseva, M.V.; Sykilinda, N.N.; Krylov, V.N.; Volckaert, G. The genome of bacteriophage phiKZ of Pseudomonas aeruginosa. J. Mol. Biol. 2002, 317, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Phillips, R.W.; Elzer, P.H.; Roop, R.M.; Peterson, K.M. pBBR1MCS: A broad-host-range cloning vector. Biotechniques 1994, 16, 800–802. [Google Scholar] [PubMed]
- West, S.E.H.; Schweizer, H.P.; Dall, C.; Sample, A.K.; Runyen-Janecky, L.J. Construction of improved Escherichia-Pseudomonas shuttle vectors derived from pUC18/19 and sequence of the region required for their replication in Pseudomonas aeruginosa. Gene 1994, 128, 81–86. [Google Scholar] [CrossRef]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: Application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinform. Appl. Note 2012, 19, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Chojnacki, S.; Cowley, A.; Lee, J.; Foix, A.; Lopez, R. Programmatic access to bioinformatics tools from EMBL-EBI update: 2017. Nucleic Acids Res. 2017, 45, W550–W553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hmelo, L.R.; Borlee, B.R.; Almblad, H.; Love, M.E.; Randall, T.E.; Tseng, B.S.; Lin, C.; Irie, Y.; Storek, K.M.; Yang, J.J.; et al. Precision-engineering the Pseudomonas aeruginosa genome with two-step allelic exchange. Nat. Protoc. 2015, 10, 1820–1841. [Google Scholar] [CrossRef] [PubMed]
- Heckman, K.L.; Pease, L.R. Gene splicing and mutagenesis by PCR-driven overlap extension. Nat. Protoc. 2007, 2, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Poole, K.; Heinrichs, D.E.; Neshat, S. Cloning and sequence analysis of an EnvCD homologue in Pseudomonas aeruginosa: Regulation by iron and possible involvement in the secretion of the siderophore pyoverdine. Mol. Microbiol. 1993, 10, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.H.; Kumar, A.; Schweizer, H.P. A 10-min method for preparation of highly electrocompetent Pseudomonas aeruginosa cells: Application for DNA fragment transfer between chromosomes and plasmid transformation. J. Microbiol. Methods 2006, 64, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Dong, H.; Huang, Y.P. Highly efficient transformation of Stenotrophomonas maltophilia S21, an environmental isolate from soil, by electroporation. J. Microbiol. Methods 2014, 107, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, L.; Whitchurch, C.B. Motility assay: Twitching motility. In Pseudomonas Methods and Protocols; Filloux, A., Ramos, J.-L., Eds.; Humana Press: New York, NY, USA, 2014; Volume 1149, pp. 73–86. ISBN 978-1-4939-0472-3. [Google Scholar]
- Pompilio, A.; Crocetta, V.; Ghosh, D.; Chakrabarti, M.; Gherardi, G.; Vitali, L.A.; Fiscarelli, E.; Di Bonaventura, G. Stenotrophomonas maltophilia phenotypic and genotypic diversity during a 10-year colonization in the lungs of a cystic fibrosis patient. Front. Microbiol. 2016, 7, 1551. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed]
- Burrows, L.L. Pseudomonas aeruginosa twitching motility: Type IV pili in action. Annu. Rev. Microbiol. 2012, 66, 493–520. [Google Scholar] [CrossRef] [PubMed]
- Whitchurch, C.B.; Mattick, J.S. Characterization of a gene, pilU, required for twitching motility but not phage sensitivity in Pseudomonas aeruginosa. Mol. Microbiol. 1994, 13, 1079–1091. [Google Scholar] [CrossRef] [PubMed]
- Burrows, L.L. Weapons of mass retraction. Mol. Microbiol. 2005, 57, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, Y.; Sugiman-Marangos, S.; Harvey, H.; Bell, S.D.; Charlton, C.L.; Junop, M.S.; Burrows, L.L. Pseudomonas aeruginosa minor pilins prime type IVa pilus assembly and promote surface display of the PilY1 adhesin. J. Biol. Chem. 2015, 290, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Kus, J.V.; Tullis, E.; Cvitkovitch, D.G.; Burrows, L.L. Significant differences in type IV pilin allele distribution among Pseudomonas aeruginosa isolates from cystic fibrosis (CF) versus non-CF patients. Microbiology 2004, 150, 1315–1326. [Google Scholar] [CrossRef] [PubMed]
- Giltner, C.L.; Rana, N.; Lunardo, M.N.; Hussain, A.Q.; Burrows, L.L. Evolutionary and functional diversity of the Pseudomonas type IVa pilin island. Environ. Microbiol. 2011, 13, 250–264. [Google Scholar] [CrossRef] [PubMed]
- Elleman, T.C.; Hoyne, P.A.; Stewart, D.J.; Mckern, N.M.; Peterson, J.E. Expression of pili from Bacteroides nodosus in Pseudomonas aeruginosa. J. Bacteriol. 1986, 168, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Beard, M.K.M.; Mattick, J.S.; Moore, L.J.; Mott, M.R.; Marrs, C.F.; Egerton, J.R. Morphogenetic expression of Moraxella bovis fimbriae (pili) in Pseudomonas aeruginosa. J. Bacteriol. 1990, 172, 2601–2607. [Google Scholar] [CrossRef] [PubMed]
- Hoyne, P.A.; Haas, R.; Meyer, T.F.; Davies, J.K.; Elleman, T.C. Production of Neisseria gonorrhoeae pili (fimbriae) in Pseudomonas aeruginosa. J. Bacteriol. 1992, 174, 7321–7327. [Google Scholar] [CrossRef] [PubMed]
- Sauvonnet, N.; Gounon, P.; Pugsley, A.P. PpdD type IV pilin of Escherichia coli K-12 can be assembled into pili in Pseudomonas aeruginosa. J. Bacteriol. 2000, 182, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Aas, F.E.; Wolfgang, M.; Frye, S.; Dunham, S.; Løvold, C.; Koomey, M. Competence for natural transformation in Neisseria gonorrhoeae: Components of DNA binding and uptake linked to type IV pilus expression. Mol. Microbiol. 2002, 46, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Roine, E.; Raineri, D.M.; Romantschuk, M.; Wilson, M.; Nunn, D.N. Characterization of type IV pilus genes in Pseudomonas syringae pv. tomato DC3000. Mol. Plant-Microbe Interact. 1998, 11, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.A.; Mattick, J.S.; Aim, R.A. Functional expression of heterologous type 4 fimbriae in Pseudomonas aeruginosa. Gene 1996, 175, 143–150. [Google Scholar] [CrossRef]
- Winther-Larsen, H.C.; Wolfgang, M.C.; Van Putten, J.P.M.; Roos, N.; Aas, F.E.; Egge-Jacobsen, W.M.; Maier, B.; Koomey, M. Pseudomonas aeruginosa type IV pilus expression in Neisseria gonorrhoeae: Effects of pilin subunit composition on function and organelle dynamics. J. Bacteriol. 2007, 189, 6676–6685. [Google Scholar] [CrossRef] [PubMed]
- Haley, C.L.; Kruczek, C.; Qaisar, U.; Colmer-Hamood, J.A.; Hamood, A.N. Mucin inhibits Pseudomonas aeruginosa biofilm formation by significantly enhancing twitching motility. Can. J. Microbiol. 2014, 60, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Harvey, H.; Bondy-Denomy, J.; Marquis, H.; Sztanko, K.M.; Davidson, A.R.; Burrows, L.L. Pseudomonas aeruginosa defends against phages through type IV pilus glycosylation. Nat. Microbiol. 2018, 3, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A.R.; Cardarelli, L.; Pell, L.G.; Radford, D.R.; Maxwell, K.L. Long Noncontractile Tail Machines of Bacteriophages. In Viral Molecular Machines; Rossman, M.G., Rao, V.B., Eds.; Springer: Boston, MA, USA, 2012; pp. 115–142. [Google Scholar]
- Watanabe, K.; Ishibashi, K.; Nakashima, Y.; Sakurai, T. A phage-resistant mutant of Lactobacillus casei which permits phage adsorption but not genome injection. J. Gen. Virol. 1984, 65, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Guttman, B.; Raya, R.; Kutter, E.M. Basic Phage Biology. In Bacteriophages: Biology and Applications; Kutter, E., Sulakvelidze, A., Eds.; CRC Press: Boca Raton, FL, USA, 2005; ISBN 0849313368. [Google Scholar]
- Bradley, D.E. A pilus-dependent Pseudomonas aeruginosa bacteriophage with a long noncontractile tail. Virology 1973, 51, 489–492. [Google Scholar] [CrossRef]
- Pemberton, J.M. F116: A DNA bacteriophage specific for the pili of Pseudomonas aeruginosa strain PAO. Virology 1973, 55, 558–560. [Google Scholar] [CrossRef]
- Budzik, J.M.; Rosche, W.A.; Rietsch, A.; O’toole, G.A. Isolation and characterization of a generalized transducing phage for Pseudomonas aeruginosa strains PAO1 and PA14. J. Bacteriol. 2004, 186, 3270–3273. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.-J.; Chung, I.-Y.; Choi, K.B.; Lau, G.W.; Cho, Y.-H. Genome sequence comparison and superinfection between two related Pseudomonas aeruginosa phages, D3112 and MP22. Microbiology 2007, 153, 2885–2895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.-W.; Cho, Y.-H. Complete genome sequence of Pseudomonas aeruginosa podophage MPK7, which requires type IV pili for infection. Genome Announc. 2013, 1, e00744-13. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. 2010, 9, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Genotype or Description | Source |
|---|---|---|
| P. aeruginosa PA01 | Wildtype host for DLP1 | [25] |
| S. maltophilia D1585 | Wildtype host for DLP1 and DLP2 | CBCCRRR * |
| D1585 ΔpilA | Clean deletion of pilA in D1585 | This study |
| S. maltophilia 280 | Wildtype host for DLP2 | PLPHN/AHS ** |
| 280 ΔpilA | Clean deletion of pilA in 280 | This study |
| E. coli S17-1 | Conjugative donor strain | [26] |
| E. coli DH5α | Host for plasmid cloning | [27] |
| Phage | ||
| DLP1 | Lytic phage | Accession: KR537872.1 [17] |
| DLP2 | Lytic phage | Accession: KR537871.1 [17] |
| φKZ | Lytic Pseudomonas phage | Accession: NC_004629.1 [23,28] |
| Plasmids | ||
| pBBR1MCS | Broad-host range cloning vector, CmR | [29] |
| pD1585pilA | pBBR1MCS carrying D1585 pilA, CmR | This study |
| pPA01pilA | pBBR1MCS carrying PA01 pilA, CmR | This study |
| p280pilA | pBBR1MCS carrying 280 pilA, GmR | This study |
| pD1585pilE | pBBR1MCS carrying D1585 pilE, CmR | This study |
| pPA01pilE | pBBR1MCS carrying PA01 pilE, CmR | This study |
| pUCP22 | Broad-host range cloning vector, GmR | [30] |
| pUCP(D1585pilA) | pUCP22 carrying D1585 pilA, GmR | This study |
| pUCP(PA01pilA) | pUCP22 carrying PA01 pilA, GmR | This study |
| pUCP(280pilA) | pUCP22 carrying 280 pilA, GmR | This study |
| pUCP(D1585pilE) | pUCP22 carrying D1585 pilE, GmR | This study |
| pUCP(PA01pilE) | pUCP22 carrying PA01 pilE, GmR | This study |
| pEX18Tc | TcR, oriT, sacB, gene replacement vector | [31] |
| pD1585ΔpilA | pEX18Tc, 2 kb ΔpilA D1585 region | This study |
| p280ΔpilA | pEX18Tc, 2 kb ΔpilA 280 region | This study |
| Number of Mutants | Gene Affected | Function | DLP1 Lysis | Source |
|---|---|---|---|---|
| 2 | pilA | Major pilin subunit | − | [21] |
| 4 | pilB | Cytoplasmic ATPase/pilin polymerase | − | [20] |
| 3 | pilE | Minor pilin subunit | − | [20] |
| 1 | pilF | Outer membrane pilotin; controls secretin localization | − | [21] |
| 1 | pilJ | Involved in pilus assembly | − | [20] |
| 1 | pilN | Inner membrane assembly protein | − | [21] |
| 2 | pilQ | Secretin monomer; forms outer membrane pore | − | [21] |
| 1 | pilR | Cytoplasmic response regulator of two-component system; regulates PilA expression | − | [20] |
| 4 | pilS | Inner membrane histidine kinase of two component system; regulates PilA expression | − | [20] |
| 2 | pilT | Cytoplasmic ATPase; pilin depolymerase | − | [20] |
| 1 | pilU | Cytoplasmic ATPase; regulation of pilus retraction | + | [21] |
| 2 | pilV | Minor pilin subunit | − | [20] |
| 4 | pilY1 | Possible adhesin; regulates pilus retraction | − | [20] |
| 3 | fimV | Inner membrane protein; aids in secretin assembly | +/− | [20] |
| 1 | algR | Regulates expression of minor pilin operon | +/− | [20] |
| 1 | PA2806 | Conserved hypothetical protein | − | [20] |
| A. Strain + DLP1 | Pilin Complement | ||
|---|---|---|---|
| pPA01pilA | pD1585pilA | p280pilA | |
| P. aeruginosa PA01 pilA− | 109 | 108 | 109 |
| S. maltophilia D1585ΔpilA | 107 | 103 | 105 |
| S. maltophilia 280ΔpilA | - | 108 | - |
| B. Strain + DLP2 | pPA01pilA | pD1585pilA | p280pilA |
| P. aeruginosa PA01 pilA− | - | 107 | 108 |
| S. maltophilia D1585ΔpilA | 109 | 103 | 103 |
| S. maltophilia 280ΔpilA | - | 105 | 107 |
 clearing at 109,
clearing at 109,  plaquing at 109,
plaquing at 109,  clearing at 108,
clearing at 108,  plaquing at 107,
plaquing at 107,  plaquing at 105, ◼ plaquing at 103.
plaquing at 105, ◼ plaquing at 103.| Bacteriophage | Accession | % Homology to DLP1 Pfam13550 | % Homology to DLP2 Pfam13550 |
|---|---|---|---|
| Stenotrophomonas phage DLP1 a | AKI28788.1 | - | 98.3 |
| Pseudomonas phage 73 | YP_001293432 | 99.5 | 98.5 |
| Pseudomonas phage vB_PaeS_C1 | AVJ48095 | 98.8 | 99.3 |
| Pseudomonas phage vB_Pae-Kakheti25 | YP_006299890 | 98.5 | 99.1 |
| Pseudomonas phage vB_Pae_PS9N | AIW01689 | 98.4 | 98.4 |
| Stenotrophomonas phage DLP2 a | AKI28730.1 | 98.3 | - |
| Pseudomonas phage vB_PaeS_SCH_Ab26 | YP_009044360 | 97.9 | 97.4 |
| Pseudomonas phage PaMx42 | YP_009205621 | 69.3 | 69.7 |
| Burkholderia phage KL1 | YP_006560777 | 46.8 | 46.8 |
| Xylella phage Sano a,b | AHB12068 | 29.5 | 29.4 |
| Xylella phage Salvo a,b | AHB12243 | 29 | 28.8 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCutcheon, J.G.; Peters, D.L.; Dennis, J.J. Identification and Characterization of Type IV Pili as the Cellular Receptor of Broad Host Range Stenotrophomonas maltophilia Bacteriophages DLP1 and DLP2. Viruses 2018, 10, 338. https://doi.org/10.3390/v10060338
McCutcheon JG, Peters DL, Dennis JJ. Identification and Characterization of Type IV Pili as the Cellular Receptor of Broad Host Range Stenotrophomonas maltophilia Bacteriophages DLP1 and DLP2. Viruses. 2018; 10(6):338. https://doi.org/10.3390/v10060338
Chicago/Turabian StyleMcCutcheon, Jaclyn G., Danielle L. Peters, and Jonathan J. Dennis. 2018. "Identification and Characterization of Type IV Pili as the Cellular Receptor of Broad Host Range Stenotrophomonas maltophilia Bacteriophages DLP1 and DLP2" Viruses 10, no. 6: 338. https://doi.org/10.3390/v10060338
APA StyleMcCutcheon, J. G., Peters, D. L., & Dennis, J. J. (2018). Identification and Characterization of Type IV Pili as the Cellular Receptor of Broad Host Range Stenotrophomonas maltophilia Bacteriophages DLP1 and DLP2. Viruses, 10(6), 338. https://doi.org/10.3390/v10060338