Protection against Virulent Infectious Bronchitis Virus Challenge Conferred by a Recombinant Baculovirus Co-Expressing S1 and N Proteins

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Virus and the Expression Vector
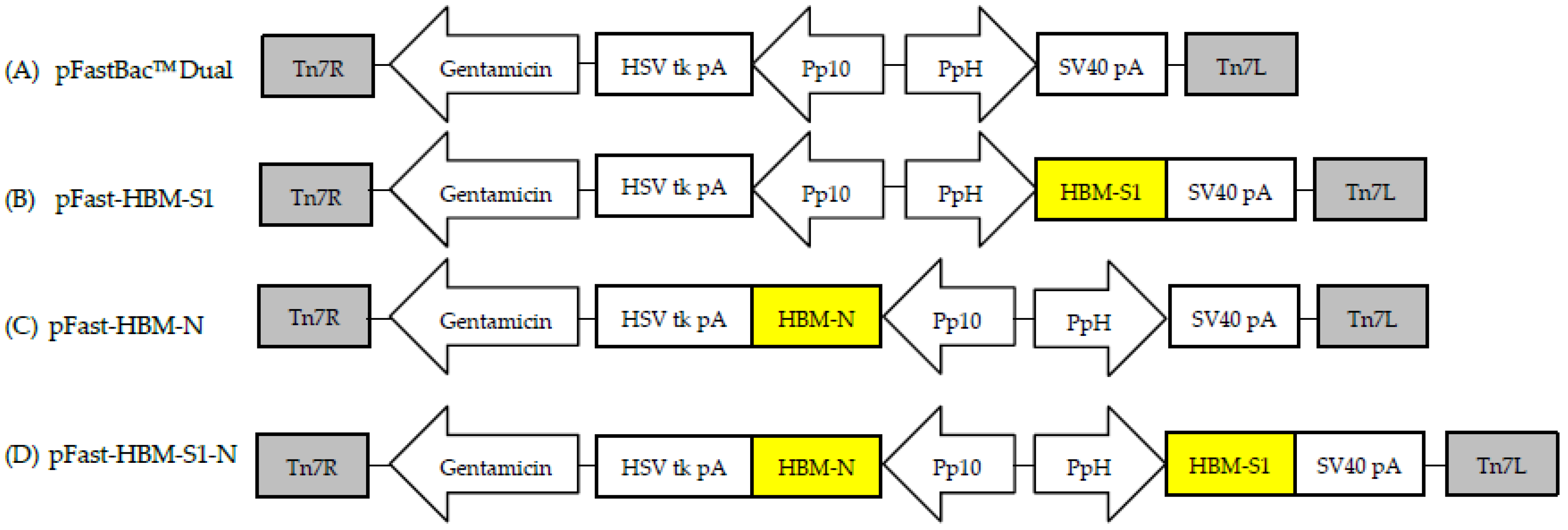
2.2. Gene Cloning and the Recombinant Baculoviruses’ Construction
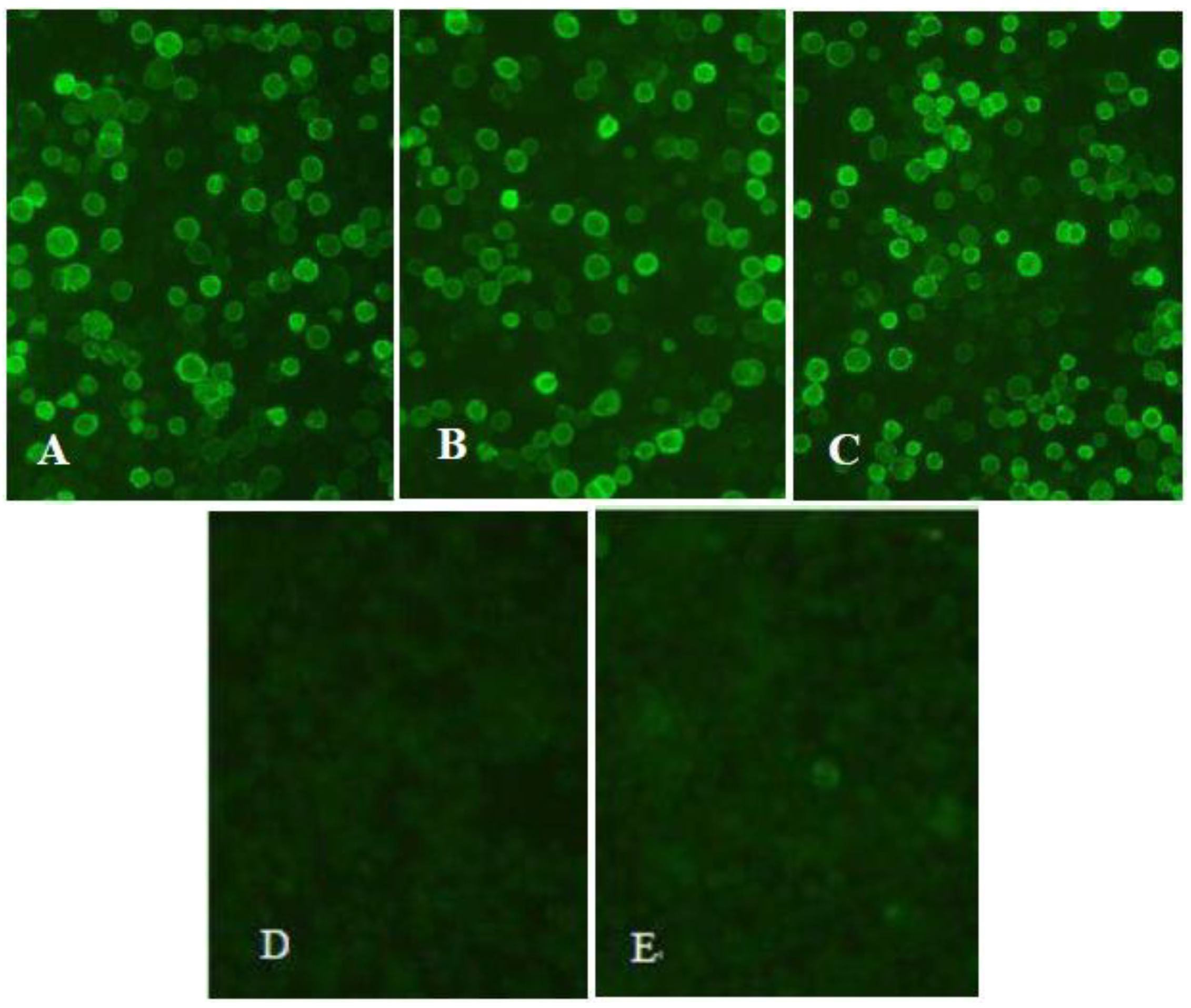
2.3. Analysis of Recombinants’ Protein Expression
2.4. Immunization of the Birds and the Virus Challenge
2.5. The Detection of IBV-Specific Antibody
2.6. Quantification of CD4+ and CD8+ T Lymphocytes
2.7. Virus Re-Isolation
2.8. Statistical Analysis
3. Results
3.1. Confirmation of the Recombinant Baculoviruses
3.2. Identification of the Recombinant Proteins
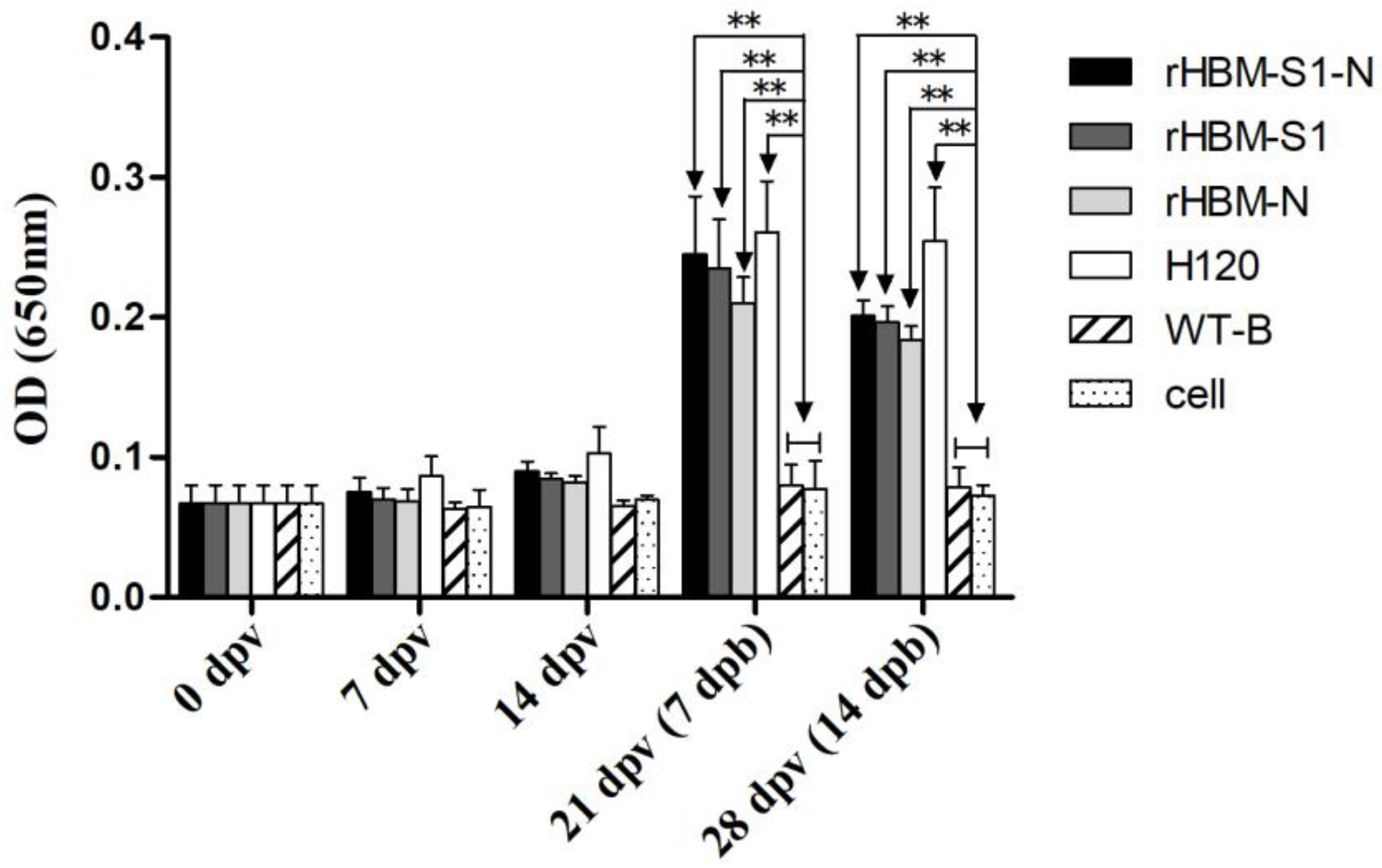
3.3. Antibody Responses Following the Immunizations
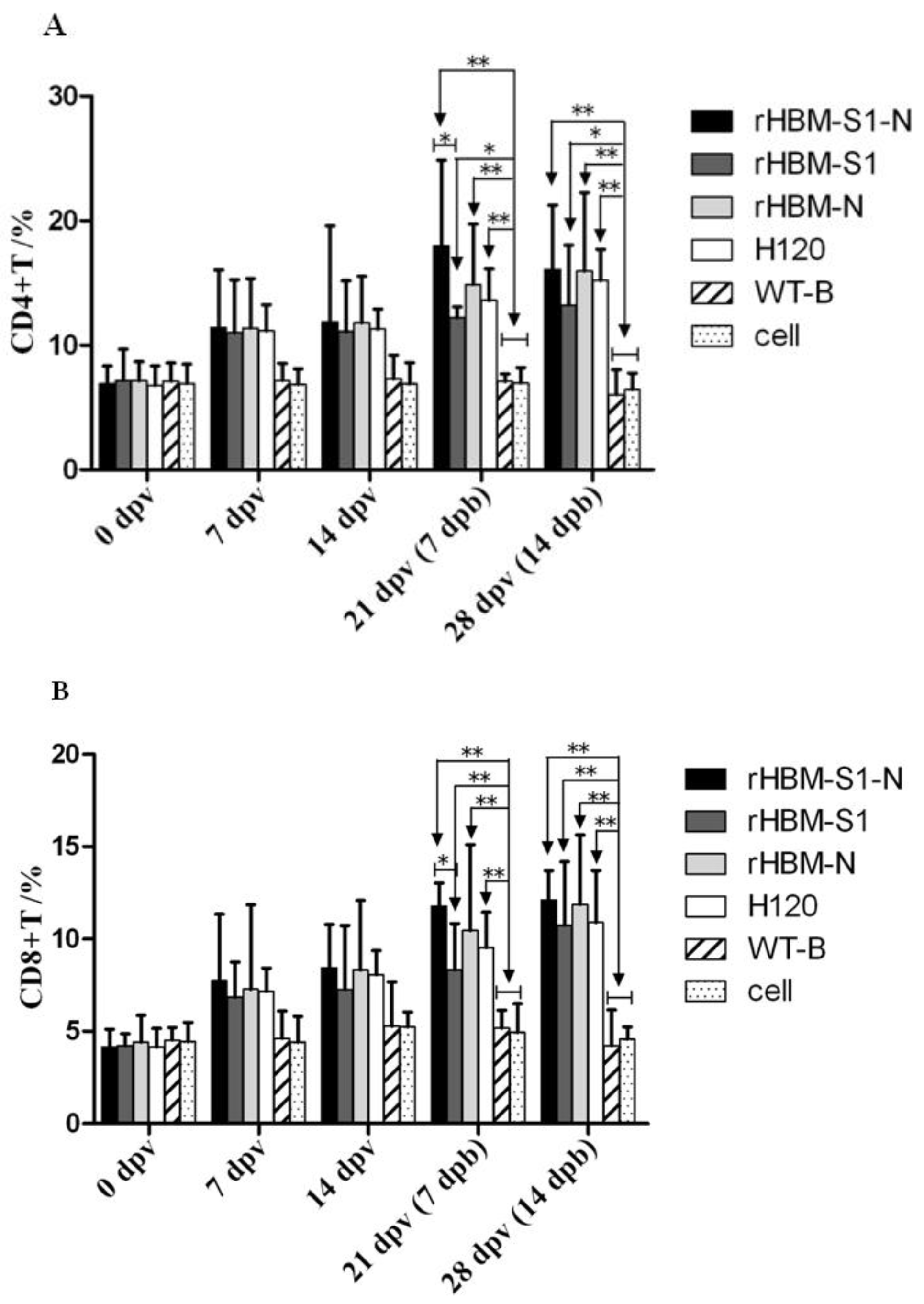
3.4. Dynamic Changes of CD4+ and CD8+ T Lymphocytes in Peripheral Blood
3.5. Protection against the IBV Challenge
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- De Wit, J.J.; Cook, J.K.; van der Heijden, H.M. Infectious bronchitis virus variants: A review of the history, current situation and control measures. Avian Pathol. 2011, 40, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Wit, J.J.; Swart, W.A.; Fabri, T.H. Efficacy of infectious bronchitis virus vaccinations in the field: Association between the alpha-IBV IgM response, protection and vaccine application parameters. Avian Pathol. 2010, 39, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, M.W. Review of infectious bronchitis virus around the world. Avian Dis. 2012, 56, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Singh, S.D.; Barathidasan, R.; Desingu, P.A.; Chakraborty, S.; Tiwari, R.; Kumar, M.A. Emergence of Avian Infectious Bronchitis Virus and its variants need better diagnosis, prevention and control strategies: A global perspective. Pak. J. Biol. Sci. 2014, 17, 751–767. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.F.; Huang, Q.H.; Lu, M.; Wu, J.Q.; Lin, S.Q.; Zhu, F.Z.; Zhang, X.M.; Huang, Y.Y.; Yang, S.H.; Xu, C.T. Attenuation mechanism of virulent infectious bronchitis virus strain with QX genotype by continuous passage in chicken embryos. Vaccine 2016, 34, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.F.; Zhang, T.T.; Han, Z.X.; Liang, S.L.; Xu, Y.; Xu, Q.Q.; Chen, Y.Q.; Zhao, Y.; Shao, Y.H.; Li, H.X.; et al. Molecular and antigenic characteristics of Massachusetts genotype infectious bronchitis coronavirus in China. Vet. Microbiol. 2015, 181, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Severe acute respiratory syndrome vaccine development: Experiences of vaccination against avian infectious bronchitis coronavirus. Avian Pathol. 2003, 32, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.J.; Wang, H.N.; Zhou, S.; Tian, G.B. Enhancement of the immunogenicity of an infectious bronchitis virus DNA vaccine by a bicistronic plasmid encoding nucleocapsid protein and interleukin-2. J. Virol. Methods 2008, 149, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Tarpey, I.; Orbell, S.J.; Britton, P.; Casais, R.; Hodgson, T.; Lin, F.; Hogan, E.; Cavanagh, D. Safety and efficacy of an infectious bronchitis virus used for chicken embryo vaccination. Vaccine 2006, 24, 6830–6838. [Google Scholar] [CrossRef] [PubMed]
- Van, V.S.; Toro, H. Rapid selection in chickens of subpopulations within ArkDPI-derived infectious bronchitis virus vaccines. Avian Pathol. 2008, 37, 293–306. [Google Scholar] [CrossRef] [Green Version]
- McKinley, E.T.; Deborah, A.H.; Jackwood, M.W. Avian coronavirus infectious bronchitis attenuated live vaccines undergo selection of subpopulations and mutations following vaccination. Vaccine 2008, 26, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.M.; Zhao, Y.; Gao, H.B.; Jing, Z.; Wang, M.; Cui, H.Y.; Tong, G.Z.; Wang, Y.F. Evaluation of recombinant fowlpox virus expressing infectious bronchitis virus S1 gene and chicken interferon- gamma gene for immune protection against heterologous strains. Vaccine 2011, 29, 1576–1582. [Google Scholar] [CrossRef] [PubMed]
- Ladman, B.S.; Pope, C.R.; Ziegler, A.F.; Swieczkowski, T.; Callahan, C.J.; Davison, S.; Gelb, J., Jr. Protection of chickens after live and inactivated virus vaccination against challenge with nephropathogenic infectious bronchitis virus PA/Wolgemuth/98. Avian Dis. 2002, 46, 938–944. [Google Scholar] [CrossRef]
- Meeusen, E.N.; Walker, J.; Peters, A.; Pastoret, P.P.; Jungersen, G. Current status of veterinary vaccines. Clin. Microbiol. Rev. 2007, 20, 489–510. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, H.N.; Wang, X.; Tang, J.N.; Lu, D.; Zhang, Y.F.; Guo, Z.C.; Li, Y.L.; Gao, R.; Kang, R.M. The protective immune response against infectious bronchitis virus induced by multi-epitope based peptide vaccines. Biosci. Biotechnol. Biochem. 2009, 73, 1500–1504. [Google Scholar] [CrossRef] [PubMed]
- Eldemery, F.; Joiner, K.S.; Toro, H.; Van, V.S. Protection against infectious bronchitis virus by spike ectodomain subunit vaccine. Vaccine 2017, 35, 5864–5871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, X.W.; Tong, T.Z.; Ye, Y.; Liao, M.; Fan, H.Y. BacMam virus-based surface display of the infectious bronchitis virus (IBV) S1 glycoprotein confers strong protection against virulent IBV challenge in chickens. Vaccine 2014, 32, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Moeini, H.; Omar, A.R. Progress and challenges toward the development of vaccines against avian infectious bronchitis. J. Immunol. Res. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mo, M.L.; Li, M.; Huang, B.C.; Fan, W.S.; Wei, P.; Wei, T.C.; Cheng, Q.Y.; Wei, Z.J.; Lang, Y.H. Molecular Characterization of Major Structural Protein Genes of Avian Coronavirus Infectious Bronchitis Virus Isolates in Southern China. Viruses 2013, 5, 3007–3020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanagh, D.; Davis, P.J.; Cook, J.K.; Li, D.; Kant, A.; Koch, G. Location of the amino acid differences in the S1 spike glycoprotein subunit of closely related serotypes of infectious bronchitis virus. Avian Pathol. 1992, 21, 33–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanagh, D.; Davis, P.J.; Darbyshire, J.H.; Peters, R.W. Coronavirus IBV: Virus retaining spike glycopolypeptide S2 but not S1 is unable to induce virus-neutralizing or haemagglutination-inhibiting antibody, or induce chicken tracheal protection. J. Gen. Virol. 1986, 67, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.M.; Jackwood, M.W.; Hilt, D.A. Identification of amino acids involved in a serotype and neutralization specific epitope within the s1 subunit of avian infectious bronchitis virus. Arch. Virol. 1997, 142, 2249–2256. [Google Scholar] [CrossRef] [PubMed]
- Wickramasinghe, I.N.; van Beurden, S.J.; Weerts, E.A.; Verheije, M.H. The avian coronavirus spike protein. Virus Res. 2014, 194, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.H.; Wang, L.; Smith, R.; Collisson, E.W. The carboxyl-terminal 120-residue polypeptide of infectious bronchitis virus nucleocapsid induces cytotoxic T lymphocytes and protects chickens from acute infection. J. Virol. 1997, 71, 7889–7894. [Google Scholar] [PubMed]
- Yu, D.; Han, Z.X.; Xu, J.; Shao, Y.H.; Li, H.X.; Kong, X.G.; Liu, S.W. A novel B-cell epitope of avian infectious bronchitis virus N protein. Virol. Immunol. 2010, 23, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Lee, J.H.; Hung, C.F.; Peng, S.; Roden, R.; Wang, M.C.; Viscidi, R.; Tsai, Y.C.; He, L.M.; Chen, P.J.; et al. Generation and characterization of DNA vaccines targeting the nucleocapsid protein of severe acute respiratory syndrome coronavirus. J. Virol. 2004, 78, 4638–4645. [Google Scholar] [CrossRef] [PubMed]
- Ikonomou, L.; Schneider, Y.J.; Agathos, S.N. Insect cell culture for industrial production of recombinant proteins. Appl. Microbiol. Biotechnol. 2003, 62, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Bahia, D.; Cheung, R.; Buchs, M.; Geisse, S.; Hunt, I. Optimisation of insect cell growth in deep-well blocks: Development of a high-throughput insect cell expression screen. Protein Expr. Purif. 2005, 39, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.M.; Lv, L.S.; Yin, L.J.; Li, X.M.; Luo, D.G.; Liu, K.; Xue, C.Y.; Cao, Y.C. Assembly and immunogenicity of coronavirus-like particles carrying infectious bronchitis virus M and S proteins. Vaccine 2013, 31, 5524–5530. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.S.; Giesow, K.; Keil, G.M. High-level protein expression following single and dual gene cloning of infectious bronchitis virus N and S genes using baculovirus systems. Viral Immunol. 2014, 27, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Song, C.S.; Lee, Y.J.; Lee, C.W.; Sung, H.W.; Kim, J.H.; Mo, I.P.; Izumiya, Y.; Jang, H.K.; Mikami, T. Induction of protective immunity in chickens vaccinated with infectious bronchitis virus S1 glycoprotein expressed by a recombinant baculovirus. J. Gen. Virol. 1998, 79, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.W.; Wu, X.; Wang, H.N.; Ma, B.C.; Ding, M.D.; Yang, X. Assembly and immunogenicity of baculovirus-derived infectious bronchitis virus–like particles carrying membrane, envelope and the recombinant spike proteins. Biotechnol. Lett. 2016, 38, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, X.Y.; Wei, P.; Chen, Q.Y.; Wei, Z.J.; Mo, M.L. Serotype and genotype diversity of infectious bronchitis viruses isolated during 1985-2008 in Guangxi, China. Arch. Virol. 2012, 157, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; Sapats, S. Identification of previously unknown antigenic epitopes on the S and N proteins of avian infectious bronchitis virus. Arch. Virol. 2005, 150, 1813–1831. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; Galli, U. Immune responses to structural proteins of avian infectious bronchitis virus. Avian Pathol. 1995, 24, 313–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meir, R.; Krispel, S.; Simanov, L.; Eliahu, D.; Maharat, O.; Pitcovski, J. Immune responses to mucosal vaccination by the recombinant S1 and N proteins of infectious bronchitis virus. Viral Immunol. 2012, 25, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.W.; Chen, J.F.; Han, Z.X.; Zhang, Q.X.; Shao, Y.H.; Kong, X.G.; Tong, G.Z. Infectious bronchitis virus: S1 gene characteristics of vaccines used in China and efficacy of vaccination against heterologous strains from China. Avian Pathol. 2006, 35, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.W.; Zhang, X.N.; Wang, Y.; Li, C.R.; Liu, Q.R.; Han, Z.X.; Zhang, Q.X.; Kong, X.G.; Tong, G.Z. Evaluation of the protection conferred by commercial vaccines and attenuated heterologous isolates in China against the CK/CH/LDL/97I strain of infectious bronchitis coronavirus. Vet. J. 2009, 79, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.N.; Wang, Y.; Li, C.R.; Liu, Q.R.; Han, Z.; Shao, Y.H.; Liu, S.W.; Kong, X.G. Evaluation of the protection conferred by several avian infectious bronchitis attenuated vaccines against the field strain CK/CH/LDL/97 I in China. Bing Du Xue Bao 2008, 24, 111–116. [Google Scholar]
- Abe, T.; Takahashi, H.; Hamazaki, H.; Miyano-Kurosaki, N.; Matsuura, Y.; Takaku, H. Baculovirus induces an innate immune response and confers protection from lethal influenzavirus infection in mice. J. Immunol. 2003, 171, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.Y.; Luo, Q.; Chen, X.W.; Ye, Y.; Xin, C.A.; Liao, M. Immunogenicity of a recombinant pseudotype baculovirus expressing the S1 protein of infectious bronchitis virus in specific pathogen free (SPF) chickens. Acta Vet. Zootech. Sin. 2013, 44, 256–261. [Google Scholar]
- Dai, Y.B.; Ding, C.; Liu, M.; Chen, D.S.; Pan, J.Y.; Lu, Y.H.; Chen, P.Y.; Cai, B.X. Immunogenicity evaluation of S1 glycoproteins from infectious bronchitis virus isolates Js/95/03 and SD/97/01 expressed by recombinant baculoviruses. Chin. J. Vet. Sci. 2003, 23, 18–21. [Google Scholar]
- Tan, L.; Zhang, Y.Q.; Liu, F.; Yuan, Y.M.; Zhan, Y.J.; Sun, Y.J.; Qiu, X.S.; Meng, C.C.; Song, C.P.; Ding, C. Infectious bronchitis virus poly-epitope-based vaccine protects chickens from acute infection. Vaccine 2016, 34, 5209–5216. [Google Scholar] [CrossRef] [PubMed]
- Ignjatovic, J.; Galli, L. The S1 glycoprotein but not the N or M proteins of avian infectious bronchitis virus induces protection in vaccinated chickens. Arch. Virol. 1994, 138, 117–134. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Hemmi, H.; Miyamoto, H.; Moriishi, K.; Tamura, S.; Takaku, H.; Akira, S.; Matsuura, Y. Involvement of the Toll-like receptor 9 signaling pathway in the induction of innate immunity by baculovirus. J. Virol. 2005, 79, 2847–2858. [Google Scholar] [CrossRef] [PubMed]
- Vihko, P.; Kurkela, R.; Porvari, K.; Herrala, A.; Lindfors, A.; Lindqvist, Y.; Schneider, G. Rat acid phosphatase: Overexpression of active, secreted enzyme by recombinant baculovirus-infected insect cells, molecular properties andcrystallization. Proc. Natl. Acad. Sci. USA 1993, 90, 799–803. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.M.; Tong, L.L.; Grant, D.; Cihlar, T. Expression and characterization of soluble human parainfluenza virus type 1 hemagglutinin-neuraminidase glycoprotein. J. Virol. Methods 2001, 98, 53–61. [Google Scholar] [CrossRef]
- Van der Geld, Y.M.; Smook, M.; Huitema, M.G.; Harmsen, M.C.; Limburg, P.C.; Kallenberg, C. Expression of recombinant proteinase 3, the autoantigen in Wegener’s granulomatosis, in insect cells. J. Immunol. Methods 2002, 264, 195–205. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Morbidity (%) a | Mortality (%) b | Virus Re-Isolation Rate (%) c | Protection Rate (%) d |
|---|---|---|---|---|
| rHBM-S1-N | 20 (2/10) | 0 (0/10) | 40 (4/10) | 60 (6/10) |
| rHBM-S1 | 30 (3/10) | 0 (0/10) | 50 (5/10) | 50 (5/10) |
| rHBM-N | 40 (4/10) | 20 (2/10) | 60 (6/10) | 40 (4/10) |
| H120 | 10 (1/10) | 0 (0/10) | 20 (2/10) | 80 (8/10) |
| WT-B | 60 (6/10) | 30 (3/10) | 80 (8/10) | 20 (2/10) |
| Cell | 70 (7/10) | 40 (4/10) | 100 (10/10) | 0 (0/10) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Y.; Zhang, Z.-P.; He, Y.-N.; Fan, W.-S.; Dong, Z.-H.; Zhang, L.-H.; Sun, X.-K.; Song, L.-L.; Wei, T.-C.; Mo, M.-L.; et al. Protection against Virulent Infectious Bronchitis Virus Challenge Conferred by a Recombinant Baculovirus Co-Expressing S1 and N Proteins. Viruses 2018, 10, 347. https://doi.org/10.3390/v10070347
Yuan Y, Zhang Z-P, He Y-N, Fan W-S, Dong Z-H, Zhang L-H, Sun X-K, Song L-L, Wei T-C, Mo M-L, et al. Protection against Virulent Infectious Bronchitis Virus Challenge Conferred by a Recombinant Baculovirus Co-Expressing S1 and N Proteins. Viruses. 2018; 10(7):347. https://doi.org/10.3390/v10070347
Chicago/Turabian StyleYuan, Yuan, Zhi-Peng Zhang, Yi-Ning He, Wen-Sheng Fan, Zhi-Hua Dong, Li-Hua Zhang, Xin-Kuan Sun, Li-Li Song, Tian-Chao Wei, Mei-Lan Mo, and et al. 2018. "Protection against Virulent Infectious Bronchitis Virus Challenge Conferred by a Recombinant Baculovirus Co-Expressing S1 and N Proteins" Viruses 10, no. 7: 347. https://doi.org/10.3390/v10070347
APA StyleYuan, Y., Zhang, Z. -P., He, Y. -N., Fan, W. -S., Dong, Z. -H., Zhang, L. -H., Sun, X. -K., Song, L. -L., Wei, T. -C., Mo, M. -L., & Wei, P. (2018). Protection against Virulent Infectious Bronchitis Virus Challenge Conferred by a Recombinant Baculovirus Co-Expressing S1 and N Proteins. Viruses, 10(7), 347. https://doi.org/10.3390/v10070347