Transcript Profiling Identifies Early Response Genes against FMDV Infection in PK-15 Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Cell Treatment and RNA Extraction
2.3. Sample Preparation and Quality Control (QC)
2.4. Library Construction and Sequencing
2.5. Sequencing Data Analysis
2.6. Functional Annotation and Differential Expression Analysis of Unigenes
2.7. RT-QPCR Validation of DEGs
3. Results
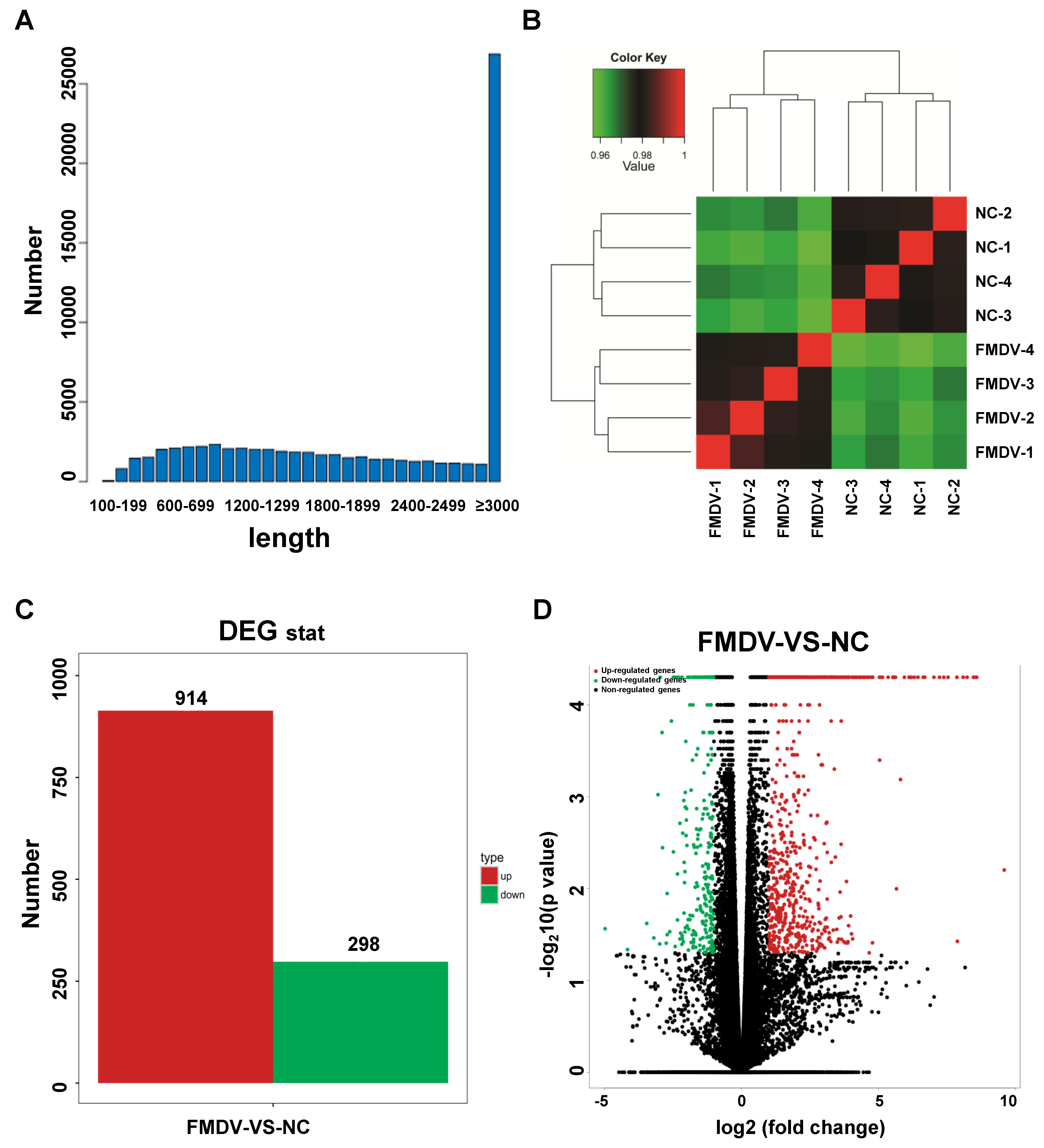
3.1. Quality Control of Sequencing Data
3.2. Assembling Transcripts
3.3. Detection of Sample Repeatability
3.4. Analysis of Differentially Expressed Genes (DEGs)
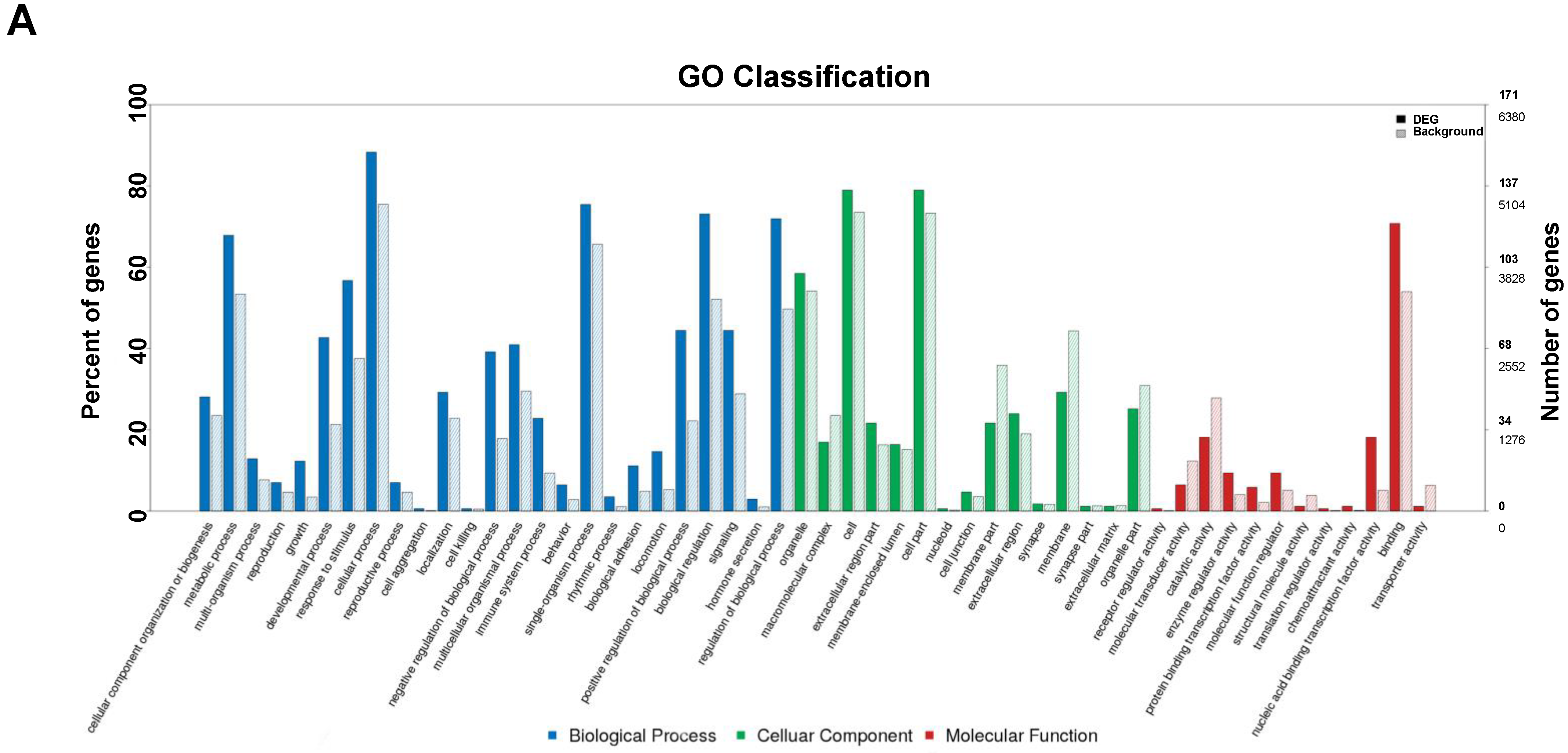
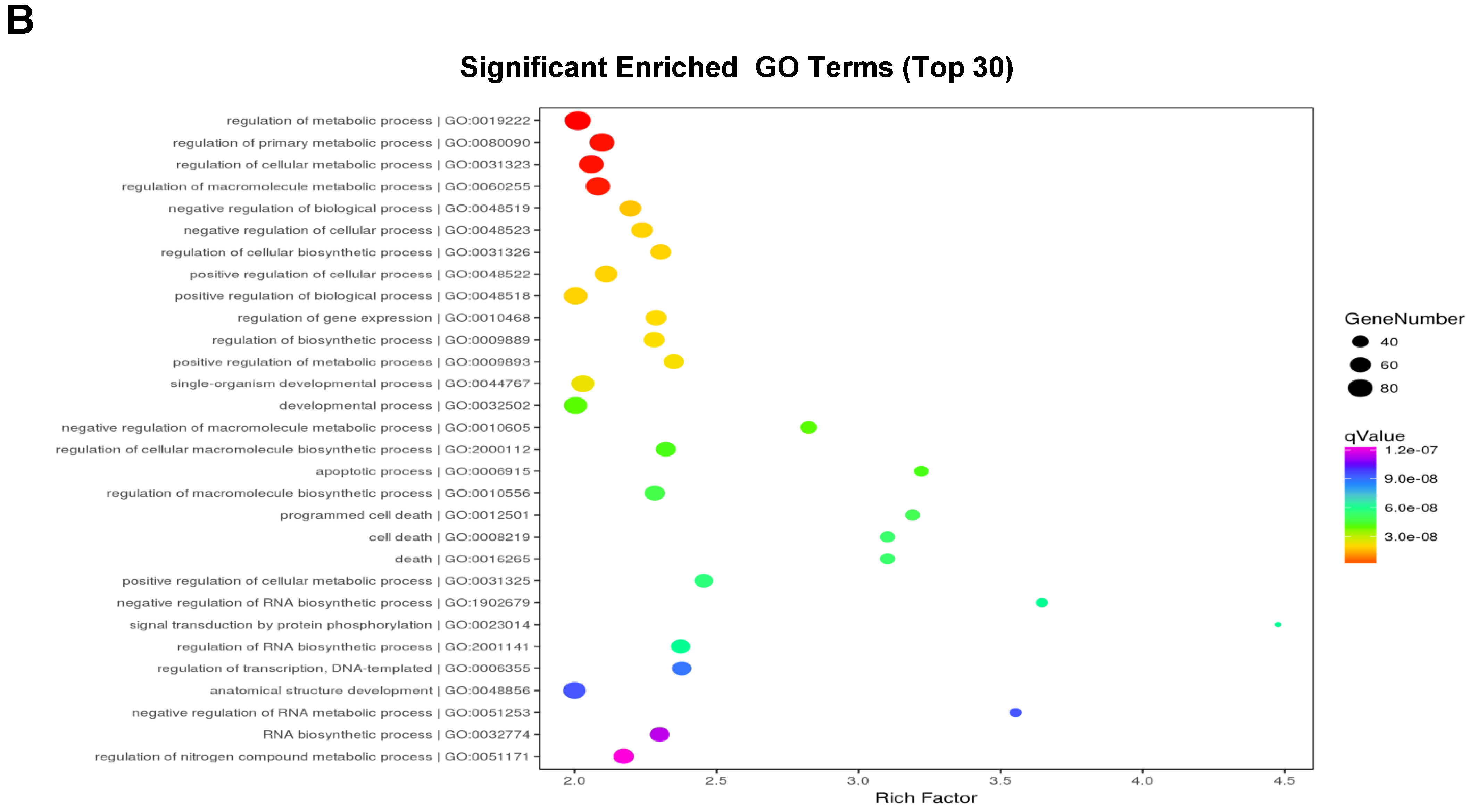
3.5. GO Annotation of DEGs
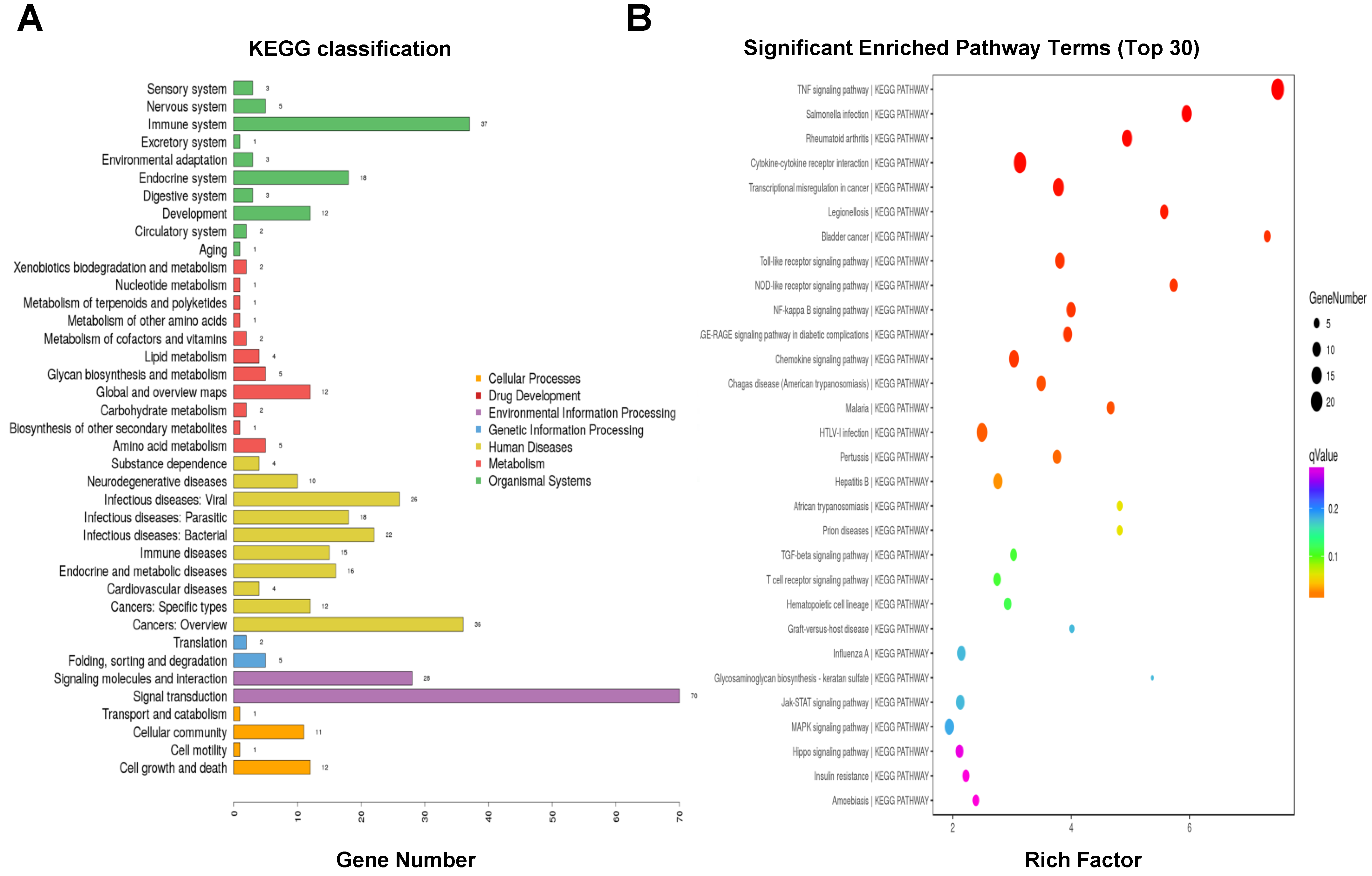
3.6. KEGG Pathway Enrichment Analysis of DEGs
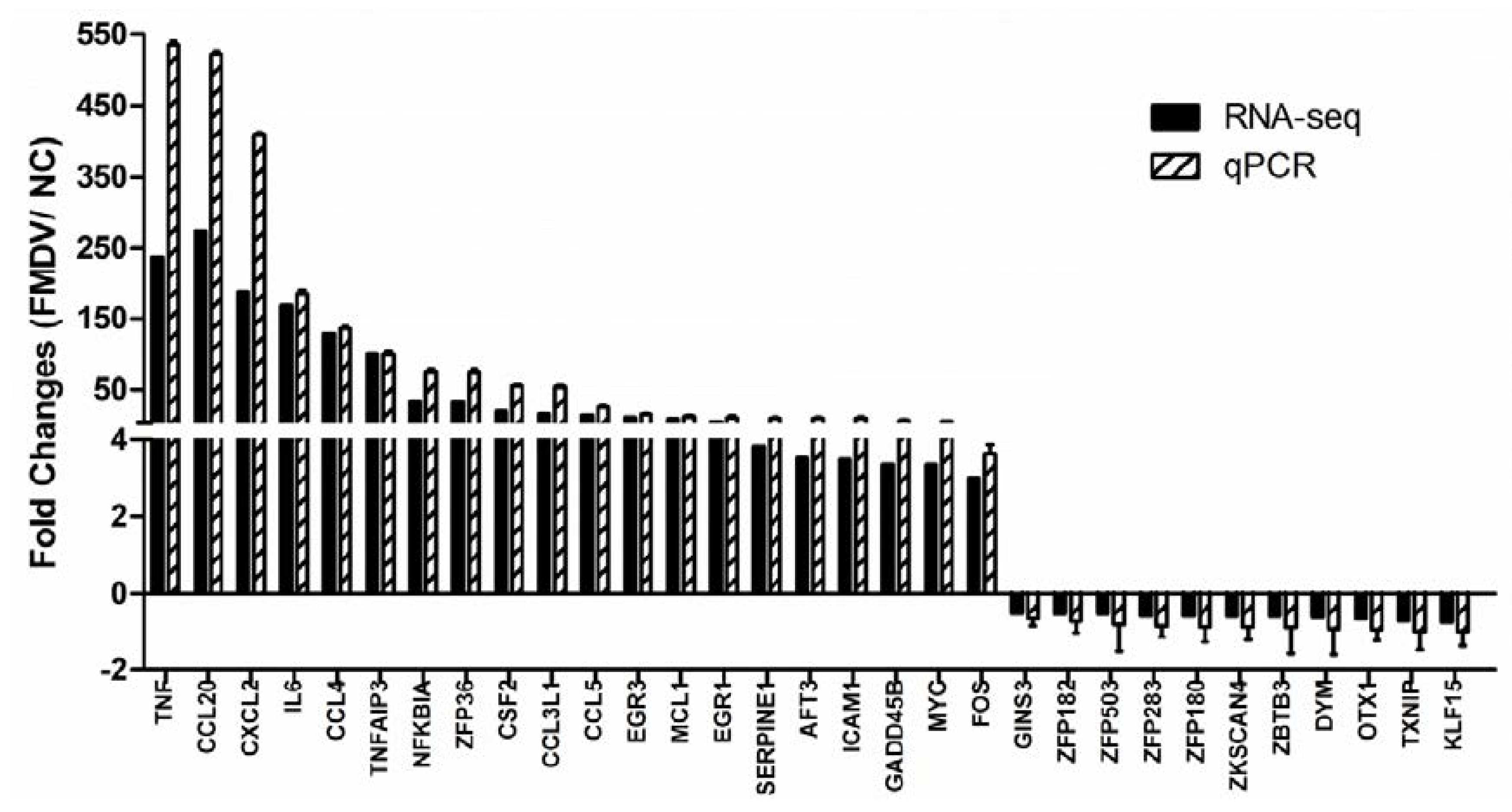
3.7. RT-QPCR Validation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kardjadj, M. Foot-and-mouth disease (FMD) in the Maghreb and its threat to southern European countries. Trop. Anim. Health Prod. 2017, 49, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Shimmon, G.; Wood, B.A.; Morris, A.; Mioulet, V.; Grazioli, S.; Brocchi, E.; Berryman, S.; Tuthill, T.; King, D.P.; Burman, A.; et al. Truncated Bovine Integrin Alpha-v/Beta-6 as a Universal Capture Ligand for FMD Diagnosis. PLoS ONE 2016, 11, e0160696. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, J.M.; Brum, M.C.; Moraes, M.P.; Golde, W.T.; Grubman, M.J. Rapid protection of cattle from direct challenge with foot-and-mouth disease virus (FMDV) by a single inoculation with an adenovirus-vectored FMDV subunit vaccine. Virology 2005, 337, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.R.; Xie, Y.L.; Liu, Z.Z.; Shao, J.J.; Li, S.F.; Zhang, Y.G.; Chang, H.Y. Lithium chloride inhibits early stages of foot-and-mouth disease virus (FMDV) replication in vitro. J. Med. Virol. 2017, 89, 2041–2046. [Google Scholar] [CrossRef] [PubMed]
- Ryoo, S.; Kim, T.; Nah, J.J.; Sagong, M.G.; Lee, S.; Lee, K.N.; Ko, Y.J.; Park, J.H.; Lee, M.H.; Wee, S.H.; et al. Complete Genome Sequence of a Foot-and-Mouth Disease Virus of Serotype O Isolated from Gimje, Republic of Korea, in 2016. Genome Announc. 2017, 5, e01694-16. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.C.; Jin, Y.; Han, S.C.; Sun, S.Q.; Wei, Y.Q.; Liu, X.J.; Feng, X.; Liu, D.X.; Liu, X.T. Quantitative Proteomic Analysis of BHK-21 Cells Infected with Foot-and-Mouth Disease Virus Serotype Asia 1. PLoS ONE 2015, 10, e0132384. [Google Scholar] [CrossRef] [PubMed]
- Cole, S.L.; Ho, L.P. Contribution of innate immune cells to pathogenesis of severe influenza virus infection. Clin. Sci. 2017, 131, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Control of adaptive immunity by the innate immune system. Nat. Immunol. 2015, 16, 343–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husser, L.; Alves, M.P.; Ruggli, N.; Summerfield, A. Identification of the role of RIG-I, MDA-5 and TLR3 in sensing RNA viruses in porcine epithelial cells using lentivirus-driven RNA interference. Virus Res. 2011, 159, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Granberg, F.; Svensson, C.; Pettersson, U.; Zhao, H. Modulation of host cell gene expression during onset of the late phase of an adenovirus infection is focused on growth inhibition and cell architecture. Virology 2005, 343, 236–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Wang, G.; Yang, F.; Cao, W.; Mao, R.; Du, X.; Zhang, X.; Li, C.; Li, D.; Zhang, K.; et al. Foot-and-Mouth Disease Virus Viroporin 2B Antagonizes RIG-I-Mediated Antiviral Effects by Inhibition of Its Protein Expression. J. Virol. 2016, 90, 11106–11121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackie, J.M. A Dictionary of Biomedicine, 1st ed.; Oxford University Press: Oxford, UK, 2010; p. 608. [Google Scholar]
- Du, P.; Sun, S.; Dong, J.; Zhi, X.; Chang, Y.; Teng, Z.; Guo, H.; Liu, Z. Purification of foot-and-mouth disease virus by heparin as ligand for certain strains. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1049–1050, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Kotecha, A.; Wang, Q.; Dong, X.; Ilca, S.L.; Ondiviela, M.; Zihe, R.; Seago, J.; Charleston, B.; Fry, E.E.; Abrescia, N.G.A.; et al. Rules of engagement between alphavbeta6 integrin and foot-and-mouth disease virus. Nat. Commun. 2017, 8, 15408. [Google Scholar] [CrossRef] [PubMed]
- Sompayrac, L. How the Immune System Works, 5th ed.; John Wiley & Sons, Ltd.: Chichester, UK; Ames, IA, USA, 2016. [Google Scholar]
- Brubaker, S.W.; Bonham, K.S.; Zanoni, I.; Kagan, J.C. Innate immune pattern recognition: A cell biological perspective. Annu. Rev. Immunol. 2015, 33, 257–290. [Google Scholar] [CrossRef] [PubMed]
- Haddad, J.J. Cytokines and related receptor-mediated signaling pathways. Biochem. Biophys. Res. Commun. 2002, 297, 700–713. [Google Scholar] [CrossRef]
- Cubillos, C.; de la Torre, B.G.; Jakab, A.; Clementi, G.; Borras, E.; Barcena, J.; Andreu, D.; Sobrino, F.; Blanco, E. Enhanced mucosal immunoglobulin A response and solid protection against foot-and-mouth disease virus challenge induced by a novel dendrimeric peptide. J. Virol. 2008, 82, 7223–7230. [Google Scholar] [CrossRef] [PubMed]
- Francis, M.J.; Black, L. Antibody response in pig nasal fluid and serum following foot-and-mouth disease infection or vaccination. J. Hyg. 1983, 91, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Salt, J.S. The carrier state in foot and mouth disease—An immunological review. Br. Vet. J. 1993, 149, 207–223. [Google Scholar] [CrossRef]
- Sobrino, F.; Saiz, M.; Jimenez-Clavero, M.A.; Nunez, J.I.; Rosas, M.F.; Baranowski, E.; Ley, V. Foot-and-mouth disease virus: A long known virus, but a current threat. Vet. Res. 2001, 32, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Grubman, M.J.; Moraes, M.P.; Diaz-San Segundo, F.; Pena, L.; de los Santos, T. Evading the host immune response: How foot-and-mouth disease virus has become an effective pathogen. FEMS Immunol. Med. Microbiol. 2008, 53, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Summerfield, A.; Guzylack-Piriou, L.; Harwood, L.; McCullough, K.C. Innate immune responses against foot-and-mouth disease virus: Current understanding and future directions. Vet. Immunol. Immunopathol. 2009, 128, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Grubman, M.J.; Baxt, B. Foot-and-mouth disease. Clin. Microbiol. Rev. 2004, 17, 465–493. [Google Scholar] [CrossRef] [PubMed]
- Rigden, R.C.; Carrasco, C.P.; Barnett, P.V.; Summerfield, A.; McCullough, K.C. Innate immune responses following emergency vaccination against foot-and-mouth disease virus in pigs. Vaccine 2003, 21, 1466–1477. [Google Scholar] [CrossRef]
- Barnett, P.V.; Cox, S.J.; Aggarwal, N.; Gerber, H.; McCullough, K.C. Further studies on the early protective responses of pigs following immunisation with high potency foot and mouth disease vaccine. Vaccine 2002, 20, 3197–3208. [Google Scholar] [CrossRef]
- Carbone, D.; Faggio, C. Importance of prebiotics in aquaculture as immunostimulants. Effects on immune system of Sparus aurata and Dicentrarchus labrax. Fish Shellfish Immunol. 2016, 54, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Afonina, I.S.; Zhong, Z.; Karin, M.; Beyaert, R. Limiting inflammation-the negative regulation of NF-kappaB and the NLRP3 inflammasome. Nat. Immunol. 2017, 18, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Jen, J.; Wang, Y.C. Zinc finger proteins in cancer progression. J. Biomed. Sci. 2016, 23, 53. [Google Scholar] [CrossRef] [PubMed]
- Sanduja, S.; Blanco, F.F.; Young, L.E.; Kaza, V.; Dixon, D.A. The role of tristetraprolin in cancer and inflammation. Front. Biosci. 2012, 17, 174–188. [Google Scholar] [CrossRef]
- Van IJzendoorn, D.G.P.; Forghany, Z.; Liebelt, F.; Vertegaal, A.C.; Jochemsen, A.G.; Bovee, J.; Szuhai, K.; Baker, D.A. Functional analyses of a human vascular tumor FOS variant identify a novel degradation mechanism and a link to tumorigenesis. J. Biol. Chem. 2017, 292, 21282–21290. [Google Scholar] [CrossRef] [PubMed]
- Harris, I.S.; Treloar, A.E.; Inoue, S.; Sasaki, M.; Gorrini, C.; Lee, K.C.; Yung, K.Y.; Brenner, D.; Knobbe-Thomsen, C.B.; Cox, M.A.; et al. Glutathione and thioredoxin antioxidant pathways synergize to drive cancer initiation and progression. Cancer Cell 2015, 27, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Krammer, P.; Gülow, K.; Ziola, S. Inhibitors of Thioredoxin-Interacting Protein (TXNIP) for Therapy. U.S. Patent Application 15/218,955[P], 5 January 2017. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Number of Raw Reads | Raw Bases | Number of Clean Reads | Clean Bases | Clean Rate (%) | Q20 (%) | Q30 (%) |
|---|---|---|---|---|---|---|---|
| NC1 | 69,947,474 | 10,471,100,044 | 66,625,990 | 9,975,061,999 | 95.25 | 97.36 | 93.02 |
| NC2 | 73,866,982 | 11,049,947,666 | 70,335,908 | 10,522,323,451 | 95.22 | 97.4 | 93.11 |
| NC3 | 81,300,244 | 12,158,932,746 | 77,781,474 | 11,633,597,601 | 95.67 | 97.58 | 93.49 |
| NC4 | 81,147,108 | 12,142,577,269 | 77,706,416 | 11,629,317,726 | 95.76 | 97.58 | 93.51 |
| FMDV1 | 78,462,120 | 11,740,662,522 | 74,801,356 | 11,193,766,188 | 95.33 | 97.53 | 93.4 |
| FMDV2 | 72,232,906 | 10,805,776,246 | 68,752,730 | 10,285,816,296 | 95.18 | 97.44 | 93.22 |
| FMDV3 | 106,781,936 | 15,980,867,239 | 98,827,180 | 14,792,807,205 | 92.55 | 97.11 | 92.11 |
| FMDV4 | 108,452,172 | 16,218,870,628 | 100,499,600 | 15,030,783,309 | 92.67 | 97.15 | 92.21 |
| Samples | Total Reads | Mapped Reads | Uniquely Mapped Reads | Multiple Mapped Reads |
|---|---|---|---|---|
| NC1 | 66,625,990 (100.00%) | 52,009,891 (78.06%) | 46,518,117 (89.44%) | 5,491,774 (10.56%) |
| NC2 | 70,335,908 (100.00%) | 54,883,603 (78.03%) | 49,189,786 (89.63%) | 5,693,817 (10.37%) |
| NC3 | 77,781,474 (100.00%) | 60,847,412 (78.23%) | 54,541,968 (89.64%) | 6,305,444 (10.36%) |
| NC4 | 77,706,416 (100.00%) | 60,412,369 (77.74%) | 54,154,641 (89.64%) | 6,257,728 (10.36%) |
| FMDV1 | 74,801,356 (100.00%) | 57,611,053 (77.02%) | 51,467,860 (89.34%) | 6,143,193 (10.66%) |
| FMDV2 | 68,752,730 (100.00%) | 52,393,703 (76.21%) | 46,872,931 (89.46%) | 5,520,772 (10.54%) |
| FMDV3 | 98,827,180 (100.00%) | 75,927,134 (76.83%) | 67,639,000 (89.08%) | 8,288,134 (10.92%) |
| FMDV4 | 100,499,600 (100.00%) | 77,810,999 (77.42%) | 69,128,894 (88.84%) | 8,682,105 (11.16%) |
| Sample | Gene Number | Transcript Number | Exon Total Length (bp) | Average Transcript Length (bp) | Max. Transcript Length (bp) | Min. Transcript Length (bp) | N50 Length (bp, without Intron) |
|---|---|---|---|---|---|---|---|
| NC1 | 25,335 | 33,078 | 67,074,593 | 2028 | 24,882 | 156 | 2849 |
| NC2 | 25,818 | 33,561 | 68,649,462 | 2046 | 25,217 | 155 | 2875 |
| NC3 | 26,538 | 34,605 | 71,034,457 | 2053 | 25,181 | 140 | 2900 |
| NC4 | 27,077 | 35,203 | 71,479,661 | 2030 | 24,449 | 149 | 2859 |
| FMDV1 | 25,299 | 33,123 | 66,066,603 | 1995 | 25,261 | 122 | 2814 |
| FMDV2 | 25,499 | 33,311 | 65,027,535 | 1952 | 19,333 | 148 | 2769 |
| FMDV3 | 28,927 | 37,145 | 73,142,162 | 1969 | 22,369 | 141 | 2872 |
| FMDV4 | 29,472 | 37,785 | 73,269,692 | 1939 | 25,126 | 143 | 2839 |
| cuffmerge | 28,141 | 72,673 | 213,510,612 | 2938 | 26,354 | 129 | 4259 |
| GO ID | Category | Gene Number | Representative Genes |
|---|---|---|---|
| GO:0002376 | immune system process | 39 | HOXB6, EPHA2, IL6, CCL5, SOCS1, IRF1, TNFRSF4, CD83, EGR1, IL1A, ZFP36L1, BATF2, CHST3, CSF2, TNF, ADM, CEBPE, CXCL2, SOX9, LIF, ULBP1, EGR3, PHLPP1, CCR7, MAP3K14, CDC37, TNFSF15, CD70, ITPKC, NFKBIA, NR4A3, CCL4, NFIL3 |
| GO:0006955 | immune response | 26 | CSF2, ADM, TNF, CXCL2, IL6, CCL5, LIF, ULBP1, CCR7, MAP3K14, SOCS1, TNFRSF4, IRF1, TNFSF15, CDC37, CD70, NFKBIA, CCL4, IL1A, NR4A3, NFIL3 |
| GO:0002682 | regulation of immune system process | 19 | TNF, SOX9, CCL5, LIF, EGR3, CCR7, PHLPP1, SOCS1, IRF1, CDC37, CD83, ITPKC, NFKBIA, NR4A3, CHST3 |
| GO:0002520 | immune system development | 16 | EPHA2, TNF, CEBPE, IL6, LIF, EGR3, CCR7, PHLPP1, IRF1, CD83, EGR1, NFKBIA, ZFP36L1, BATF2 |
| GO:0002684 | positive regulation of immune system process | 12 | IRF1, CD83, TNF, NFKBIA, CCL5, NR4A3, LIF, EGR3, CCR |
| GO:0045088 | regulation of innate immune response | 4 | NFKBIA, SOCS1, IRF1, CDC37 |
| GO:0050776 | regulation of immune response | 7 | NFKBIA, SOCS1, NR4A3, IRF1, CDC37, CCR7, TNF |
| GO:0002683 | negative regulation of immune system process | 5 | SOX9, NFKBIA, IRF1, TNF |
| GO:0002253 | activation of immune response | 4 | NFKBIA, NR4A3, IRF1, CCR7 |
| GO:0002252 | immune effector process | 8 | IRF1, TNF, ITPKC, NR4A3, IL6, ULBP1, CHST3 |
| GO:0050778 | positive regulation of immune response | 5 | NFKBIA, NR4A3, IRF1, CCR7, TNF |
| GO:0045087 | innate immune response | 7 | NFKBIA, SOCS1, CCL5, CCL4, IRF1, ULBP1, CDC37 |
| GO:0002218 | activation of innate immune response | 2 | NFKBIA, IRF1 |
| GO:0002697 | regulation of immune effector process | 4 | ITPKC, NR4A3, CHST3, TNF |
| GO:0002366 | leukocyte activation involved in immune response | 3 | IL6, NR4A3 |
| GO:0045089 | positive regulation of innate immune response | 2 | NFKBIA, IRF1 |
| GO:0002521 | leukocyte differentiation | 13 | EPHA2, IRF1, CD83, TNF, CEBPE, EGR1, IL6, LIF, ZFP36L1, BATF2, EGR3, CCR7 |
| GO:0097529 | myeloid leukocyte migration | 6 | CCL5, CCL4, CCR7 |
| GO:0007159 | leukocyte cell-cell adhesion | 11 | IRF1, CD83, TNF, EGR1, NR4A3, CCL5, IL6, ZFP36L1, EGR3, CCR7 |
| GO:0045321 | leukocyte activation | 13 | IRF1, CD83, EGR1, IL6, NR4A3, CCL5, ZFP36L1, BATF2, EGR3, CCR7 |
| GO:0050900 | leukocyte migration | 7 | CCL5, CCL4, CCR7, TNF |
| GO:0002274 | myeloid leukocyte activation | 5 | CCL5, NR4A3, BATF2 |
| GO:0070486 | leukocyte aggregation | 10 | IRF1, CD83, EGR1, NR4A3, CCL5, IL6, ZFP36L1, EGR3, CCR7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Chen, H.; Qi, L.; Zhang, J.; Wu, R.; Zhang, Y.; Sun, Y. Transcript Profiling Identifies Early Response Genes against FMDV Infection in PK-15 Cells. Viruses 2018, 10, 364. https://doi.org/10.3390/v10070364
Zhang T, Chen H, Qi L, Zhang J, Wu R, Zhang Y, Sun Y. Transcript Profiling Identifies Early Response Genes against FMDV Infection in PK-15 Cells. Viruses. 2018; 10(7):364. https://doi.org/10.3390/v10070364
Chicago/Turabian StyleZhang, Tianliang, Haotai Chen, Linlin Qi, Jie Zhang, Run Wu, Yongguang Zhang, and Yuefeng Sun. 2018. "Transcript Profiling Identifies Early Response Genes against FMDV Infection in PK-15 Cells" Viruses 10, no. 7: 364. https://doi.org/10.3390/v10070364
APA StyleZhang, T., Chen, H., Qi, L., Zhang, J., Wu, R., Zhang, Y., & Sun, Y. (2018). Transcript Profiling Identifies Early Response Genes against FMDV Infection in PK-15 Cells. Viruses, 10(7), 364. https://doi.org/10.3390/v10070364