Establishment of Baculovirus-Expressed VLPs Induced Syncytial Formation Assay for Flavivirus Antiviral Screening


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Viruses
2.2. Antibodies and Reagents
2.3. Construction of Recombinant Baculoviruses and Plasmids
2.4. Cell Transfection
2.5. Production and Purification of ZIKV VLPs
2.6. TEM and Immune-Electron Microscopy (IEM)
2.7. Western Blot Analysis of Protein Expression
2.8. Immunofluorescence Assay
2.9. Cytotoxicity Assay
2.10. Syncytial Formation Inhibition Assay
2.11. Viral Plaque Assay
2.12. Assays for Antiviral Activity
2.13. Quantitative PCR
2.14. Statistical Analysis
3. Results
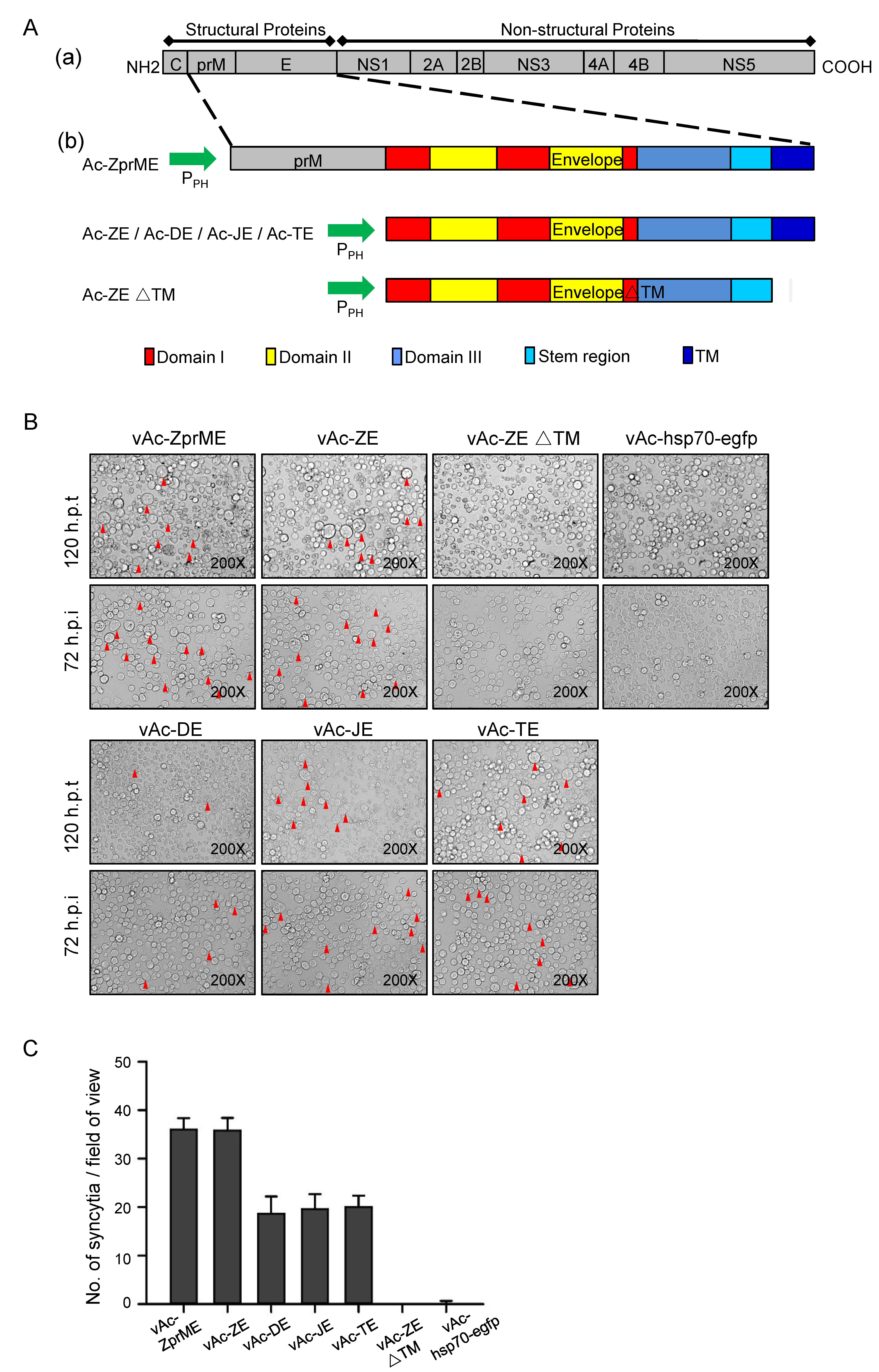
3.1. Generation of Recombinant Baculoviruses Expressing Glycoproteins of Flaviviruses
3.2. Baculovirus-Expressed Flavivirus E Protein Triggers Syncytial Formation
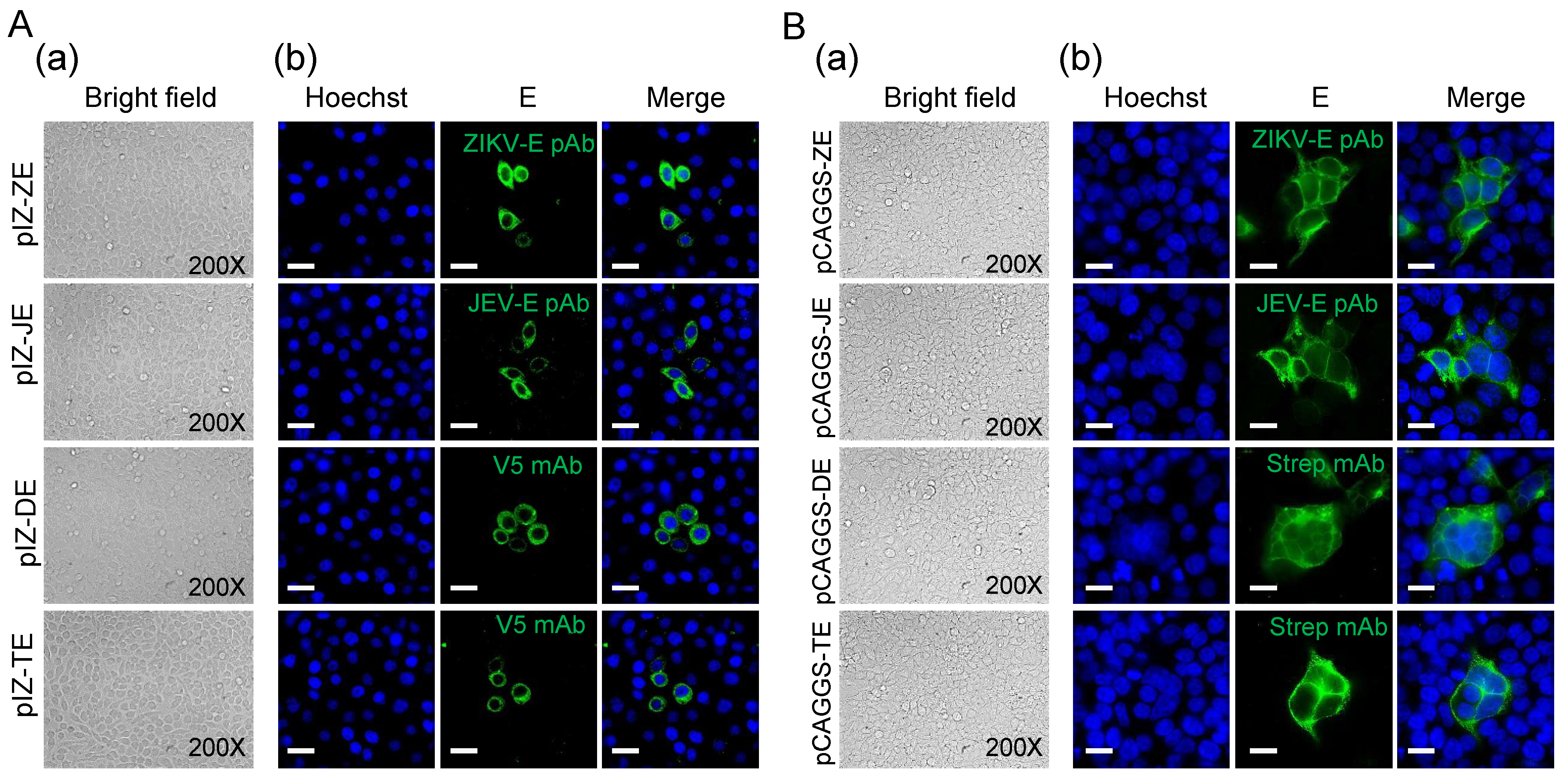
3.3. Localization of Flavivirus E protein in Sf9 Cells
3.4. ZIKV VLPs Were Generated Using Baculovirus Expression System
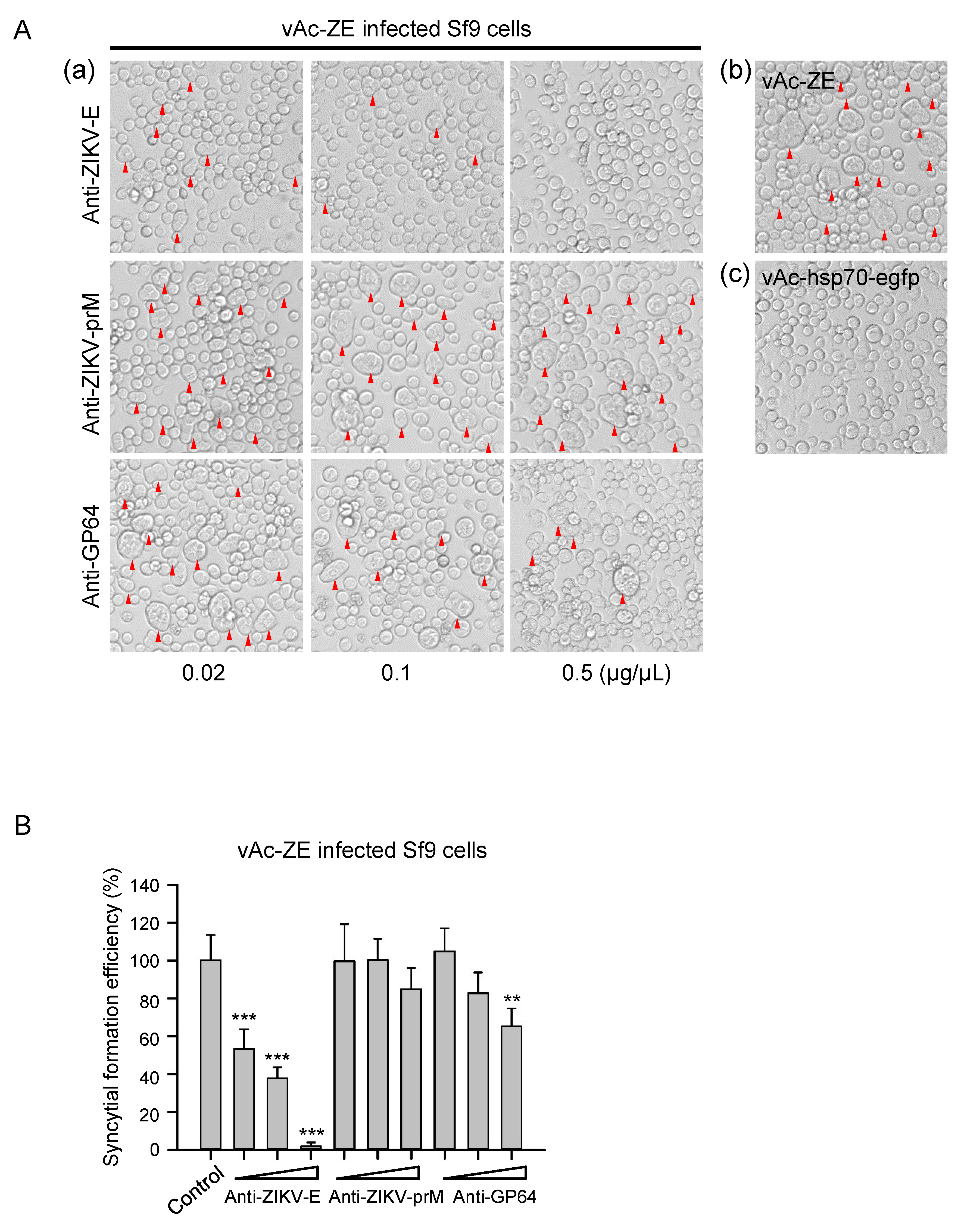
3.5. Syncytial Formation Was Adapted to Screen Antiviral Inhibitor
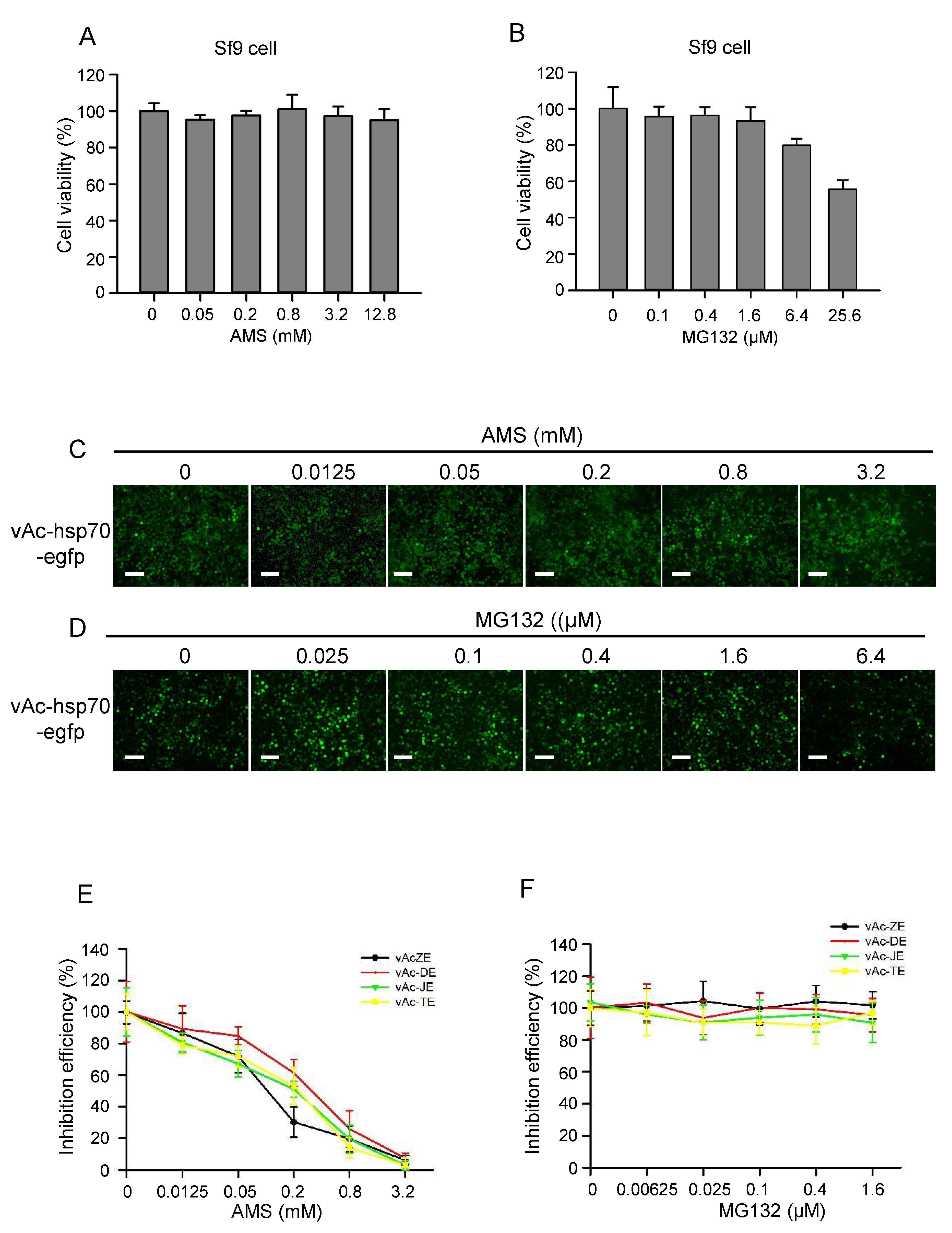
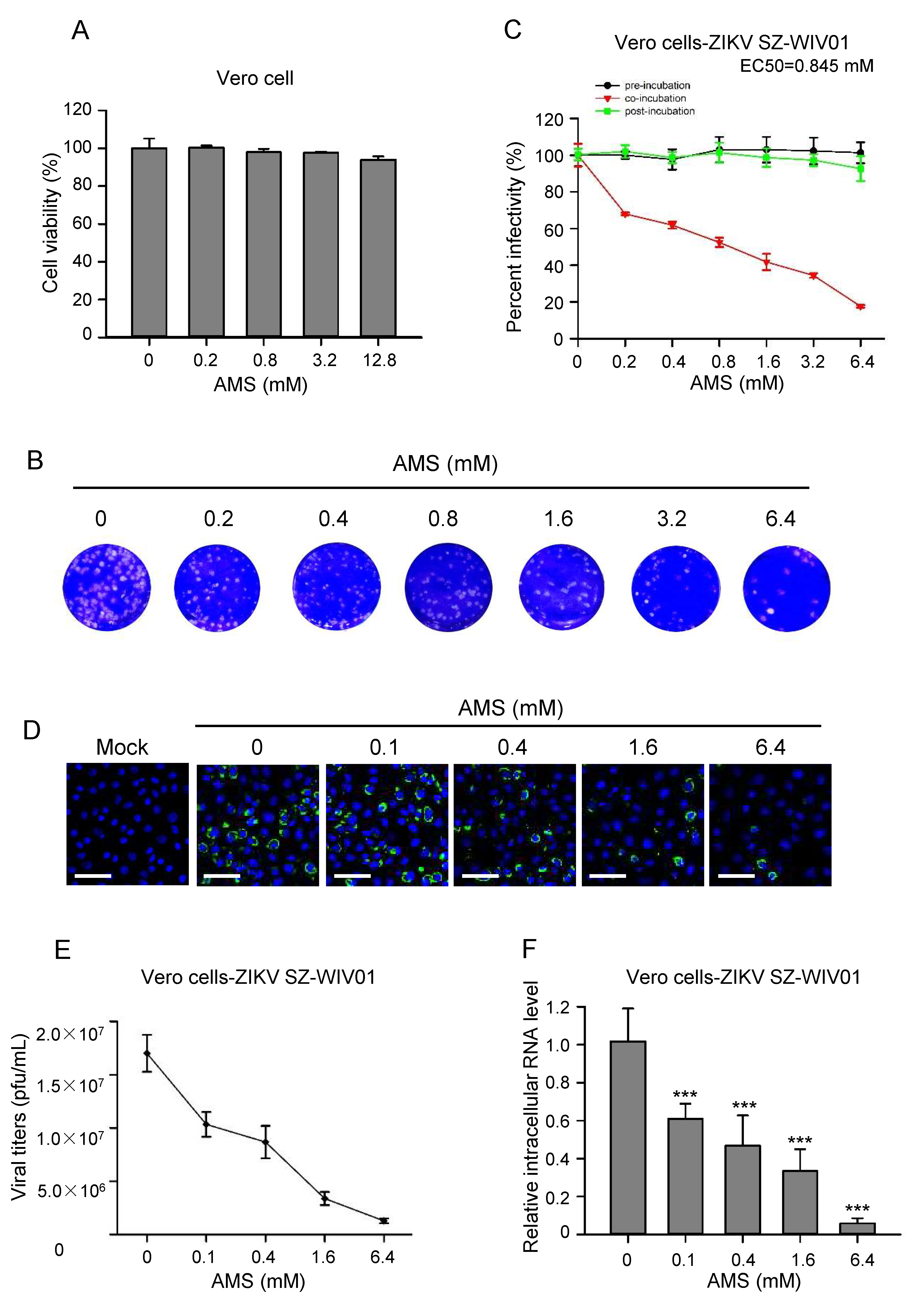
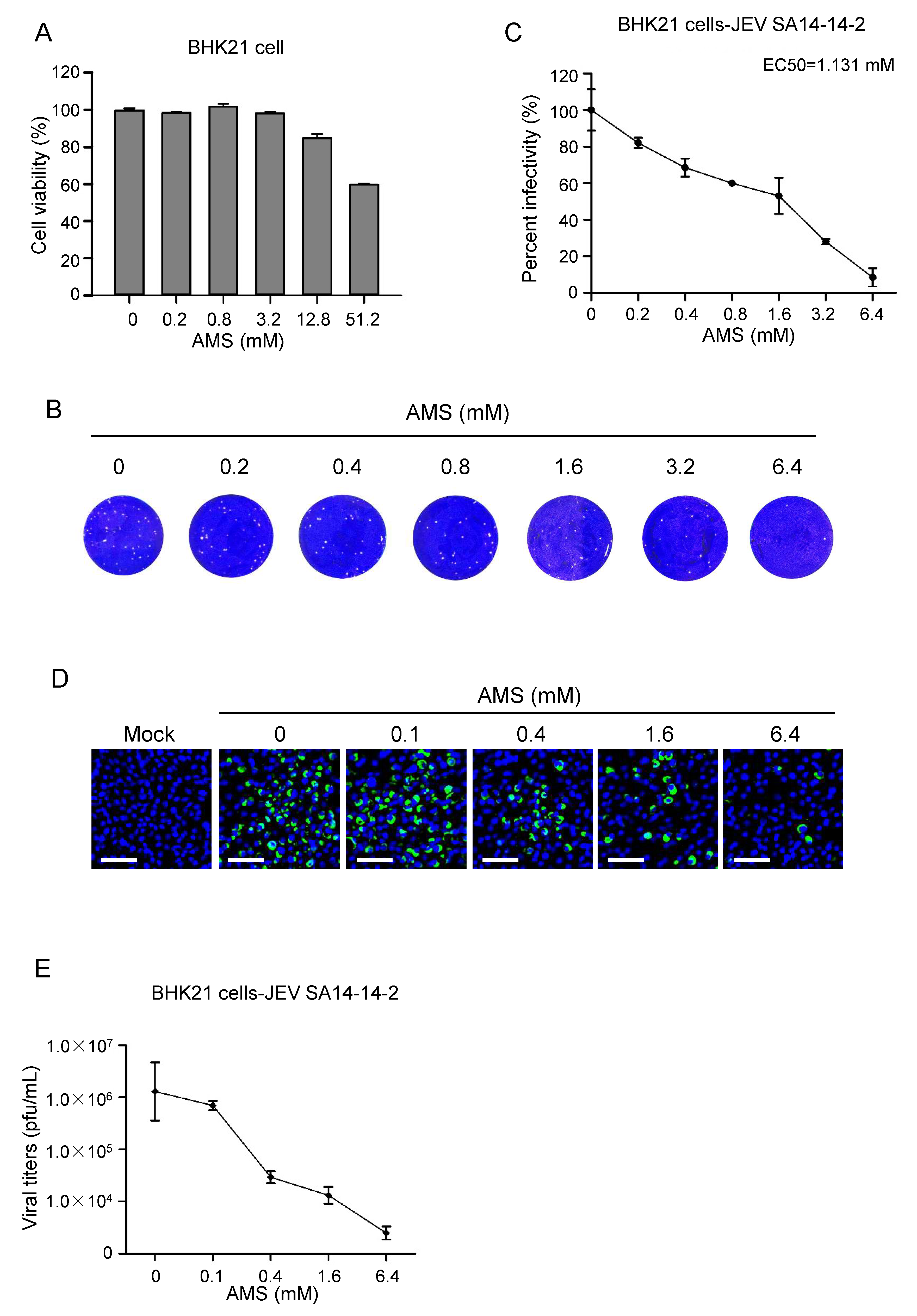
3.6. AMS Broadly Inhibited Flavivirus Infection by Blocking Viral Entry
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Oers, M.M. Opportunities and challenges for the baculovirus expression system. J. Invertebr. Pathol. 2011, 107, S3–S15. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-X.; Wu, X.-D.; Li, L.; Liu, Z.-S.; Wang, Z.-L. Use of baculovirus expression system for generation of virus-like particles: Successes and challenges. Protein Expr. Purif. 2013, 90, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Yamaji, H. Suitability and perspectives on using recombinant insect cells for the production of virus-like particles. Appl. Microbiol. Biotechnol. 2014, 98, 1963–1970. [Google Scholar] [CrossRef] [PubMed]
- Harper, D.-M.; Franco, E.-L.; Wheeler, C.; Ferris, D.-G.; Jenkins, D.; Schuind, A.; Zahaf, T.; Innis, B.; Naud, P.; De Carvalho, N.-S.; et al. Efficacy of a bivalent L1 virus-like particle vaccine in prevention of infection with human papillomavirus types 16 and 18 in young women: A randomised controlled trial. Lancet 2004, 364, 1757–1765. [Google Scholar] [CrossRef]
- Gould, E.; Solomon, T. Pathogenic flaviviruses. Lancet 2008, 371, 500–509. [Google Scholar] [CrossRef]
- Fauci, A.; Morens, D. Zika Virus in the Americas—Yet Another Arbovirus Threat. N. Engl. J. Med. 2016, 374, 601–604. [Google Scholar] [CrossRef] [PubMed]
- Ioos, S.; Mallet, H.-P.; Goffart, I.-L.; Gauthier, V.; Cardoso, T.; Herida, M. Current Zika virus epidemiology and recent epidemics. Med. Mal. Infect. 2014, 44, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Lessler, J.; Chaisson, L.; Kucirka, L.; Bi, Q.; Grantz, K.; Salje, H.; Carcelen, A.; Ott, C.; Sheffield, J.; Ferguson, N.; et al. Assessing the global threat from Zika virus. Science 2016, 353, aaf8160. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.-X.; Stiasny, K. The Antigenic Structure of Zika Virus and Its Relation to Other Flaviviruses: Implications for Infection and Immunoprophylaxis. Microbiol. Mol. Biol. Rev. 2017, 81, e00055-16. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.-X.; Stiasny, K. Flaviviruses and their antigenic structure. J. Clin. Virol. 2012, 55, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Kielian, M.; Rey, F.-A. Virus membrane-fusion proteins: More than one way to make a hairpin. Nat. Rev. Microbiol. 2006, 4, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.-P.; Song, J.; Lu, X.-S.; Deng, Y.-Q.; Musyoki, A.-M.; Cheng, H.-J.; Zhang, Y.-F.; Yuan, Y.; Song, H.; Haywood, J.; et al. Structures of the Zika Virus Envelope Protein and Its Complex with a Flavivirus Broadly Protective Antibody. Cell Host Microbe 2016, 19, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Fu, D.-W.; Zhang, P.-F. Establishment and characterization of Japanese B encephalitis virus persistent infection in the Sf9 insect cell line. Biologicals 1996, 24, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-F.; Klutch, M.; Muller, J.; Marcus-Sekura, C. St Louis encephalitis virus establishes a productive, cytopathic and persistent infection of Sf9 cells. J. Gen. Virol. 1993, 74, 1703–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, C.-L.; Liu, S.-Q.; Zhang, Q.-Y.; Xu, M.-Y.; Zhang, H.-L.; Gu, D.-Y.; Shi, L.; He, J.-A.; Xiao, G.-F.; Zhang, B. Isolation and characterization of Zika virus imported to China using C6/36 mosquito cells. Virol. Sin. 2016, 31, 176–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, R.-K.; Yin, F.-F.; Wang, M.-L.; Hu, Z.-H.; Wang, H.-L.; Deng, F. Glycoprotein E of the Japanese encephalitis virus forms virus-like particles and induces syncytia when expressed by a baculovirus. J. Gen. Virol. 2015, 96, 1006–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.-L.; Tan, Y.; Yin, F.-F.; Deng, F.; Vlak, J.; Hu, Z.-H.; Wang, H.-L. The F-like protein Ac23 enhances the infectivity of the budded virus of gp64-null Autographa californica multinucleocapsid nucleopolyhedrovirus pseudotyped with baculovirus envelope fusion protein F. J. Virol. 2008, 82, 9800–9804. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.-Y.; Zhang, T.; Zhang, Y.-F.; Wang, H.-L.; Deng, F. Zika Virus Baculovirus-Expressed Virus-Like Particles Induce Neutralizing Antibodies in Mice. Virol. Sin. 2018, 33, 213–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.-D.; Livak, K.-J. Analyzing real-time PCR data by the comparative C-T method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-F.; Klutch, M.; Muller, J.; Marcus-Sekura, C. Susceptibility of the Sf9 insect cell line to infection with adventitious viruses. Biologicals 1994, 22, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.-C.; Wang, M.-L.; Qiu, Z.-J.; Deng, F.; Vlak, J.-M.; Hu, Z.-H.; Wang, H.-L. Autographa californica Multicapsid Nucleopolyhedrovirus Efficiently Infects Sf9 Cells and Transduces Mammalian Cells via Direct Fusion with the Plasma Membrane at Low pH. J. Virol. 2010, 84, 5351–5359. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.; Boo, I.; Poumbourios, P.; Drummer, H. Hepatitis C virus (HCV) envelope glycoproteins E1 and E2 contain reduced cysteine residues essential for virus entry. J. Biol. Chem. 2011, 286, 31984–31992. [Google Scholar] [CrossRef] [PubMed]
- Ryser, H.; Levy, E.; Mandel, R.; DiSciullo, G. Inhibition of human immunodeficiency virus infection by agents that interfere with thiol-disulfide interchange upon virus-receptor interaction. Proc. Natl. Acad. Sci. USA 1994, 91, 4559–4563. [Google Scholar] [CrossRef] [PubMed]
- Strandin, T.; Hepojoki, J.; Wang, H.; Vaheri, A.; Lankinen, H. Inactivation of hantaviruses by N-ethylmaleimide preserves virion integrity. J. Gen. Virol. 2011, 92, 1189–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Jaoudé, G.; Sureau, C. Entry of hepatitis delta virus requires the conserved cysteine residues of the hepatitis B virus envelope protein antigenic loop and is blocked by inhibitors of thiol-disulfide exchange. J. Virol. 2007, 81, 13057–13066. [Google Scholar] [CrossRef] [PubMed]
- Xin, Q.-L.; Deng, C.-L.; Chen, X.; Wang, J.; Wang, S.-B.; Wang, W.; Deng, F.; Zhang, B.; Xiao, G.-F.; Zhang, L.-K. Quantitative Proteomic Analysis of Mosquito C6/36 Cells Reveals Host Proteins Involved in Zika Virus Infection. J. Virol. 2017, 91, e00554-17. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-J.; Peng, J.-G.; Wu, S.-C. Characterization of the GXXXG motif in the first transmembrane segment of Japanese encephalitis virus precursor membrane (prM) protein. J. Biomed. Sci. 2010, 17, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barros, M.; Galasso, T.; Chaib, A.; Degallier, N.; Nagata, T.; Ribeiro, B. Yellow fever virus envelope protein expressed in insect cells is capable of syncytium formation in lepidopteran cells and could be used for immunodetection of YFV in human sera. Virol. J. 2011, 8, 261. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Metz, S. Progress and Works in Progress: Update on Flavivirus Vaccine Development. Clin. Ther. 2017, 39, 1519–1536. [Google Scholar] [CrossRef] [PubMed]
- Wahid, B.; Ali, A.; Rafique, S.; Idrees, M. Current status of therapeutic and vaccine approaches against Zika virus. Eur. J. Intern. Med. 2017, 44, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Boldescu, V.; Behnam, M.; Vasilakis, N.; Klein, C. Broad-spectrum agents for flaviviral infections: Dengue, Zika and beyond. Nat. Rev. Drug Discov. 2017, 16, 565–586. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.-F.; Deng, Y.-Q.; Zou, P.; Wang, Q.; Dai, Y.-Y.; Yu, F.; Du, L.-Y.; Zhang, N.-N.; Tian, M.; Hao, J.-N.; et al. A peptide-based viral inactivator inhibits Zika virus infection in pregnant mice and fetuses. Nat. Commun. 2017, 8, 15672. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-L.; Yang, H.-B.; Liu, X.-Q.; Dai, L.-P.; Ma, T.; Qi, J.-X.; Wong, G.; Peng, R.-C.; Liu, S.; Li, J.-F.; et al. Molecular determinants of human neutralizing antibodies isolated from a patient infected with Zika virus. Sci. Transl. Med. 2016, 8, 369ra179. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Murali, A.; Singh, S.; Giri, R. Epigallocatechin gallate, an active green tea compound inhibits the Zika virus entry into host cells via binding the envelope protein. Int. J. Biol. Macromol. 2017, 104, 1046–1054. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) |
|---|---|
| Actin-F | CATCCGTAAAGACCTCTATGCCAAC |
| Actin-R | ATGGAGCCACCGATCCACA |
| ZIKV-E-F | ACTGGTAGAGTTCAAGGACGCAC |
| ZIKV-E-R | TTCAGGCGACATTTCAAGTGG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, S.; Zhang, Y.; Zhang, T.; Zhang, B.; Wang, H.; Deng, F. Establishment of Baculovirus-Expressed VLPs Induced Syncytial Formation Assay for Flavivirus Antiviral Screening. Viruses 2018, 10, 365. https://doi.org/10.3390/v10070365
Dai S, Zhang Y, Zhang T, Zhang B, Wang H, Deng F. Establishment of Baculovirus-Expressed VLPs Induced Syncytial Formation Assay for Flavivirus Antiviral Screening. Viruses. 2018; 10(7):365. https://doi.org/10.3390/v10070365
Chicago/Turabian StyleDai, Shiyu, Yanfang Zhang, Tao Zhang, Bo Zhang, Hualin Wang, and Fei Deng. 2018. "Establishment of Baculovirus-Expressed VLPs Induced Syncytial Formation Assay for Flavivirus Antiviral Screening" Viruses 10, no. 7: 365. https://doi.org/10.3390/v10070365
APA StyleDai, S., Zhang, Y., Zhang, T., Zhang, B., Wang, H., & Deng, F. (2018). Establishment of Baculovirus-Expressed VLPs Induced Syncytial Formation Assay for Flavivirus Antiviral Screening. Viruses, 10(7), 365. https://doi.org/10.3390/v10070365