Transcriptome Analysis and In Situ Hybridization for FcaGHV1 in Feline Lymphoma


 , and
, and 
Abstract
:Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Troyer, R.M.; Beatty, J.A.; Stutzman-Rodriguez, K.R.; Carver, S.; Lozano, C.C.; Lee, J.S.; Lappin, M.R.; Riley, S.P.D.; Serieys, L.E.K.; Logan, K.A.; et al. Novel Gammaherpesviruses in North American Domestic Cats, Bobcats, and Pumas: Identification, Prevalence, and Risk Factors. J. Virol. 2014, 88, 3914–3924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatty, J.A.; Troyer, R.M.; Brewster, C.; Rovnak, R.; Barrs, V.R.; Quackenbush, S.L.; VandeWoude, S. Feline immunodeficiency virus (FIV)-associated lymphoma. Is a gammaherpesvirus involved? In Proceedings of the 2nd International Society for Companion Animal Infectious Diseases Symposium, San Fraancisco, CA, USA, 14–17 November 2012; p. 27. [Google Scholar]
- Pinzone, M.R.; Berretta, M.; Cacopardo, B.; Nunnari, G. Epstein-Barr Virus- and Kaposi Sarcoma-Associated Herpesvirus-Related Malignancies in the Setting of Human Immunodeficiency Virus Infection. Semin. Oncol. 2015, 42, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M. Pathogenesis of gammaherpesvirus infections. Vet. Microbiol. 2006, 113, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Pierangeli, A.; Antonelli, G.; Gentile, G. Immunodeficiency-associated viral oncogenesis. Clin. Microbiol. Infect. 2015, 21, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Tierney, R.J.; Shannon-Lowe, C.D.; Fitzsimmons, L.; Bell, A.I.; Rowe, M. Unexpected patterns of Epstein–Barr virus transcription revealed by a High throughput PCR array for absolute quantification of viral mRNA. Virology 2015, 474, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Grundhoff, A.; Ganem, D. Inefficient establishment of KSHV latency suggests an additional role for continued lytic replication in Kaposi sarcoma pathogenesis. J. Clin. Investig. 2004, 113, 124–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshino, Y.; Katano, H.; Zou, P.; Hohman, P.; Marques, A.; Tyring, S.K.; Follmann, D.; Cohen, J.I. Long-Term Administration of Valacyclovir Reduces the Number of Epstein-Barr Virus (EBV)-Infected B Cells but Not the Number of EBV DNA Copies per B Cell in Healthy Volunteers. J. Virol. 2009, 83, 11857–11861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borozan, I.; Zapatka, M.; Frappier, L.; Ferretti, V. Analysis of Epstein-Barr Virus Genomes and Expression Profiles in Gastric Adenocarcinoma (vol 92, e01239-17, 2018). J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J.A.; Troyer, R.M.; Carver, S.; Barrs, V.R.; Espinasse, F.; Conradi, O.; Stutzman-Rodriguez, K.; Chan, C.C.; Tasker, S.; Lappin, M.R.; et al. Felis catus gammaherpesvirus 1; a widely endemic potential pathogen of domestic cats. Virology 2014, 460, 100–107. [Google Scholar] [CrossRef] [PubMed]
- McLuckie, A.; Tasker, S.; Dhand, N.K.; Spencer, S.; Beatty, J.A. High prevalence of Felis catus gammaherpesvirus 1 infection in haemoplasma-infected cats supports co-transmission. Vet. J. 2016, 214, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Ertl, R.; Korb, M.; Langbein-Detsch, I.; Klein, D. Prevalence and risk factors of gammaherpesvirus infection in domestic cats in Central Europe. Virol. J. 2015, 12, 146. [Google Scholar] [CrossRef] [PubMed]
- Tateno, M.; Endo, Y.; Takahashi, M. An Epidemiological Study of Gammaherpesviruses in Domestic Cats in Japan. J. Vet. Intern. Med. 2016, 30, 1476. [Google Scholar]
- Kurissio, J.K.; Rodrigues, M.V.; Taniwaki, S.A.; Zanutto, M.D.; Filoni, C.; Galdino, M.V.; Araujo, J.P. Felis catus gammaherpesvirus 1 (FcaGHV1) and coinfections with feline viral pathogens in domestic cats in Brazil. Cienc. Rural 2018, 48. [Google Scholar] [CrossRef]
- Stutzman-Rodriguez, K.; Rovnak, J.; VandeWoude, S.; Troyer, R.M. Domestic cats seropositive for Felis catus gammaherpesvirus 1 are often qPCR negative. Virology 2016, 498, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liem, B.; Dhand, N.; Pepper, A.; Barrs, V.; Beatty, J. Clinical Findings and Survival in Cats Naturally Infected with Feline Immunodeficiency Virus. J. Vet. Intern. Med. 2013, 27, 798–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLuckie, A.; Barrs, V.; Lindsay, S.; Aghazadeh, M.; Sangster, C.; Beatty, J. Molecular Diagnosis of Felis catus Gammaherpesvirus 1 (FcaGHV1) Infection in Cats of Known Retrovirus Status with and without Lymphoma. Viruses 2018, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Beatty, J. Viral causes of feline lymphoma: Retroviruses and beyond. Vet. J. 2014, 201, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, M.; Shi, M.; Barrs, V.; McLuckie, A.; Lindsay, S.; Jameson, B.; Hampson, B.; Holmes, E.; Beatty, J. A Novel Hepadnavirus Identified in an Immunocompromised Domestic Cat in Australia. Viruses 2018, 10, 269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Penning, L.C.; Vrieling, H.E.; Brinkhof, B.; Riemers, F.M.; Rothuizen, J.; Rutteman, G.R.; Hazewinkel, H.A.W. A validation of 10 feline reference genes for gene expression measurements in snap-frozen tissues. Vet. Immunol. Immunopathol. 2007, 120, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Damania, B.; Jeong, J.H.; Bowser, B.S.; DeWire, S.M.; Staudt, M.R.; Dittmer, D.P. Comparison of the Rta/Orf50 transactivator proteins of gamma-2-herpesviruses. J. Virol. 2004, 78, 5491–5499. [Google Scholar] [CrossRef] [PubMed]
- Troyer, R.M.; Lee, J.S.; Vuyisich, M.; Chain, P.; Lo, C.-C.; Kronmiller, B.; Bracha, S.; Avery, A.C.; VandeWoude, S. First Complete Genome Sequence of Felis catus Gammaherpesvirus 1. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Giffin, L.; Damania, B. KSHV: Pathways to Tumorigenesis and Persistent Infection. In Advances in Virus Research; Maramorosch, K., Murphy, F.A., Eds.; Elsevier: London, UK, 2014; Volume 88, pp. 111–159. [Google Scholar]
- Chapman, J.R.; Webster, A.C.; Wong, G. Cancer in the Transplant Recipient. Cold Spring Harb. Perspect. Med. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Louwerens, M.; London, C.A.; Pedersen, N.C.; Lyons, L.A. Feline lymphoma in the post-feline leukemia virus era. J. Vet. Intern. Med. 2005, 19, 329–335. [Google Scholar] [PubMed]
- Meichner, K.; Kruse, B.D.; Hirschberger, J.; Hartmann, K. Changes in prevalence of progressive feline leukaemia virus infection in cats with lymphoma in Germany. Vet. Rec. 2012, 171, 348. [Google Scholar] [CrossRef] [PubMed]
- Ballon, G.; Chen, K.; Perez, R.; Tam, W.; Cesarman, E. Kaposi sarcoma herpesvirus (KSHV) vFLIP oncoprotein induces B cell transdifferentiation and tumorigenesis in mice. J. Clin. Investig. 2011, 121, 1141–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fakhari, F.D.; Jeong, J.H.; Kanan, Y.; Dittmer, D.P. The latency-associated nuclear antigen of Kaposi sarcoma-associated herpesvirus induces B cell hyperplasia and lymphoma. J. Clin. Investig. 2006, 116, 735–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariggio, G.; Koch, S.; Schulz, T.F. Kaposi sarcoma herpesvirus pathogenesis. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372. [Google Scholar] [CrossRef] [PubMed]


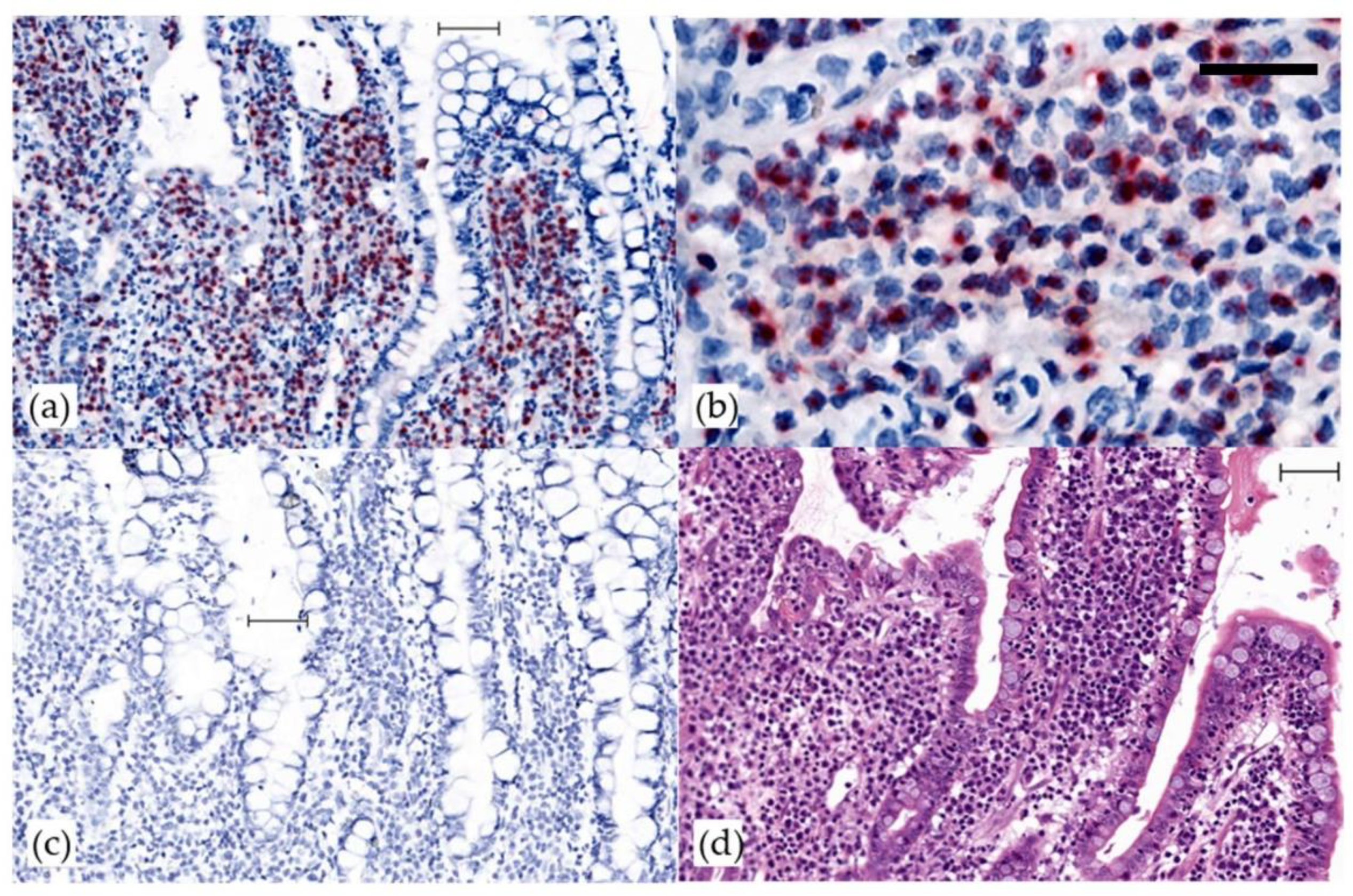{kind=link}
| Cases | NextSeq FcaGHV1 Reads | HiSeq Number of Reads | RT-PCR | |||||
|---|---|---|---|---|---|---|---|---|
| Total | GUSB | FIV | FcaGHV1 | FcaGHV1 ORF50 | FcaGHV1 ORF73 | FcaGHV1 F7 | ||
| 1 | Neg | 294,779,800 | 4149 | 1023 | 0 | Neg | Neg | Neg |
| 2 | 4 (ORF 50) | 338,384,008 | 8070 | 2959 | 0 | POS | POS | POS |
| 3 | - | 288,176,064 | 4152 | 48,270 | 0 | Neg | Neg | Neg |
| 4 | - | 317,655,184 | 6526 | 32 | 0 | POS | Neg | POS |
| 5 | - | 300,408,704 | 1608 | 4090 | 0 | Neg | Neg | Neg |
| 6 1 | - | 321,297,408 | 12,866 | 1636 | 2 (ORF59) | Neg | Neg | POS |
| 7 | - | 337,067,856 | 21,636 | 100 | 0 | Neg | Neg | Neg |
| 8 | Neg | 328,445,328 | 5103 | 2207 | 0 | POS | Neg | POS |
| 9 | - | 313,060,728 | 1410 | 141 | 2 (F10) 2 (F20) 2 (ORF6) | Neg | Neg | Neg |
| 10 | - | 239,825,576 | 1973 | 11 | 0 | Neg | Neg | Neg |
| Oligonucleotide | Sequence | Size (bp) | Tm (°C) |
|---|---|---|---|
| ORF50-F1 | 5′-CCCAGGGCTTTTTGTGTGGA-3′ | 301 | 57 |
| ORF50-R1 | 5′-GGGCTTGACTCATAAGGGCA-3′ | ||
| ORF73-F2 | 5′-CAACTGGGCATTGGCATAC-3′ | 110 | 57 |
| ORF73-R2 | 5′-CCTTAGTTCACCCAACTTGTGC-3′ | ||
| F7-F | 5′-ACTCTGTGTCTGGGAATGTGAC-3′ | 153 | 56 |
| F7-R | 5′-TGGCTTGTGTATATGGCCAGC-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aghazadeh, M.; Shi, M.; Pesavento, P.A.; Durham, A.C.; Polley, T.; Donahoe, S.L.; Troyer, R.M.; Barrs, V.R.; Holmes, E.C.; Beatty, J.A. Transcriptome Analysis and In Situ Hybridization for FcaGHV1 in Feline Lymphoma. Viruses 2018, 10, 464. https://doi.org/10.3390/v10090464
Aghazadeh M, Shi M, Pesavento PA, Durham AC, Polley T, Donahoe SL, Troyer RM, Barrs VR, Holmes EC, Beatty JA. Transcriptome Analysis and In Situ Hybridization for FcaGHV1 in Feline Lymphoma. Viruses. 2018; 10(9):464. https://doi.org/10.3390/v10090464
Chicago/Turabian StyleAghazadeh, Mahdis, Mang Shi, Patricia A. Pesavento, Amy C. Durham, Tamsen Polley, Shannon L. Donahoe, Ryan M. Troyer, Vanessa R. Barrs, Edward C. Holmes, and Julia A. Beatty. 2018. "Transcriptome Analysis and In Situ Hybridization for FcaGHV1 in Feline Lymphoma" Viruses 10, no. 9: 464. https://doi.org/10.3390/v10090464
APA StyleAghazadeh, M., Shi, M., Pesavento, P. A., Durham, A. C., Polley, T., Donahoe, S. L., Troyer, R. M., Barrs, V. R., Holmes, E. C., & Beatty, J. A. (2018). Transcriptome Analysis and In Situ Hybridization for FcaGHV1 in Feline Lymphoma. Viruses, 10(9), 464. https://doi.org/10.3390/v10090464