Virucidal and Synergistic Activity of Polyphenol-Rich Extracts of Seaweeds against Measles Virus

 , ,
, ,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Antiviral Agents
2.1.1. Collection of Seaweeds
2.1.2. Polyphenol-Rich Extracts Isolation
2.2. Cell Line and Virus
2.3. Cytotoxicity Assay
2.4. Antiviral Activity
2.5. Quantitative Real-Time PCR
2.6. Evaluation of PPs Synergy
2.7. Virucidal Assays
2.8. Time of Addition Assays
2.9. Viral Penetration Assays
2.10. Total Phenolic Content (TPC) Evaluation
2.11. High Performance Liquid Chromatography (HPLC) and Mass (MS) Analysis
2.12. Statistical Analysis
3. Results
3.1. Cytotoxicity and Antiviral Activity In Vitro of Polyphenol-Rich Extracts of Seaweeds (PPs)
3.2. Combined Effect of Polyphenol-Rich Extracts (PPs) with Sulphated Polysaccharides (Sps) and Ribavirin
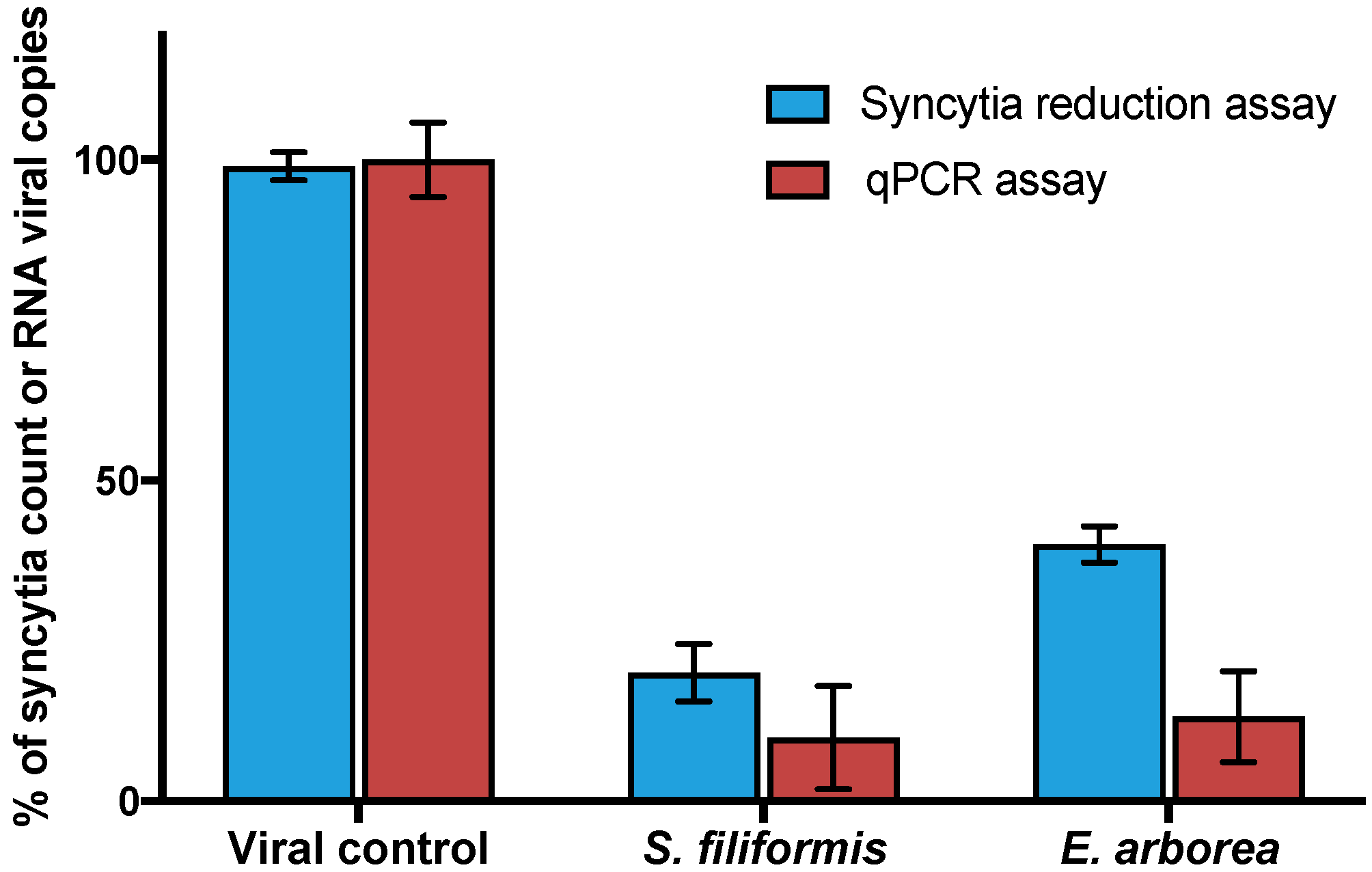
3.3. Virucidal Activity of Polyphenol-Rich Extracts (PPs)
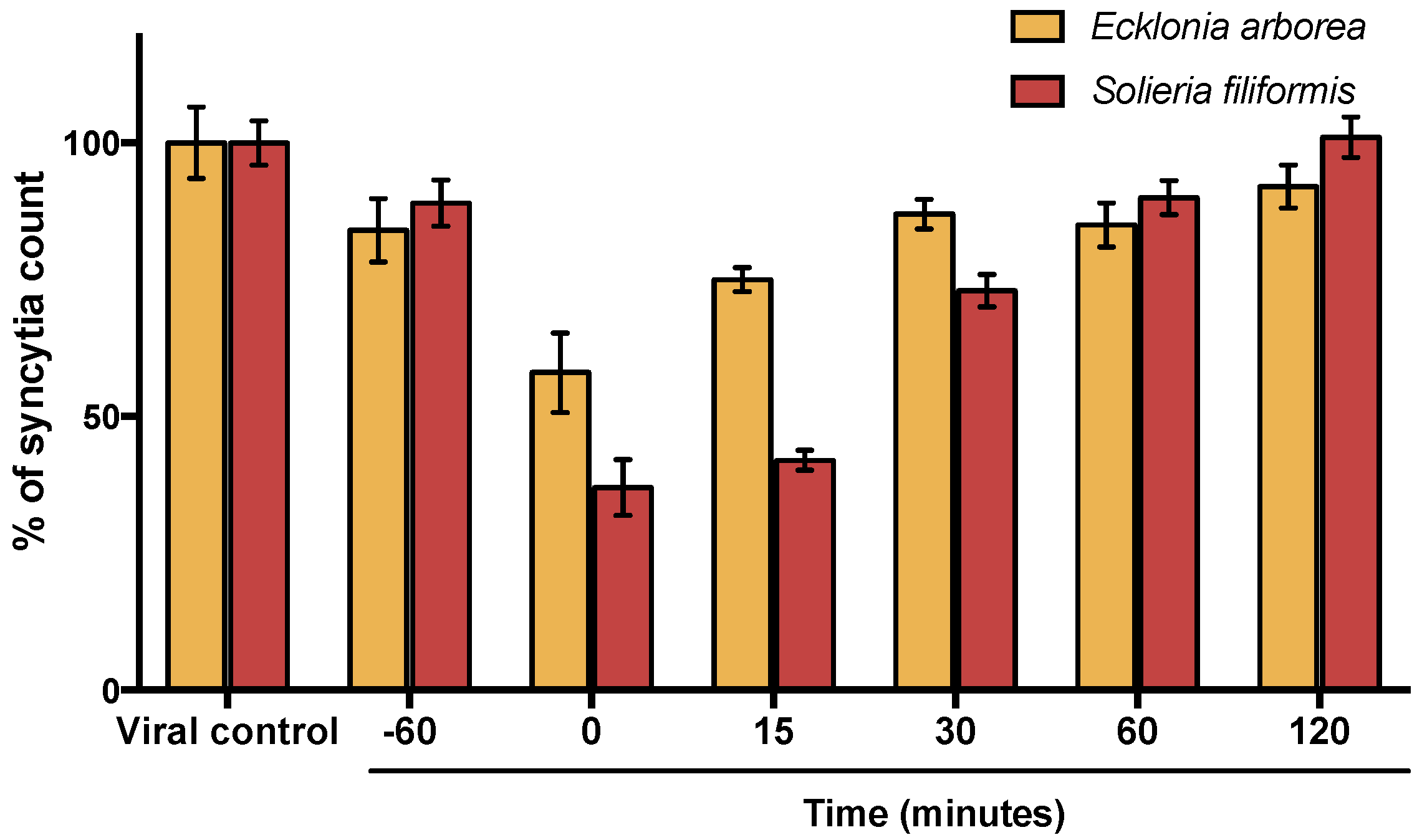
3.4. Effect of Polyphenol-Rich Extracts (PPs) at Different Times of Addition
3.5. Effect of Polyphenol-Rich Extracts (PPs) in Viral Entry
3.6. Estimated Total Phenolic Content (TPC)
3.7. Polyphenol-Rich Extracts (PPs) Characterization by High Performance Liquid Chromatography (HPLC) and Mass (MS) Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Minh, C.; Van Kiem, P.; Hai Dang, N. Marine natural products and their potential application in the future. ASEAN J. Sci. Technol. Dev. 2005, 22, 297–311. [Google Scholar] [CrossRef]
- Gheda, S.F.; El-Adawi, H.I.; El-Deeb, N.M. Antiviral profile of brown and red seaweed polysaccharides against Hepatitis C Virus. Iran J. Pharm. Res. 2016, 15, 483–491. [Google Scholar] [PubMed]
- Chojnacka, K.; Saeid, A.; Witkowska, Z.; Tuhy, Ł. Biologically active compounds in seaweed extracts—The prospects for the application. Open Conf. Proc. J. 2012, 3, 20–28. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falqué, E.; Domínguez, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Xia, Q.; Yang, G.; Zhu, D.; Shao, Y.; Zhang, J.; Cui, Y.; Wang, R.; Zhang, L. The anti-HIV-1 activity of polyphenols from Phyllanthus urinaria and the pharmacokinetics and tissue distribution of its marker compound, gallic acid. J. Tradit. Chin. Med. Sci. 2017, 4, 158–166. [Google Scholar] [CrossRef]
- Yang, Z.F.; Bai, L.P.; Huang, W.B.; Li, X.Z.; Zhao, S.S.; Zhong, N.S.; Jiang, Z.H. Comparison of in vitro antiviral activity of tea polyphenols against influenza A and B viruses and structure-activity relationship analysis. Fitoterapia 2014, 93, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Stockfleth, E.; Meyer, T. The use of sinecatechins (polyphenon E) ointment for treatment of external genital warts. Expert Opin. Biol. Ther. 2012, 12, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Calvo, A.; de Oya, N.J.; Martín-Acebes, M.A.; Garcia-Moruno, E.; Saiz, J.C. Antiviral properties of the natural polyphenols delphinidin and epigallocatechin gallate against the flaviviruses West Nile Virus, Zika Virus, and Dengue Virus. Front. Microbiol. 2017, 8, 1314. [Google Scholar] [CrossRef] [PubMed]
- Date, A.A.; Destache, C.J. Natural polyphenols: Potential in the prevention of sexually transmitted viral infections. Drug Discov. Today 2016, 21, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Orenstein, W.A.; Cairns, L.; Hinman, A.; Nkowane, B.; Olivé, J.M.; Reingold, A.L. Measles and Rubella Global Strategic Plan 2012–2020 midterm review report: Background and summary. Vaccine 2018, 36, A35–A42. [Google Scholar] [CrossRef] [PubMed]
- Teeraananchai, S.; Kerr, S.J.; Amin, J.; Ruxrungtham, K.; Law, M.G. Life expectancy of HIV-positive people after starting combination antiretroviral therapy: A meta-analysis. HIV Med. 2017, 18, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Pires de Mello, C.P.; Drusano, G.L.; Adams, J.R.; Shudt, M.; Kulawy, R.; Brown, A.N. Oseltamivir-zanamivir combination therapy suppresses drug-resistant H1N1 influenza A viruses in the hollow fiber infection model (HFIM) system. Eur. J. Pharm. Sci. 2018, 111, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Morán-Santibañez, K.; Cruz-Suárez, L.E.; Ricque-Marie, D.; Robledo, D.; Freile-Pelegrín, Y.; Peña-Hernández, M.A.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Synergistic effects of sulfated polysaccharides from Mexican seaweeds against Measles virus. BioMed Res. Int. 2016, 2016, 8502123. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Shen, D.; Zhao, S.; Lu, B.; Li, Y.; Zhang, R. Characterization of polyphenols from green tea leaves using a high hydrostatic pressure extraction. Int. J. Pharm. 2009, 382, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.S.; Wagner, R.R. Penetration of Herpes Simplex Virus into Human Epidermoid Cells. Exp. Biol. Med. 1964, 116, 863–868. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Skouta, R.; Morán-Santibañez, K.; Valenzuela, C.A.; Vasquez, A.H.; Fenelon, K. Assessing the Antioxidant Properties of Larrea tridentata Extract as a Potential Molecular Therapy against Oxidative Stress. Molecules 2018, 23, 1826. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Neveu, V.; Perez-Jiménez, J.; Vos, F.; Crespy, V.; Chaffaut, L.; Mennen, L.; Knox, C.; Neveu, V.; Eisner, R.; Cruz, J.; et al. Phenol-Explorer: An online comprehensive database on polyphenol contents in foods. Database 2010, 2010, bap024. [Google Scholar] [CrossRef] [PubMed]
- MassBank of North America. Available online: http://mona.fiehnlab.ucdavis.edu/ (accessed on 27 July 2018).
- Choi, J.-S.; Lee, K.; Lee, B.-B.; Kim, Y.-C.; Kim, Y.D.; Hong, Y.-K.; Cho, K.K.; Choi, I.S. Antibacterial activity of the phlorotannins dieckol and phlorofucofuroeckol-A from Ecklonia cava against Propionibacterium acnes. Bot. Sci. 2014, 92, 425–431. [Google Scholar] [CrossRef]
- Karadeniz, F.; Karagozlu, M.Z.; Kim, S. Antiviral Activities of Marine Algal Extracts. In Marine Algae Extracts; Wiley-VCH: Weinheim, Germany, 2015; pp. 371–380. [Google Scholar]
- Namvar, F.; Mohamad, R.; Baharara, J.; Zafar-Balanejad, S.; Fargahi, F.; Rahman, H.S. Antioxidant, Antiproliferative, and Antiangiogenesis Effects of Polyphenol-Rich Seaweed (Sargassum muticum). BioMed Res. Int. 2013, 2013, 604787. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Gonzalez, R.; Cruz-Suarez, L.E.; Ricque-Marie, D.; Mendoza-Gamboa, E.; Rodriguez-Padilla, C.; Trejo-Avila, L.M. In vitro characterization of the antiviral activity of fucoidan from Cladosiphon okamuranus against Newcastle Disease Virus. Virol. J. 2012, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Briseño, J.A.; Cruz-Suarez, L.E.; Sassi, J.F.; Ricque-Marie, D.; Zapata-Benavides, P.; Mendoza-Gamboa, E.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Sulphated polysaccharides from Ulva clathrata and Cladosiphon okamuranus seaweeds both inhibit viral attachment/entry and cell-cell fusion, in NDV infection. Mar. Drugs 2015, 13, 697–712. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Avila, L.M.; Morales-Martínez, M.E.; Ricque-Marie, D.; Cruz-Suarez, L.E.; Zapata-Benavides, P.; Morán-Santibañez, K.; Rodríguez-Padilla, C. In vitro anti-canine distemper virus activity of fucoidan extracted from the brown alga Cladosiphon okamuranus. Virusdisease 2014, 25, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Wang, A.; Lu, Z.; Qin, C.; Hu, J.; Yin, J. Overview on the antiviral activities and mechanisms of marine polysaccharides from seaweeds. Carbohydr. Res. 2017, 453–454, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Kwak, J. Antiviral phlorotannin from Eisenia bicyclis against human papilloma virus in vitro. Planta Med. 2015, 81, 22. [Google Scholar] [CrossRef]
- Ahn, M.J.; Yoon, K.D.; Min, S.Y.; Lee, J.S.; Kim, J.H.; Kim, T.G.; Kim, S.H.; Kim, N.G.; Huh, H.; Kim, J. Inhibition of HIV-1 reverse transcriptase and protease by phlorotannins from the brown alga Ecklonia cava. Biol. Pharm. Bull. 2004, 27, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Eom, S.H.; Moon, S.Y.; Lee, D.S.; Kim, H.J.; Park, K.; Lee, E.W.; Kim, T.H.; Chung, Y.H.; Lee, M.S.; Kim, Y.M. In vitro antiviral activity of dieckol and phlorofucofuroeckol-A isolated from edible brown alga Eisenia bicyclis against murine norovirus. Algae 2015, 30, 241–246. [Google Scholar] [CrossRef]
- Wright, P.F.; Deatly, A.M.; Karron, R.A.; Belshe, R.B.; Shi, J.R.; Gruber, W.C.; Zhu, Y.; Randolph, V.B. Comparison of results of detection of rhinovirus by PCR and viral culture in human nasal wash specimens from subjects with and without clinical symptoms of respiratory illness. J. Clin. Microbiol. 2007, 45, 2126–2129. [Google Scholar] [CrossRef] [PubMed]
- Bahramsoltani, R.; Sodagari, H.R.; Farzaei, M.H.; Abdolghaffari, A.H.; Gooshe, M.; Rezaei, N. The preventive and therapeutic potential of natural polyphenols on influenza. Expert Rev. Anti-Infect. Ther. 2016, 14, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Al-Saif, S.S.A.; Abdel-Raouf, N.; Aref, I.A. Antibacterial substances from marine algae isolated from Jeddah coast of Red sea, Saudi Arabia. Saudi J. Biol. Sci. 2014, 21, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Namvar, F.; Mohamed, S.; Ghasemi, S.; Behravan, J. Polyphenol-rich seaweed (Eucheuma cottonii) extract suppresses breast tumour via hormone modulation and apoptosis induction. Food Chem. 2012, 130, 376–382. [Google Scholar] [CrossRef]
- El-toumy, S.A.; Salib, J.Y.; El-kashak, W.A.; Marty, C.; Bedoux, G.; Bourgougnon, N. Antiviral effect of polyphenol rich plant extracts on herpes simplex virus type 1. Food Sci. Hum. Wellness 2018, 7, 91–101. [Google Scholar] [CrossRef]
- Amoros, M.; Simóes, C.M.O. Synergistic effect of playones and flavonols against herpes simplex virus type 1 in cell culture. Comparison with the antiviral activity of propolis. J. Nat. Prod. 1992, 55, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Nagavama, K.; Uchida, K. Antioxidant Activity from the Brown of Phlorotannins Isolated Alga Eisenia bicyclis. Nineteenth International Seaweed Symposium. Dev. Appl. Phycol. 1996, 62, 923–926. [Google Scholar]
- Sugiura, Y.; Matsuda, K.; Okamoto, T.; Kakinuma, M.; Amano, H. Anti-allergic effects of the brown alga Eisenia arborea on Brown Norway rats. Fish. Sci. 2008, 74, 180–186. [Google Scholar] [CrossRef]
- Kwon, H.J.; Ryu, Y.B.; Kim, Y.M.; Song, N.; Kim, C.Y.; Rho, M.C.; Jeong, J.H.; Cho, K.O.; Lee, W.S.; Park, S.J. In vitro antiviral activity of phlorotannins isolated from Ecklonia cava against porcine epidemic diarrhea coronavirus infection and hemagglutination. Bioorg. Med. Chem. 2013, 21, 4706–4713. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract or Compound a | CC50 (μg/mL) b | IC50 (μg/mL) c | SI d |
|---|---|---|---|
| Macrocystis pyrifera (Phaeophyceae) PP | >1500 | 3 ± 0.33 | >500 |
| Ecklonia arborea (formerly Eisenia arborea, Phaeophyceae) PP | >1500 | 2.6 ± 0.28 | >576.9 |
| Silvetia compressa (formerly Pelvetia compressa, Phaeophyceae) PP | 165.03 ± 9.7 | 1.86 ± 0.61 | 306.9 |
| Ulva intestinalis (Chlorophyta) PP | >1500 | 3.1 ± 0.23 | >483.9 |
| Solieria filiformis (Rhodophyta) PP | >1500 | 0.4 ± 0.11 | >3750 |
| Ribavirin | 405 ± 4.1 | 35 ± 1.8 | 11.57 |
| Combination | Compound Concentration (μg/mL) | % Relative Syncytia of Each Combination | SD | CI | Description | |||
|---|---|---|---|---|---|---|---|---|
| Polyphenol PPE or PPS * | Polysaccharides SPE or SPS * | |||||||
| Ecklonia arborea | Solieria filiformis | Ecklonia arborea | Solieria filiformis | |||||
| PPE50-SPE50 | 2.6 | - | 0.275 | - | 38.8 | 3.5 | 0.79 | Synergism |
| PPE25-SPE50 | 0.03 | - | 0.275 | - | 49.1 | 6.8 | 1.75 | Antagonism |
| PPS50-SPE50 | - | 0.4 | 0.275 | - | 55.1 | 8.1 | 4.4 | Antagonism |
| PPS25-SPE50 | - | 0.07 | 0.275 | - | 36.7 | 2.9 | 0.65 | Synergism |
| PPE50/SPE25 | 2.6 | - | 0.01 | - | 40 | 6.4 | 0.19 | Synergism |
| PPE25/SPE25 | 0.03 | - | 0.01 | - | 73.5 | 2.6 | 3.6 | Antagonism |
| PPS50/SPE25 | - | 0.4 | 0.01 | - | 44.9 | 3.3 | 0.4 | Synergism |
| PPS25/SPE25 | - | 0.07 | 0.01 | - | 57.14 | 6.7 | 0.4 | Synergism |
| PPE50/SPS50 | 2.6 | - | - | 0.985 | 22.5 | 6.2 | 0.57 | Synergism |
| PPE25/SPS50 | 0.03 | - | - | 0.985 | 34.5 | 8.3 | 1.85 | Antagonism |
| PPS50/SPS50 | - | 0.4 | - | 0.985 | 28.8 | 1.5 | 1.37 | Antagonism |
| PPS25/SPS50 | - | 0.07 | - | 0.985 | 36.7 | 2 | 2.25 | Antagonism |
| PPE50/SPS25 | 2.6 | - | - | 0.01 | 30.6 | 5.3 | 0.03 | Synergism |
| PPE25/SPS25 | 0.03 | - | - | 0.01 | 42.9 | 1.1 | 0.4 | Synergism |
| PPS50/SPS25 | - | 0.4 | - | 0.01 | 33.3 | 1.8 | 0.03 | Synergism |
| PPS25/SPS25 | - | 0.07 | - | 0.01 | 38.8 | 2.5 | 0.11 | Synergism |
| Combination | Concentration (μg/mL) | % Relative Syncytia of Each Combination | SD | CI | Description | ||
|---|---|---|---|---|---|---|---|
| Polyphenols PPE or PPS * | Ribavirin | ||||||
| Ecklonia arborea | Solieria filiformis | ||||||
| PPE50/R50 | 2.6 | - | 35 | 4 | 0.5 | 9.1 | Antagonism |
| PPE50/R25 | 2.6 | - | 16 | 32 | 1.1 | 2.6 | Antagonism |
| PPE25/R50 | 0.03 | - | 35 | 20 | 1.8 | 1.3 | Antagonism |
| PPE25/R25 | 0.03 | - | 16 | 33.2 | 1.5 | 1.2 | Antagonism |
| PPS50/R50 | - | 0.4 | 35 | 13.6 | 1.2 | 3.6 | Antagonism |
| PPS50/R25 | - | 0.4 | 16 | 36.4 | 1.9 | 2.5 | Antagonism |
| PPS25/R50 | - | 0.07 | 35 | 24.3 | 2.1 | 2.2 | Antagonism |
| PPS25/R25 | - | 0.07 | 16 | 30.3 | 1.8 | 1.3 | Antagonism |
| Fraction | Retention Time (min) | Major Fragment Ions m/z (% Base Peak) | Tentative Identification |
|---|---|---|---|
| F1 | 0.802 | 298.339 (100), 136.989 (92.09), 150.115 (91.36) | 5-Methylthioadenosine a,b Inosine a,b, L-Methionine a |
| F2 | 8.109 | 230.243 (100) | Terbutylazine a,b |
| F3 | 8.620 | 507.299 (100), 683.430 (98.13), 639.405 (97.21), 551.349 (95.74), 595.376 (91.57) | Quercetin 3-(6-O-acetyl-beta-glucoside) a,b, Methyllycaconitine a,b, Demethoxycentaureidin 7-O-rutinosideab, Quercetin 3-O-(6′′-malonyl-glucoside) a,b,c, Kaempferol-3-O-rutinoside a,b,c. |
| F4 | 9.382 | 301.134 (100) | Kaempferidea |
| F5 | 10.716 | 413. 256 (100) | 7-acetyloxy-2-(3,4-diacetyloxyphenyl)-4-oxochromen-5-yl acetate a |
| F6 | 10.860 | 264.237 (100) | Abscisic acid a,b |
| Fraction | Retention Time (min) | Major Fragment Ions m/z (% Base Peak) | Tentative Identification |
|---|---|---|---|
| F1 | 1.052 | 365. 125(100), 205.064 (98.23), 601.138 (40) | Cellobiose a,b, Tryptophan a Phlorofucofuroeckol-B c |
| F2 | 5.982 | 602.140 (100), 268.993 (73.23), 230.244 (68.75) | Phlorofucofuroeckol-B c, Formononetin a,b, Apigenin 7-O-glucoside a |
| F3 | 6.195 | 413.259 (100), 327.195 (67.57) | Leganin a Feruloyl tartaric acid a |
| F4 | 6.733 | 205.074 (100) | Tryptophan a |
| F5 | 8.844 | 601.139 (100) | Phlorofucofuroeckol-B c |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morán-Santibañez, K.; Peña-Hernández, M.A.; Cruz-Suárez, L.E.; Ricque-Marie, D.; Skouta, R.; Vasquez, A.H.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Virucidal and Synergistic Activity of Polyphenol-Rich Extracts of Seaweeds against Measles Virus. Viruses 2018, 10, 465. https://doi.org/10.3390/v10090465
Morán-Santibañez K, Peña-Hernández MA, Cruz-Suárez LE, Ricque-Marie D, Skouta R, Vasquez AH, Rodríguez-Padilla C, Trejo-Avila LM. Virucidal and Synergistic Activity of Polyphenol-Rich Extracts of Seaweeds against Measles Virus. Viruses. 2018; 10(9):465. https://doi.org/10.3390/v10090465
Chicago/Turabian StyleMorán-Santibañez, Karla, Mario A. Peña-Hernández, Lucia Elizabeth Cruz-Suárez, Denis Ricque-Marie, Rachid Skouta, Abimael H. Vasquez, Cristina Rodríguez-Padilla, and Laura M. Trejo-Avila. 2018. "Virucidal and Synergistic Activity of Polyphenol-Rich Extracts of Seaweeds against Measles Virus" Viruses 10, no. 9: 465. https://doi.org/10.3390/v10090465
APA StyleMorán-Santibañez, K., Peña-Hernández, M. A., Cruz-Suárez, L. E., Ricque-Marie, D., Skouta, R., Vasquez, A. H., Rodríguez-Padilla, C., & Trejo-Avila, L. M. (2018). Virucidal and Synergistic Activity of Polyphenol-Rich Extracts of Seaweeds against Measles Virus. Viruses, 10(9), 465. https://doi.org/10.3390/v10090465