Identification of Novel Subcellular Localization and Trafficking of HIV-1 Nef Variants from Reference Strains G (F1.93.HH8793) and H (BE.93.VI997)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Pseudovirus Production and Viral Transduction
2.3. Immunoblotting
2.4. Flow Cytometry
2.5. Transfections and Microscopy
2.6. Quantitative RT-PCR
3. Results
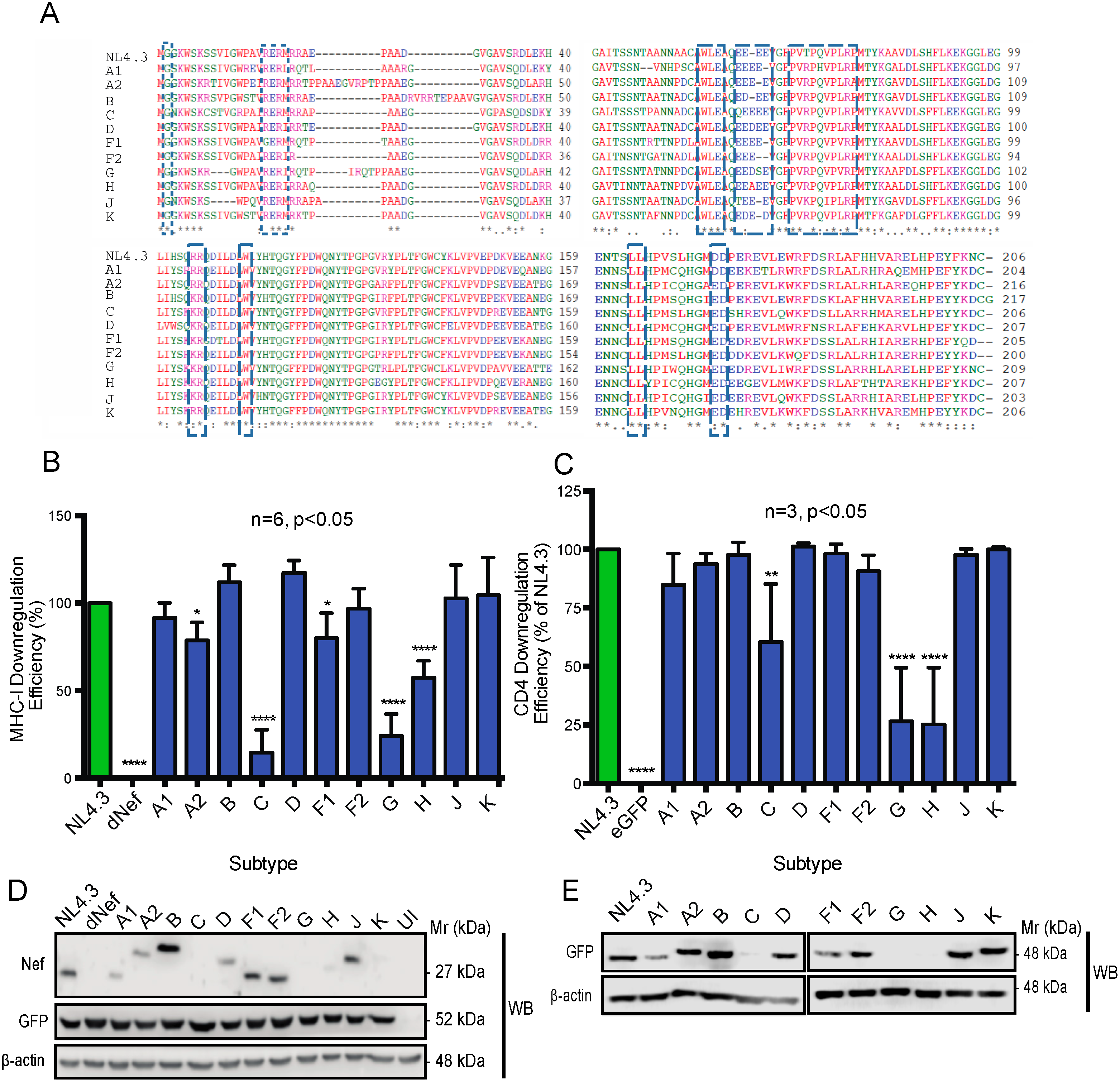
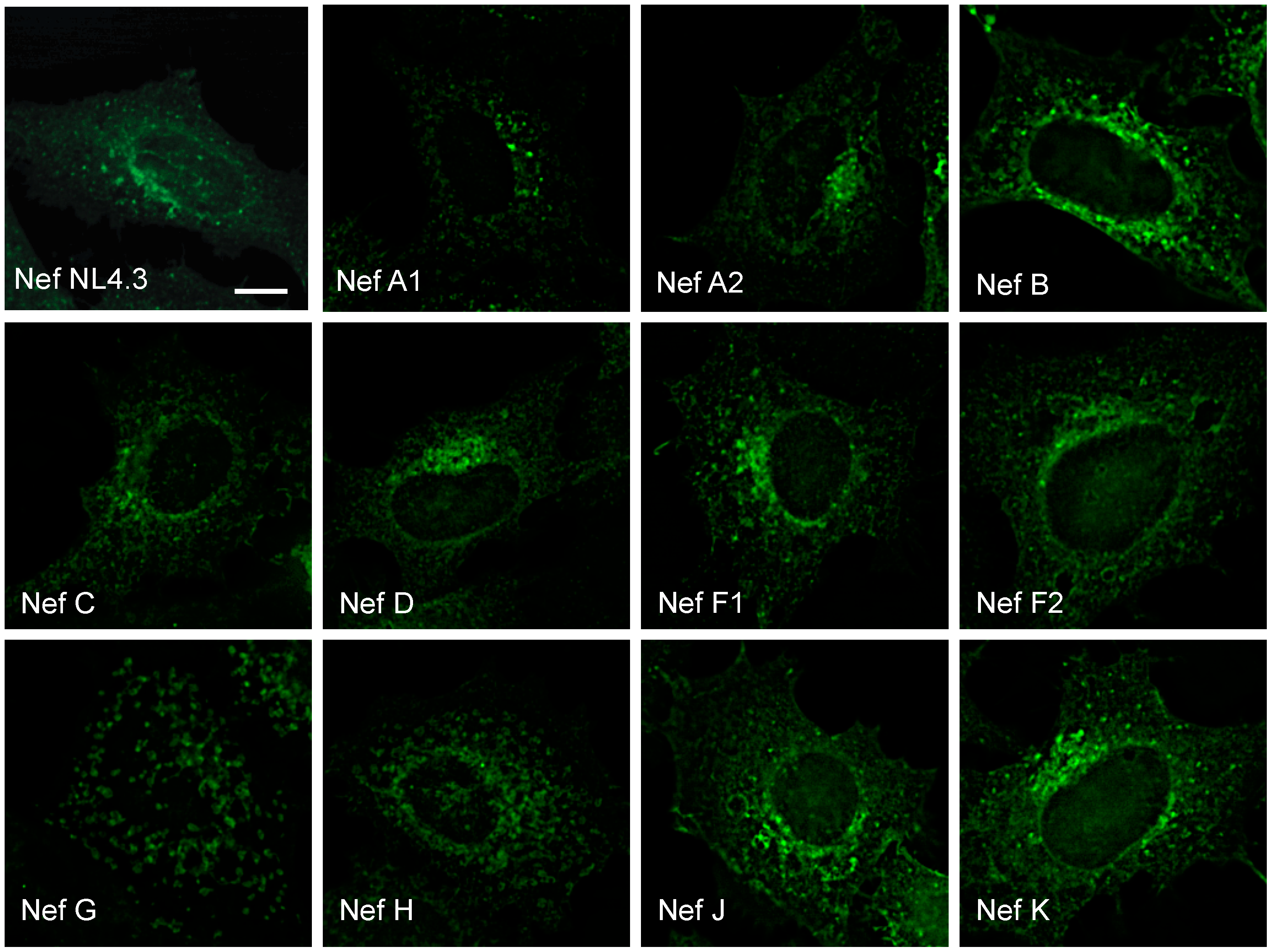
3.1. Nef from Reference Strains of HIV-1 Group-M Subtypes Exhibit Differential Functionality, Expression, and Subcellular Distribution
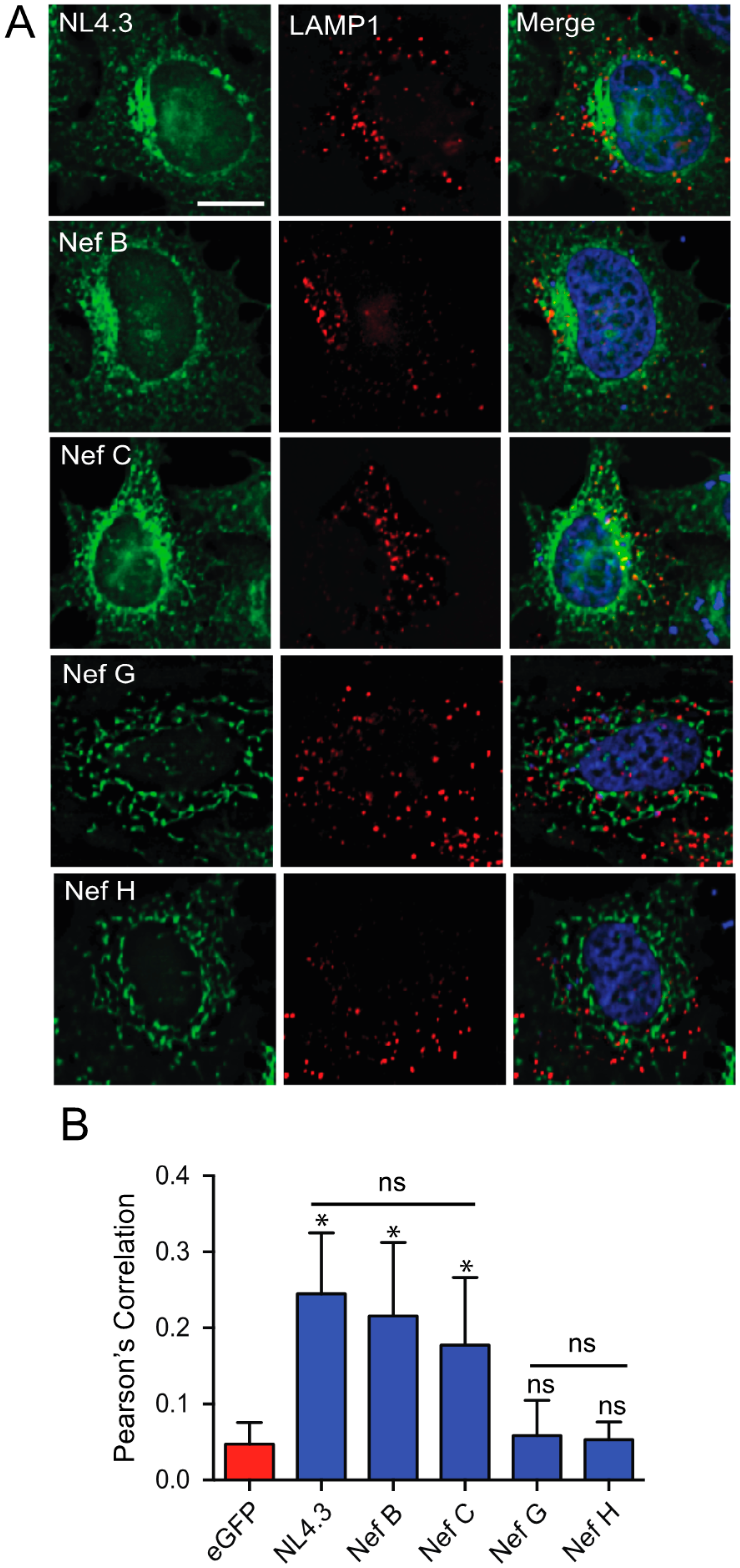
3.2. Nef Isolates from HIV-1 Reference Strains of Group-M Subtypes G and H Do Not Display Selective Colocalization with the Lysosome
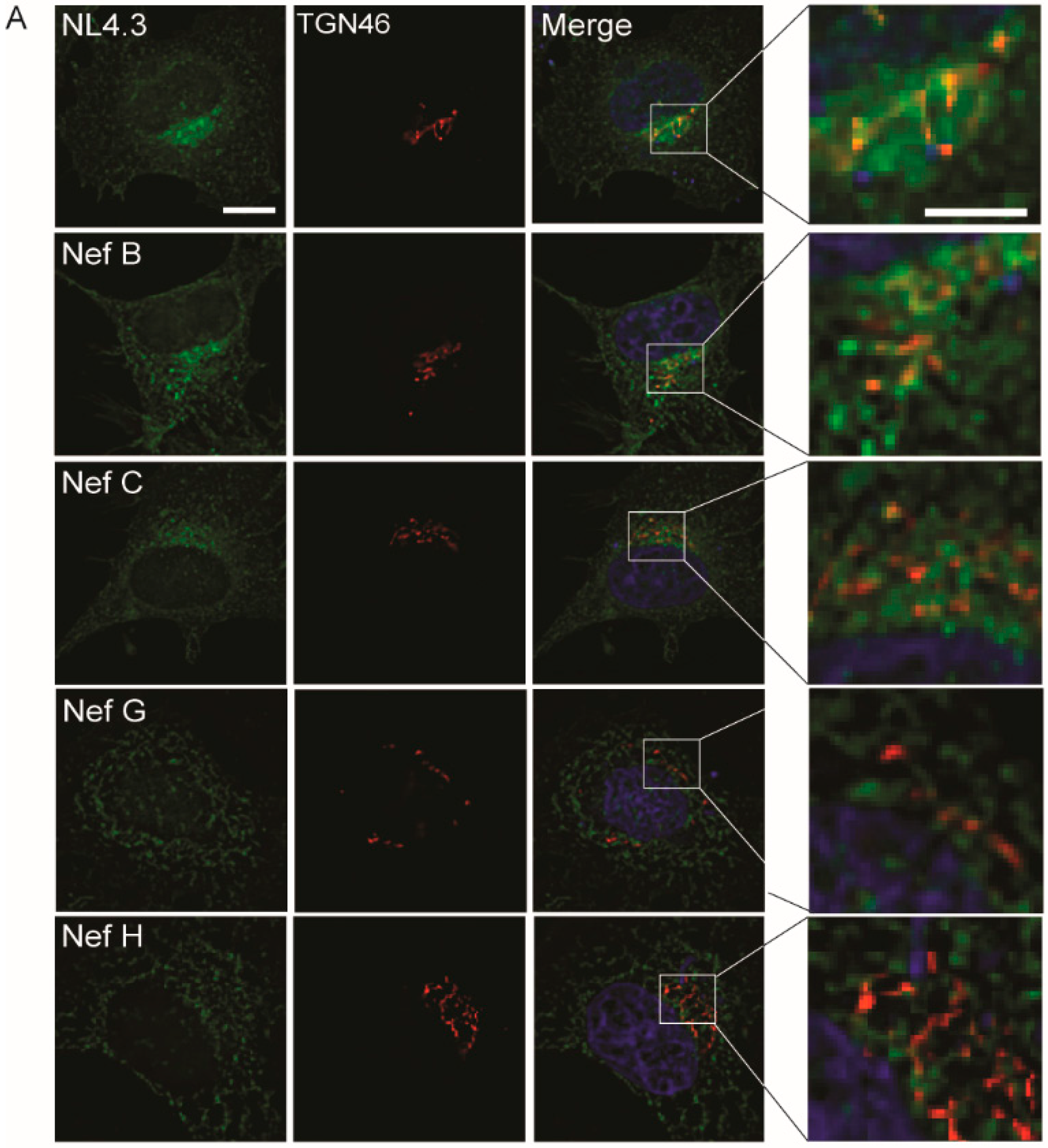
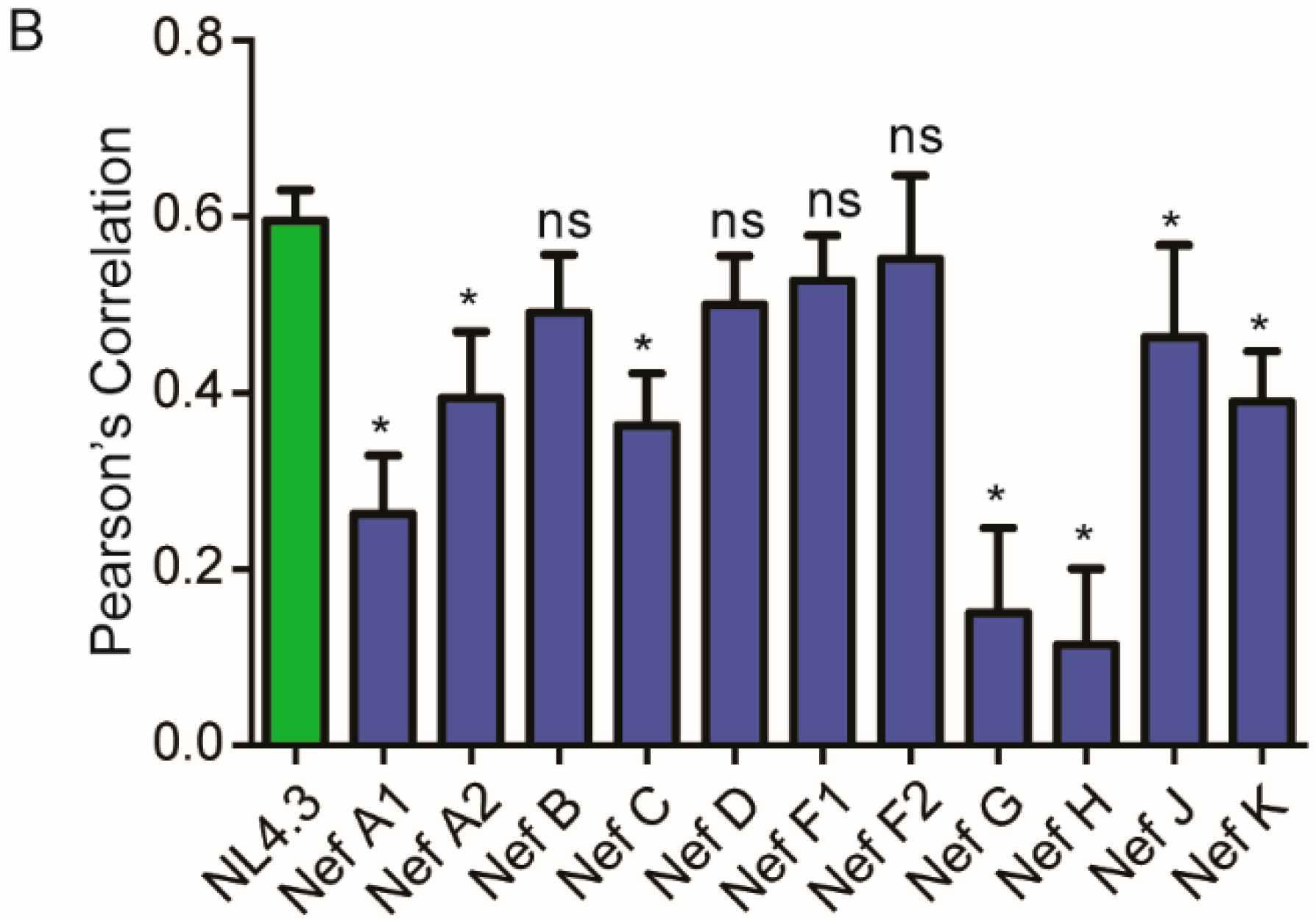
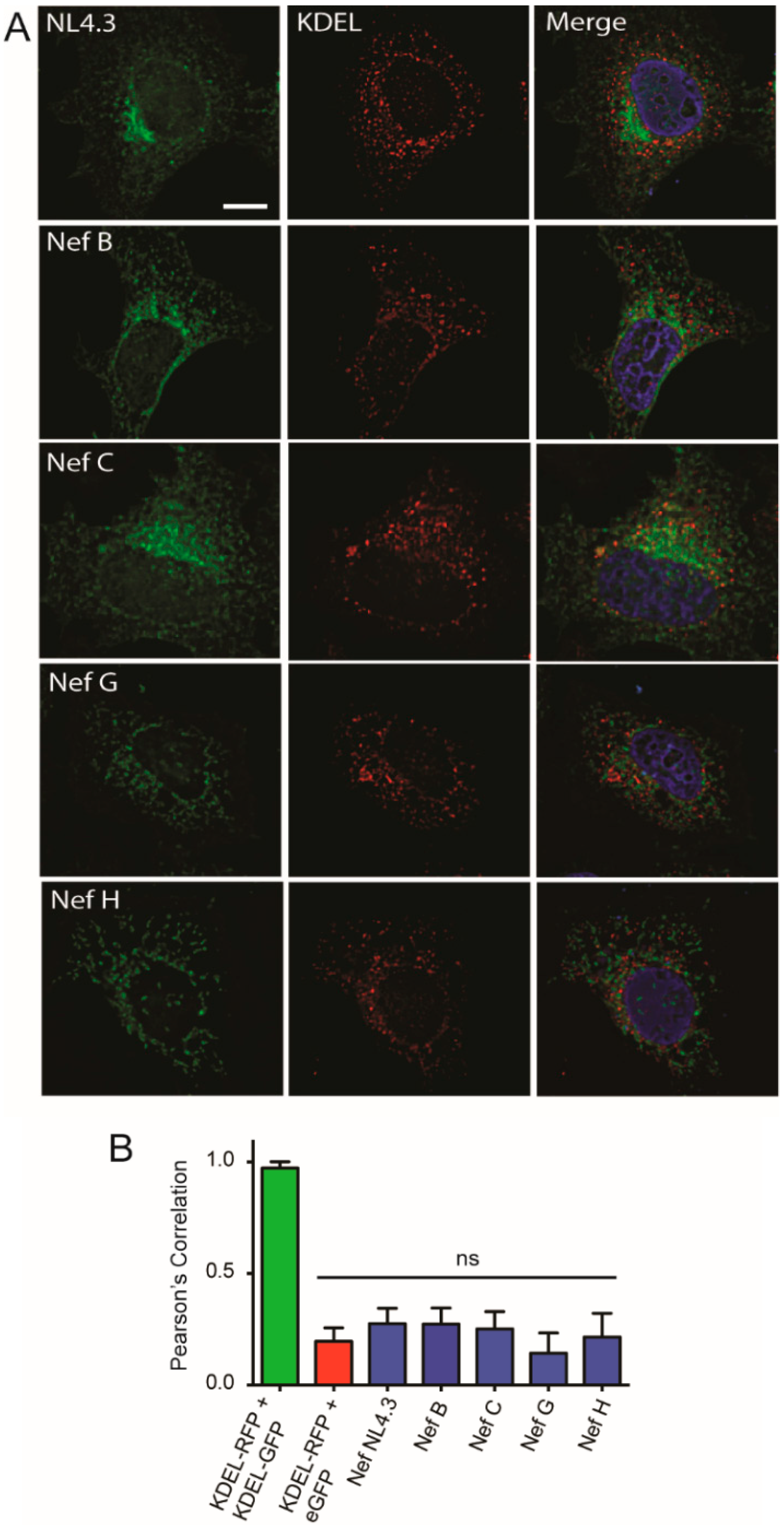
3.3. Nef from HIV-1 Group-M Subtypes G (F1.93.HH8793) and H (BE.93.VI997) Reference Strains Do Not Display Differential Colocalization with the Endoplasmic Reticulum
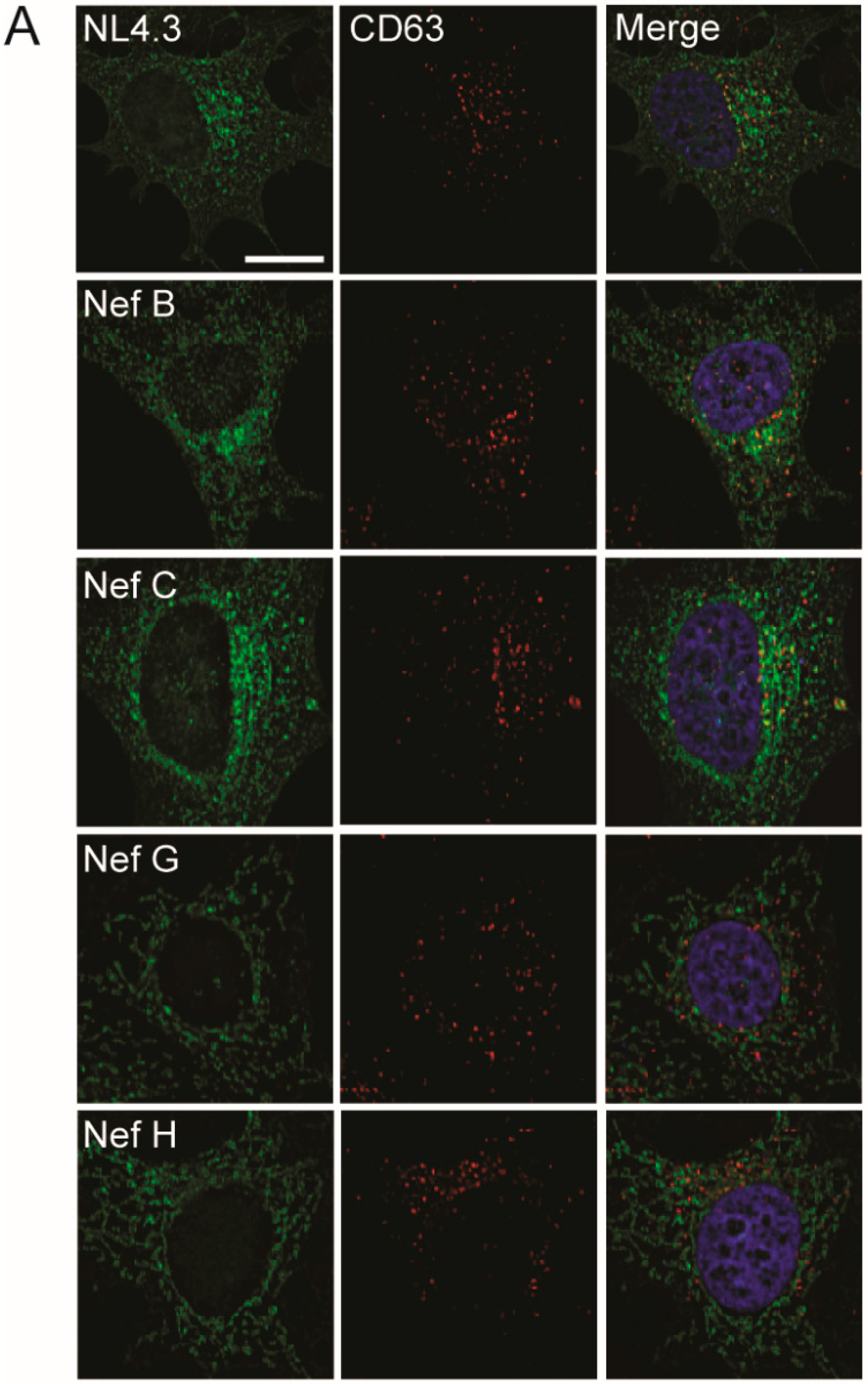
3.4. Nef from Reference Strains of HIV-1 Group-M Subtypes G or H Do Not Colocalize with CD63
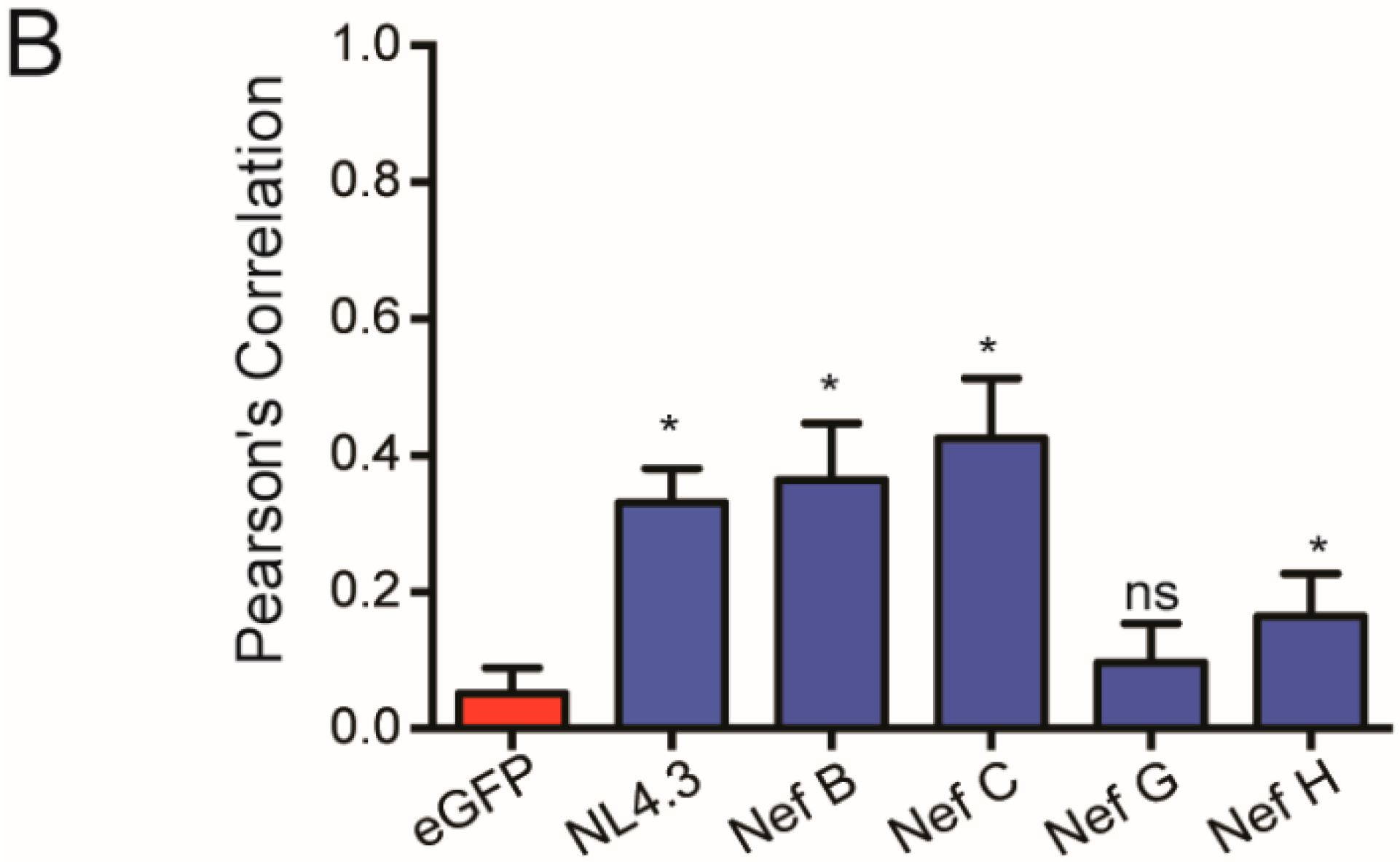
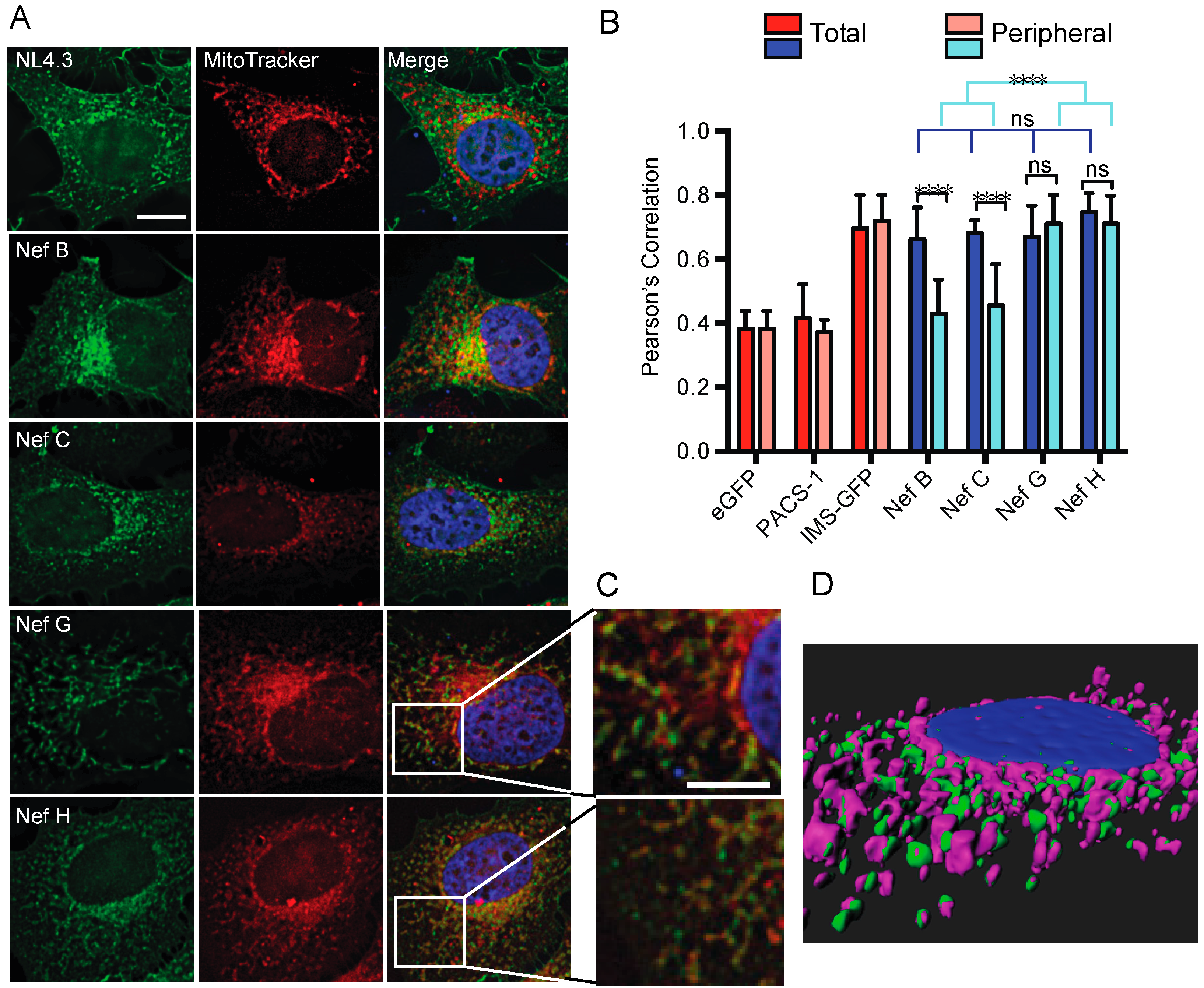
3.5. Nef from Reference Strains of HIV-1 Group-M Subtypes G (F1.93.HH8793) and H (BE.93.VI997) Selectively Colocalize with Mitochondria
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Robertson, D.; Anderson, J.; Bradac, J.; Carr, J.; Foley, B.; Funkhouser, R.; Gao, F.; Hahn, B.; Kalish, M.; Kuiken, C. HIV-1 Nomenclature Proposal. Science 2000, 288, 55–56. [Google Scholar] [CrossRef] [PubMed]
- Hemelaar, J.; Gouws, E.; Ghys, P.D.; Osmanov, S. Global Trends in Molecular Epidemiology of HIV-1 During 2000–2007. AIDS 2011, 25, 679. [Google Scholar] [CrossRef] [PubMed]
- Bobkova, M. Current Status of HIV-1 Diversity and Drug Resistance Monitoring in the Former USSR. AIDS Rev. 2013, 15, 204–212. [Google Scholar] [PubMed]
- Vidal, N.; Peeters, M.; Mulanga-Kabeya, C.; Nzilambi, N.; Robertson, D.; Ilunga, W.; Sema, H.; Tshimanga, K.; Bongo, B.; Delaporte, E. Unprecedented Degree of Human Immunodeficiency Virus Type 1 (HIV-1) Group M Genetic Diversity in the Democratic Republic of Congo Suggests that the HIV-1 Pandemic Originated in Central Africa. J. Virol. 2000, 74, 10498–10507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lihana, R.W.; Ssemwanga, D.; Abimiku, A.l.; Ndembi, N. Update on Hiv-1 Diversity in Africa: A Decade in Review. AIDS Rev. 2012, 14, 83–100. [Google Scholar] [PubMed]
- Bhargava, M.; Cajas, J.M.; Wainberg, M.A.; Klein, M.B.; Pai, N.P. Do HIV-1 Non-B Subtypes Differentially Impact Resistance Mutations and Clinical Disease Progression in Treated Populations? Evidence from a Systematic Review. J. Int. AIDS Soc. 2014, 17, 18944. [Google Scholar] [CrossRef] [PubMed]
- Kiwanuka, N.; Laeyendecker, O.; Robb, M.; Kigozi, G.; Arroyo, M.; McCutchan, F.; Eller, L.A.; Eller, M.; Makumbi, F.; Birx, D. Effect of Human Immunodeficiency Virus Type 1 (HIV-1) Subtype on Disease Progression in Persons from Rakai, Uganda, with Incident HIV-1 Infection. J. Infect. Dis. 2008, 197, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Amornkul, P.N.; Karita, E.; Kamali, A.; Rida, W.N.; Sanders, E.J.; Lakhi, S.; Price, M.A.; Kilembe, W.; Cormier, E.; Anzala, O. Disease Progression by Infecting HIV-1 Subtype in a Seroconverter Cohort in Sub-Saharan Africa. AIDS 2013, 27, 2775. [Google Scholar] [CrossRef] [PubMed]
- Bousheri, S.; Burke, C.; Ssewanyana, I.; Harrigan, R.; Martin, J.; Bangsberg, D.R.; Cao, H. Infection with Different HIV Subtypes Is Associated with CD4 Activation-Associated Dysfunction and Apoptosis. J. Acquir. Immune Defic. Syndr. 2009, 52, 548. [Google Scholar] [CrossRef] [PubMed]
- Chaix, M.-L.; Seng, R.; Frange, P.; Tran, L.; Avettand-Fenoël, V.; Ghosn, J.; Reynes, J.; Yazdanpanah, Y.; Raffi, F.; Goujard, C. Increasing HIV-1 Non-B Subtype Primary Infections in Patients in France and Effect of HIV Subtypes on Virological and Immunological Responses to Combined Antiretroviral Therapy. Clin. Infect. Dis. 2012, 56, 880–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherrer, A.U.; Ledergerber, B.; von Wyl, V.; Böni, J.; Yerly, S.; Klimkait, T.; Bürgisser, P.; Rauch, A.; Hirschel, B.; Cavassini, M. Improved Virological Outcome in White Patients Infected with HIV-1 Non-B Subtypes Compared to Subtype B. Clin. Infect. Dis. 2011, 53, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Greenough, T.C.; Brettler, D.B.; Kirchhoff, F.; Alexander, L.; Alexander, R.C.; O’Brien, S.J.; Somasundaran, M.; Luzuriaga, K.; Sullivan, J.L. Long-Term Nonprogressive Infection with Human Immunodeficiency Virus Type 1 in a Hemophilia Cohort. J. Infect. Dis. 1999, 180, 1790–1802. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, F.; Greenough, T.C.; Brettler, D.B.; Sullivan, J.L.; Desrosiers, R.C. Absence of Intact Nef Sequences in a Long-Term Survivor with Nonprogressive HIV-1 Infection. New Engl. J. Med. 1995, 332, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Salvi, R.; Garbuglia, A.R.; Di Caro, A.; Pulciani, S.; Montella, F.; Benedetto, A. Grossly Defective Nef Gene Sequences in a Human Immunodeficiency Virus Type 1-Seropositive Long-Term Nonprogressor. J. Virol. 1998, 72, 3646–3657. [Google Scholar] [PubMed]
- Kestier, H.W., III; Ringler, D.J.; Mori, K.; Panicali, D.L.; Sehgal, P.K.; Daniel, M.D.; Desrosiers, R.C. Importance of the Nef Gene for Maintenance of High Virus Loads and for Development of AIDS. Cell 1991, 65, 651–662. [Google Scholar] [CrossRef]
- Dirk, B.S.; van Nynatten, L.R.; Dikeakos, J.D. Where in the Cell Are You? Probing HIV-1 Host Interactions through Advanced Imaging Techniques. Viruses 2016, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, E.N.; Dikeakos, J.D. HIV-1 Nef: A Master Manipulator of the Membrane Trafficking Machinery Mediating Immune Evasion. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2015, 1850, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Dirk, B.S.; Pawlak, E.N.; Johnson, A.L.; Van Nynatten, L.R.; Jacob, R.A.; Heit, B.; Dikeakos, J.D. HIV-1 Nef Sequesters MHC-I Intracellularly by Targeting Early Stages of Endocytosis and Recycling. Sci. Rep. 2016, 6, 37021. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, E.N.; Dirk, B.S.; Jacob, R.A.; Johnson, A.L.; Dikeakos, J.D. The HIV-1 Accessory Proteins Nef and Vpu Downregulate Total and Cell Surface CD28 in CD4+ T Cells. Retrovirology 2018, 15, 6. [Google Scholar] [CrossRef] [PubMed]
- Atkins, K.M.; Thomas, L.; Youker, R.T.; Harriff, M.J.; Pissani, F.; You, H.; Thomas, G. HIV-1 Nef Binds PACS-2 to Assemble a Multikinase Cascade That Triggers Major Histocompatibility Complex Class I (MHC-I) Down-Regulation Analysis Using Short Interfering RNA and Knock-out Mice. J. Biol. Chem. 2008, 283, 11772–11784. [Google Scholar] [CrossRef] [PubMed]
- Trible, R.P.; Emert-Sedlak, L.; Smithgall, T.E. HIV-1 Nef Selectively Activates Src Family Kinases Hck, Lyn, and C-Src through Direct Sh3 Domain Interaction. J. Biol. Chem. 2006, 281, 27029–27038. [Google Scholar] [CrossRef] [PubMed]
- Prévost, J.; Richard, J.; Medjahed, H.; Alexander, A.; Jones, J.; Kappes, J.C.; Ochsenbauer, C.; Finzi, A. Incomplete Downregulation of CD4 Expression Affects HIV-1 Env Conformation and Antibody-Dependent Cellular Cytotoxicity Responses. J. Virol. 2018, 92, e00484-18. [Google Scholar] [CrossRef] [PubMed]
- Dikeakos, J.D.; Thomas, L.; Kwon, G.; Elferich, J.; Shinde, U.; Thomas, G. An Interdomain Binding Site on HIV-1 Nef Interacts with PACS-1 and PACS-2 on Endosomes to Down-Regulate MHC-I. Mol. Biol. Cell 2012, 23, 2184–2197. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.; Lindwasser, O.W.; Smith, W.J.; Hurley, J.H.; Bonifacino, J.S. Downregulation of CD4 by Human Immunodeficiency Virus Type 1 Nef Is Dependent on Clathrin and Involves Direct Interaction of Nef with the AP2 Clathrin Adaptor. J. Virol. 2007, 81, 3877–3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veillette, M.; Coutu, M.; Richard, J.; Batraville, L.-A.; Dagher, O.; Bernard, N.; Tremblay, C.; Kaufmann, D.E.; Roger, M.; Finzi, A. The HIV-1 Gp120 CD4-Bound Conformation Is Preferentially Targeted by Antibody-Dependent Cellular Cytotoxicity-Mediating Antibodies in Sera from HIV-1-Infected Individuals. J. Virol. 2015, 89, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Alsahafi, N.; Ding, S.; Richard, J.; Markle, T.; Brassard, N.; Walker, B.; Lewis, G.K.; Kaufmann, D.E.; Brockman, M.A.; Finzi, A. Nef Proteins from HIV-1 Elite Controllers Are Inefficient at Preventing Antibody-Dependent Cellular Cytotoxicity. J. Virol. 2016, 90, 2993–3002. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Park, S.Y.; Bonifacino, J.S.; Hurley, J.H. How HIV-1 Nef Hijacks the AP-2 Clathrin Adaptor to Downregulate CD4. Elife 2014, 3, e01754. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.-J.; Cai, C.Y.; Zhang, X.; Zhang, H.-T.; Hirst, J.A.; Burakoff, S.J. HIV Nef-Mediated CD4 Down-Regulation Is Adaptor Protein Complex 2 Dependent. J. Immunol. 2005, 175, 3157–3164. [Google Scholar] [CrossRef] [PubMed]
- Piguet, V.; Gu, F.; Foti, M.; Demaurex, N.; Gruenberg, J.; Carpentier, J.-L.; Trono, D. Nef-Induced CD4 Degradation: A Diacidic-Based Motif in Nef Functions as a Lysosomal Targeting Signal through the Binding of Β-Cop in Endosomes. Cell 1999, 97, 63–73. [Google Scholar] [CrossRef]
- Benichou, S.; Bomsel, M.; Bodeus, M.; Durand, H.; Doute, M.; Letourneur, F.; Camonis, J.; Benarous, R. Physical Interaction of the HIV-1 Nef Protein with Beta-Cop, a Component of Non-Clathrin-Coated Vesicles Essential for Membrane Traffic. J. Biol. Chem. 1994, 269, 30073–30076. [Google Scholar] [PubMed]
- Schaefer, M.R.; Wonderlich, E.R.; Roeth, J.F.; Leonard, J.A.; Collins, K.L. HIV-1 Nef Targets MHC-I and CD4 for Degradation via a Final Common Β-Cop–Dependent Pathway in T Cells. PLoS Pathog. 2008, 4, e1000131. [Google Scholar] [CrossRef] [PubMed]
- Dirk, B.S.; Jacob, R.A.; Johnson, A.L.; Pawlak, E.N.; Cavanagh, P.C.; van Nynatten, L.; Haeryfar, S.M.; Dikeakos, J.D. Viral Bimolecular Fluorescence Complementation: A Novel Tool to Study Intracellular Vesicular Trafficking Pathways. PLoS ONE 2015, 10, e0125619. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.L.; Dirk, B.S.; Coutu, M.; Haeryfar, S.M.; Arts, E.J.; Finzi, A.; Dikeakos, J.D. A Highly Conserved Residue in HIV-1 Nef Alpha Helix 2 Modulates Protein Expression. mSphere 2016, 1, e00288-16. [Google Scholar] [CrossRef] [PubMed]
- Heit, B.; Yeung, T.; Grinstein, S. Changes in Mitochondrial Surface Charge Mediate Recruitment of Signaling Molecules during Apoptosis. Am. J. Physiol.-Cell Physiol. 2010, 300, C33–C41. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to Imagej: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671. [Google Scholar] [CrossRef] [PubMed]
- Bolte, S.; Cordelieres, F. A Guided Tour into Subcellular Colocalization Analysis in Light Microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef] [PubMed]
- Piguet, V.; Schwartz, O.; Le Gall, S.; Trono, D. The Downregulation of CD4 and MHC-I by Primate Lentiviruses: A Paradigm for the Modulation of Cell Surface Receptors. Immunol. Rev. 1999, 168, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.A.; daSilva, L.L. HIV-1 Nef: Taking Control of Protein Trafficking. Traffic 2016, 17, 976–996. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A New Bioinformatics Analysis Tools Framework at EMBL–EBI. Nucleic Acids Res. 2010, 38, W695–W699. [Google Scholar] [CrossRef] [PubMed]
- Shugars, D.C.; Smith, M.S.; Glueck, D.H.; Nantermet, P.V.; Seillier-Moiseiwitsch, F.; Swanstrom, R. Analysis of Human Immunodeficiency Virus Type 1 Nef Gene Sequences Present in Vivo. J. Virol. 1993, 67, 4639–4650. [Google Scholar] [PubMed]
- Arold, S.T.; Baur, A.S. Dynamic Nef and Nef Dynamics: How Structure Could Explain the Complex Activities of This Small HIV Protein. Trends Biochem. Sci. 2001, 26, 356–363. [Google Scholar] [CrossRef]
- Collins, D.R.; Collins, K.L. HIV-1 Accessory Proteins Adapt Cellular Adaptors to Facilitate Immune Evasion. PLoS Pathog. 2014, 10, e1003851. [Google Scholar] [CrossRef] [PubMed]
- Rohrer, J.; Schweizer, A.; Russell, D.; Kornfeld, S. The Targeting of LAMP1 to Lysosomes Is Dependent on the Spacing of Its Cytoplasmic Tail Tyrosine Sorting Motif Relative to the Membrane. J. Cell Biol. 1996, 132, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Seguin, S.; Morelli, F.; Vinet, J.; Amore, D.; de Biasi, S.; Poletti, A.; Rubinsztein, D.; Carra, S. Inhibition of Autophagy, Lysosome and Vcp Function Impairs Stress Granule Assembly. Cell Death Differ. 2014, 21, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, P.; Ron, D. The Unfolded Protein Response: From Stress Pathway to Homeostatic Regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Lewis, M.J.; Pelham, H.R. Ligand-Induced Redistribution of a Human KDEL Receptor from the Golgi Complex to the Endoplasmic Reticulum. Cell 1992, 68, 353–364. [Google Scholar] [CrossRef]
- Mathivanan, S.; Ji, H.; Simpson, R.J. Exosomes: Extracellular Organelles Important in Intercellular Communication. J. Proteomics 2010, 73, 1907–1920. [Google Scholar] [CrossRef] [PubMed]
- Mukerji, J.; Olivieri, K.C.; Misra, V.; Agopian, K.A.; Gabuzda, D. Proteomic Analysis of HIV-1 Nef Cellular Binding Partners Reveals a Role for Exocyst Complex Proteins in Mediating Enhancement of Intercellular Nanotube Formation. Retrovirology 2012, 9, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenassi, M.; Cagney, G.; Liao, M.; Vaupotič, T.; Bartholomeeusen, K.; Cheng, Y.; Krogan, N.J.; Plemenitaš, A.; Peterlin, B.M. HIV Nef Is Secreted in Exosomes and Triggers Apoptosis in Bystander CD4+ T Cells. Traffic 2010, 11, 110–122. [Google Scholar] [CrossRef] [PubMed]
- McNamara, R.P.; Costantini, L.M.; Myers, T.A.; Schouest, B.; Maness, N.J.; Griffith, J.D.; Damania, B.A.; MacLean, A.G.; Dittmer, D.P. Nef Secretion into Extracellular Vesicles or Exosomes Is Conserved across Human and Simian Immunodeficiency Viruses. mBio 2018, 9, e02344-17. [Google Scholar] [CrossRef] [PubMed]
- Logozzi, M.; de Milito, A.; Lugini, L.; Borghi, M.; Calabro, L.; Spada, M.; Perdicchio, M.; Marino, M.L.; Federici, C.; Iessi, E. High Levels of Exosomes Expressing CD63 and Caveolin-1 in Plasma of Melanoma Patients. PLoS ONE 2009, 4, e5219. [Google Scholar] [CrossRef] [PubMed]
- Chazotte, B. Labeling Mitochondria with Mitotracker Dyes. Cold Spring Harb. Protoc. 2011, 2011, pdb-prot5648. [Google Scholar] [CrossRef] [PubMed]
- Wan, L.; Molloy, S.S.; Thomas, L.; Liu, G.; Xiang, Y.; Rybak, S.L.; Thomas, G. PACS-1 Defines a Novel Gene Family of Cytosolic Sorting Proteins Required for Trans-Golgi Network Localization. Cell 1998, 94, 205–216. [Google Scholar] [CrossRef]
- Piguet, V.; Wan, L.; Borel, C.; Mangasarian, A.; Demaurex, N.; Thomas, G.; Trono, D. HIV-1 Nef Protein Binds to the Cellular Protein PACS-1 to Downregulate Class I Major Histocompatibility Complexes. Nat. Cell Biol. 2000, 2, 163. [Google Scholar] [CrossRef] [PubMed]
- Sica, V.; Galluzzi, L.; Bravo-San Pedro, J.M.; Izzo, V.; Maiuri, M.C.; Kroemer, G. Organelle-Specific Initiation of Autophagy. Mol. Cell 2015, 59, 522–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Klionsky, D.J. Mitochondria Removal by Autophagy. Autophagy 2011, 7, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Rowland, A.A.; Voeltz, G.K. Endoplasmic Reticulum–Mitochondria Contacts: Function of the Junction. Nat. Rev. Mol. Cell Biol. 2012, 13, 607. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Wen, H.; Yu, Y.; Taxman, D.J.; Zhang, L.; Widman, D.G.; Swanson, K.V.; Wen, K.-W.; Damania, B.; Moore, C.B. The Mitochondrial Proteins Nlrx1 and Tufm Form a Complex That Regulates Type I Interferon and Autophagy. Immunity 2012, 36, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Vance, J.E. MAM (Mitochondria-Associated Membranes) in Mammalian Cells: Lipids and Beyond. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2014, 1841, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Williamson, C.D.; Colberg-Poley, A.M. Access of Viral Proteins to Mitochondria via Mitochondria-Associated Membranes. Rev. Med. Virol. 2009, 19, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Missiroli, S.; Patergnani, S.; Caroccia, N.; Pedriali, G.; Perrone, M.; Previati, M.; Wieckowski, M.R.; Giorgi, C. Mitochondria-Associated Membranes (MAMS) and Inflammation. Cell Death Dis. 2018, 9, 329. [Google Scholar] [CrossRef] [PubMed]
- Castanier, C.; Arnoult, D. Mitochondrial Localization of Viral Proteins as a Means to Subvert Host Defense. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2011, 1813, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-Y.; Chiang, S.-F.; Lin, T.-Y.; Chiou, S.-H.; Chow, K.-C. HIV-1 Vpr Triggers Mitochondrial Destruction by Impairing Mfn2-Mediated ER-Mitochondria Interaction. PLoS ONE 2012, 7, e33657. [Google Scholar] [CrossRef] [PubMed] [Green Version]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Nynatten, L.R.; Johnson, A.L.; Dirk, B.S.; Pawlak, E.N.; Jacob, R.A.; Haeryfar, S.M.M.; Dikeakos, J.D. Identification of Novel Subcellular Localization and Trafficking of HIV-1 Nef Variants from Reference Strains G (F1.93.HH8793) and H (BE.93.VI997). Viruses 2018, 10, 493. https://doi.org/10.3390/v10090493
Van Nynatten LR, Johnson AL, Dirk BS, Pawlak EN, Jacob RA, Haeryfar SMM, Dikeakos JD. Identification of Novel Subcellular Localization and Trafficking of HIV-1 Nef Variants from Reference Strains G (F1.93.HH8793) and H (BE.93.VI997). Viruses. 2018; 10(9):493. https://doi.org/10.3390/v10090493
Chicago/Turabian StyleVan Nynatten, Logan R., Aaron L. Johnson, Brennan S. Dirk, Emily N. Pawlak, Rajesh Abraham Jacob, S. M. Mansour Haeryfar, and Jimmy D. Dikeakos. 2018. "Identification of Novel Subcellular Localization and Trafficking of HIV-1 Nef Variants from Reference Strains G (F1.93.HH8793) and H (BE.93.VI997)" Viruses 10, no. 9: 493. https://doi.org/10.3390/v10090493
APA StyleVan Nynatten, L. R., Johnson, A. L., Dirk, B. S., Pawlak, E. N., Jacob, R. A., Haeryfar, S. M. M., & Dikeakos, J. D. (2018). Identification of Novel Subcellular Localization and Trafficking of HIV-1 Nef Variants from Reference Strains G (F1.93.HH8793) and H (BE.93.VI997). Viruses, 10(9), 493. https://doi.org/10.3390/v10090493