Pathogen Risk Analysis for Wild Amphibian Populations Following the First Report of a Ranavirus Outbreak in Farmed American Bullfrogs (Lithobates catesbeianus) from Northern Mexico

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
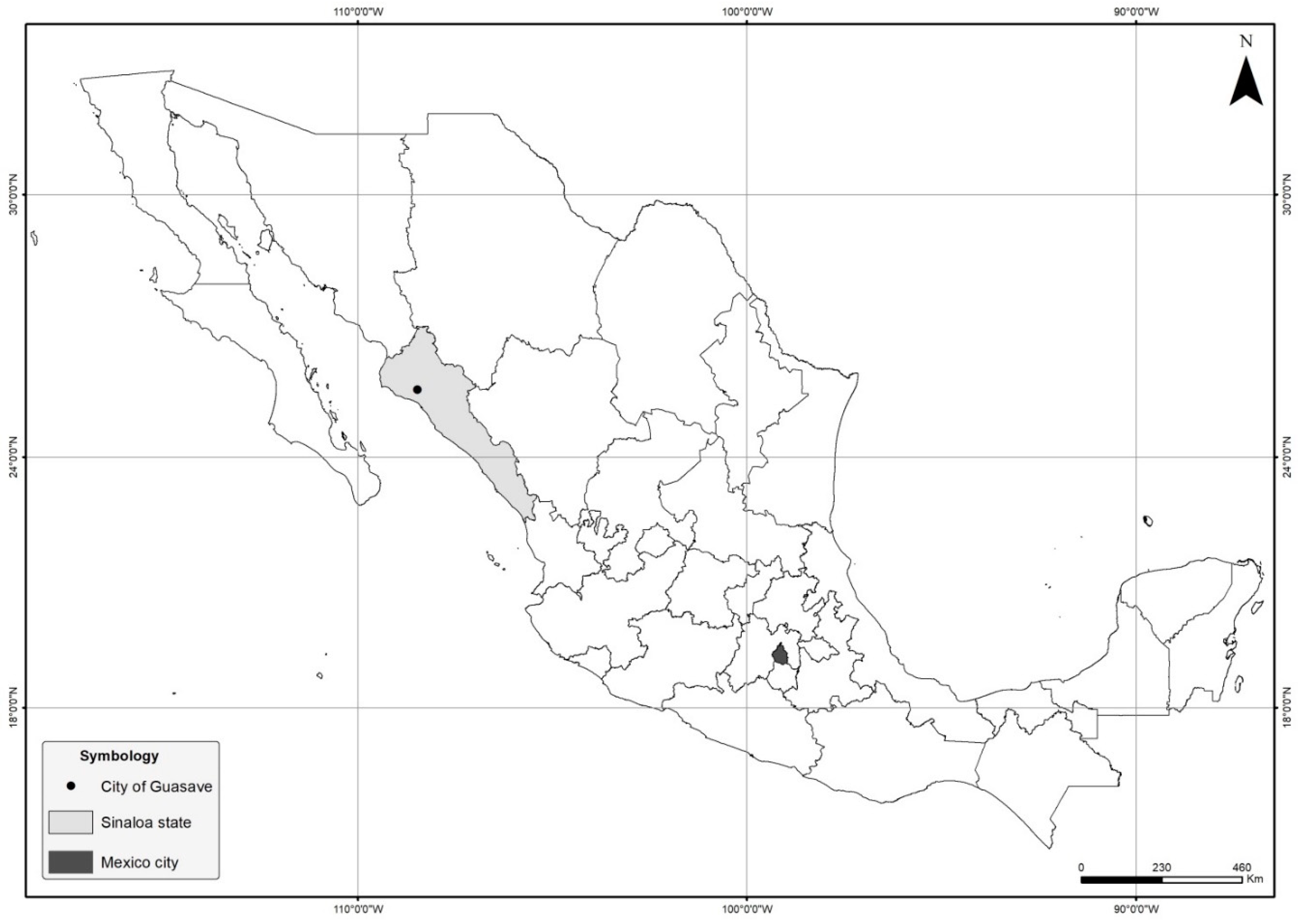
2.1. General Information on the Rana-Culture Facility
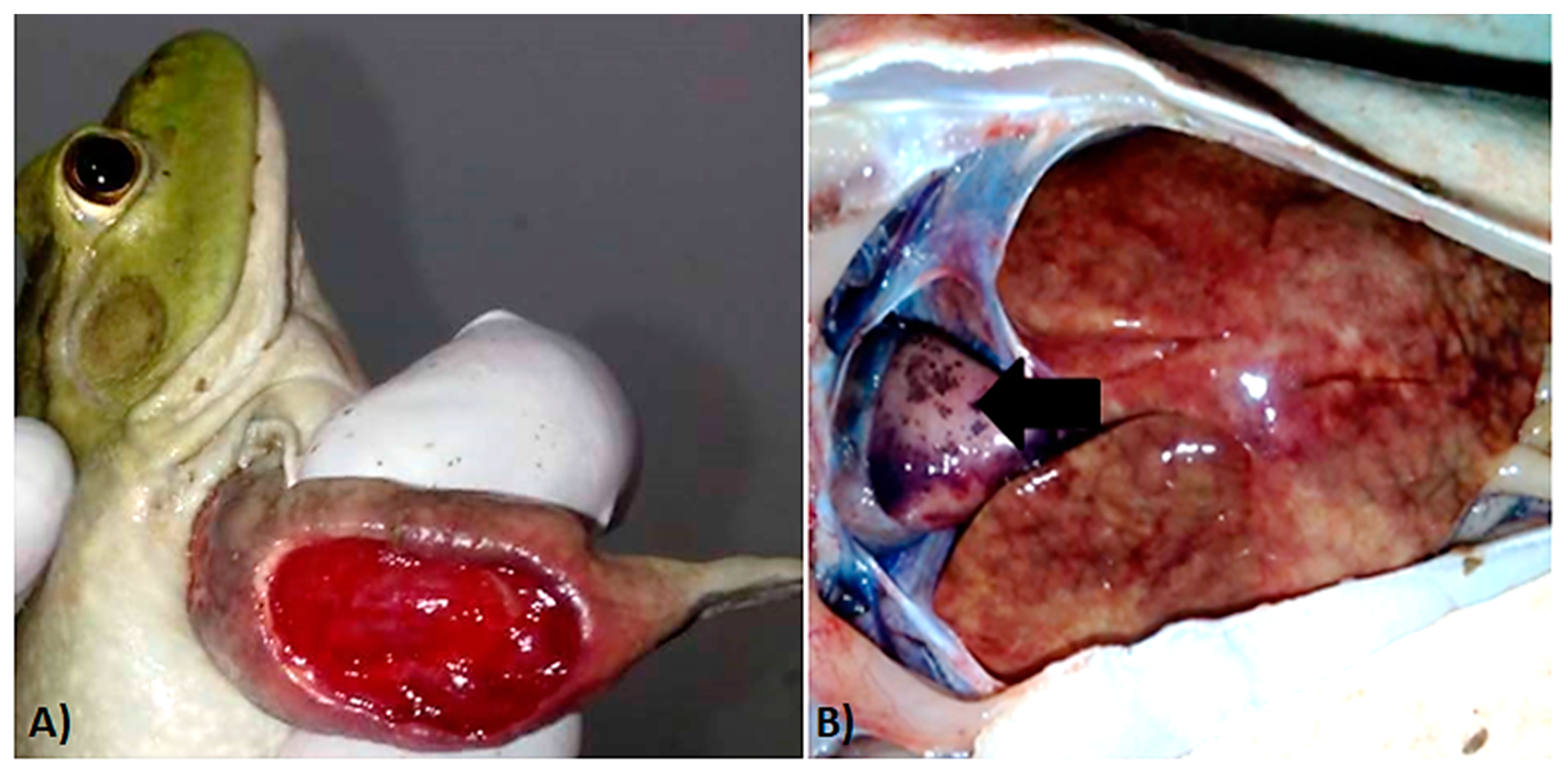
2.2. Necropsies and Sample Collection
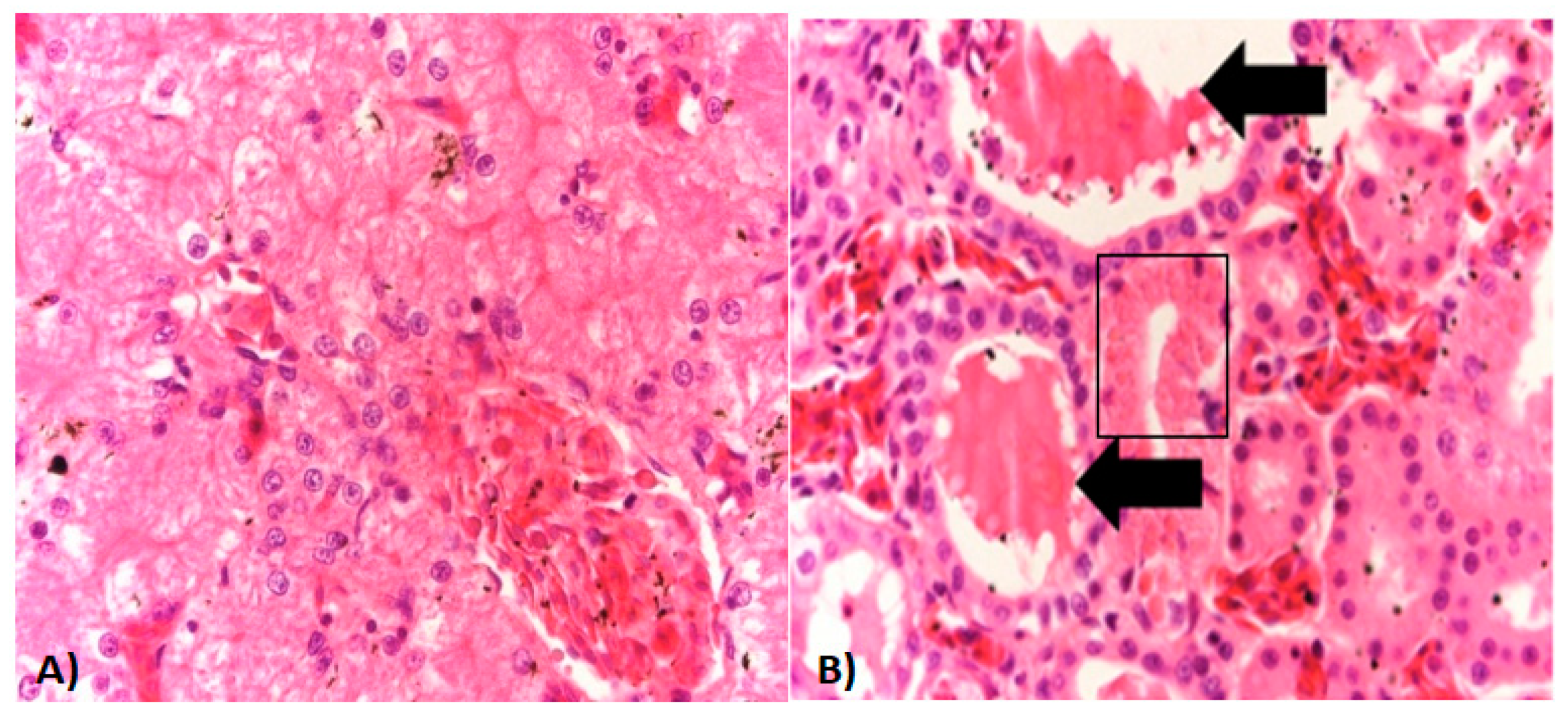
2.3. Histopathology and Immunohistochemistry
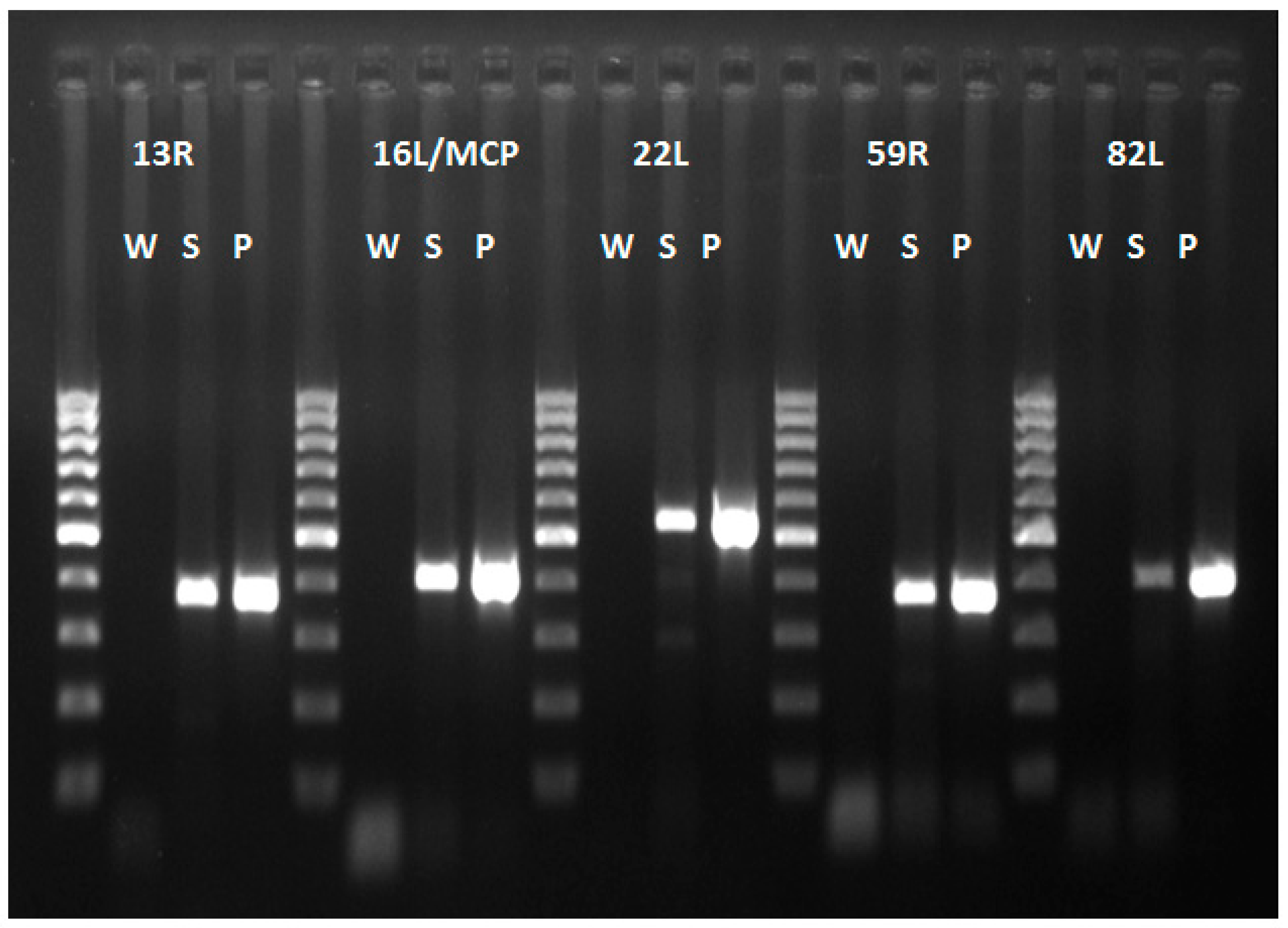
2.4. DNA Extraction, PCR and Sequencing
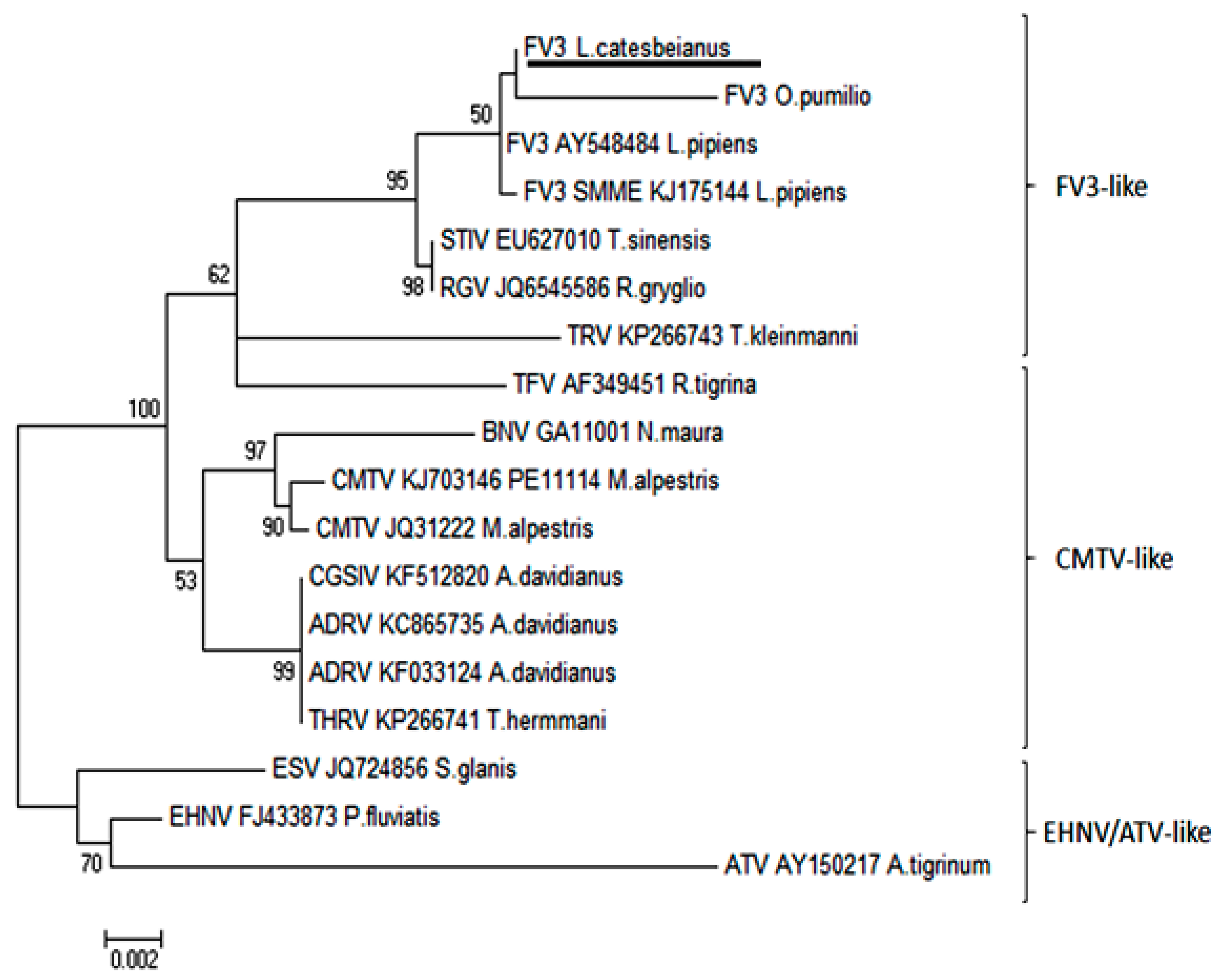
2.5. Phylogenetic Analysis
2.6. Risk Classification Using the PANDORA + Protocol
2.7. Level of Wild Species Threat
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- IUCN 2017. The IUCN Red List of Threatened Species. Version 2017-3. Available online: http://www.iucnredlist.org (accessed on 5 December 2017).
- Bower, D.S.; Lips, K.R.; Schwarzkopf, L.; Georges, A.; Clulow, S. Amphibians on the brink. Science 2017, 357, 454–455. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, A.R.; Kiesecker, J.M. Complexity in conservation: Lessons from the global decline of amphibian populations. Ecol. Lett. 2002, 5, 597–608. [Google Scholar] [CrossRef]
- Chinchar, V.G. Ranaviruses (family Iridoviridae): Emerging cold-blooded killers. Arch. Virol. 2002, 147, 447–470. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Almeralla, C.; Burrowes, P.; Parra-Olea, G. Chytridiomycosis in amphibians from Mexico: A revision. Rev. Mex. Biodivers. 2015, 86, 238–248. [Google Scholar] [CrossRef]
- Duffus, A.L.J.; Waltzek, T.B. Distribution and Host Range of Ranaviruses. In Ranaviruses: Lethal Pathogenics of Ectothermic Vertebrates, 1st ed.; Gray, M.J., Chinchar, G.V., Eds.; Springer: New York, NY, USA, 2015; Volume 1, pp. 9–57. ISBN 978-3-319-13755-1. [Google Scholar]
- Epstein, B.; Storfer, A. Comparative genomics of an emerging amphibian virus. G3 Genes Genom. Genet. 2016, 6, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Wheelwright, N.T.; Gray, M.J.; Hill, R.D.; Miller, D.L. Sudden mass die-off of large population of wood frog (Lithobates sylvaticus) tadpoles in Maine, USA, likely due to ranavirus. Herpetol. Rev. 2014, 45, 240–242. [Google Scholar] [CrossRef]
- Balseiro, A.; Dalton, K.; del Cerro, A.; Marquez, I.; Cunningham, A.A.; Parra, F.; Prieto, J.M.; Casais, R. Pathology, isolation and molecular characterisation of a ranavirus from the common midwife toad Alytes obstetricans on the Iberian Peninsula. Dis. Aquat. Org. 2009, 84, 95–104. [Google Scholar] [CrossRef]
- Kik, M.; Martel, A.; van der Sluijs, A.S.; Pasmans, F.; Wohlsein, P.; Gröne, A.; Rijks, J.M. Ranavirus-associated mass mortality in wild amphibians, The Netherlands, 2010: A first report. Vet. J. 2011, 190, 284–286. [Google Scholar] [CrossRef]
- Miaud, C.; Pozet, F.; Gaudin, N.C.G.; Martel, A.; Pasmans, F.; Labrut, S. Ranavirus causes mass die-offs of alpine amphibians in the Southwestern Alps, France. J. Wildl. Dis. 2016, 52, 242–252. [Google Scholar] [CrossRef]
- He, J.G.; Lu, L.; Deng, M.; He, H.H.; Weng, S.P.; Wang, X.H.; Zhou, S.Y.; Long, Q.X.; Wang, X.Z.; Chan, S.M. Sequence analysis of the complete genome of an iridovirus isolated from tiger frog. Virology 2002, 292, 185–197. [Google Scholar] [CrossRef]
- Lei, X.Y.; Ou, T.; Zhu, R.L.; Zhang, Q.J. Sequencing and analysis of the complete genome of Rana grylio virus (RGV). Arch. Virol. 2012, 157, 1559–1564. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Wang, K.Y.; Zhou, Z.Y.; Li, C.W.; Wang, J.; He, M.; Yin, Z.Q.; Lai, W.M. First report of a ranavirus-associated with morbidity and mortality in farmed Chinese giant salamanders (Andrias davidianus). J. Comp. Pathol. 2011, 141, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Marschang, R.E.; Becher, P.; Posthaus, H.; Wild, P.; Thiel, H.J.; Müller-Doblies, U.; Kalet, E.F.; Bacciarini, L.N. Isolation and characterization of an iridovirus isolated from Hermann’s tortoises (Testudo hermanni). Arch. Virol. 1999, 144, 1909–1922. [Google Scholar] [CrossRef] [PubMed]
- Whittington, R.J.; Kearns, C.; Hyatt, A.D.; Hengstberger, S.; Rutzou, T. Spread of Epizootic hematopoietic necrosis virus (EHNV) in red fin perch (Perca fluviatilis) in southern Australia. Aust. Vet. J. 1996, 73, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Stöhr, A.C.; Hoffmann, A.; Papp, T.; Robert, N.; Pruvost, N.B.M.; Reyer, H.U.; Marschang, R.E. Long-term study of an infection with ranaviruses in a group of edible frogs (Pelophylax kl. esculentus) and partial characterization of two viruses based on four genomic regions. Vet. J. 2013, 197, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.M.; Berlanga-Robles, C.A.; Ruiz-Luna, A. High amphibian diversity related to unexpected environmental values in a biogeographic transitional area in north-western Mexico. Contrib. Zool. 2014, 83, 151–166. [Google Scholar]
- Casas-Andreu, G.; Aguilar-Miguel, X.; Cruz-Aviña, R. La introducción y el cultivo de la rana toro (Rana catesbeiana). ¿Un atentado a la biodiversidad de México? Revista Científica Multidisciplinaria de Prospectiva 2001, 8, 62–67. [Google Scholar]
- Cifuentes-Lemus, J.L.; Torres-García, M.P.; Frías-Mondragón, M. El Océano y Sus Recursos XI. Acuicultura, 2nd ed.; Serie La ciencia Para Todos; Fondo de Cultura Económica: México City, Mexico, 1997; pp. 1–97. ISBN 968-16-5242-897. [Google Scholar]
- Luja, V.H.; Rodrìguez-Estrella, R. The invasive bullfrog Lithobates catesbeianus in oases of Baja California Sur Mexico: Potential effect in a fragile ecosystem. Biol. Invasions 2010, 12, 2979–2983. [Google Scholar] [CrossRef]
- Morales-Salinas, E.; Aguilar-Arriaga, B.O.; Ramírez-Lezama, J.; Méndez-Bernal, A.; López-Garrido, S. Oral fibrosarcoma in a black iguana (Ctenosaura pectinata). J. Zoo Wildl. Med. 2013, 44, 513–516. [Google Scholar] [CrossRef]
- Rijks, J.M.; Saucedo, B.; Spitzen-van der Sluijs, A.M.; Wilkie, G.S.; van Asten, A.J.A.M.; van den Broek, J.; Boonyarittichaikij, R.; Stege, M.; Van der Sterren, F.; Martel, A.; et al. Investigation of amphibian mortality events in wildlife reveals an on-going ranavirus epidemic in the North of The Netherlands. PLoS ONE 2016, 11, e0157473. [Google Scholar] [CrossRef]
- Zhang, Y.; Uyemoto, J.K.; Kirkpatrick, B.C. A small-scale procedure for extracting nucleic acids from woody plants infected with various phytopathogens for PCR assay. J. Virol. Methods 1998, 71, 45–50. [Google Scholar] [CrossRef]
- Mao, J.; Hendrick, R.P.; Chinchar, V.G. Molecular characterization, sequence analysis, and taxonomic position of newly isolated fish iridoviruses. Virology 1997, 229, 212–220. [Google Scholar] [CrossRef]
- CLUSTAL Omega. Available online: https://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 30 August 2018).
- D’hondt, B.; Vanderhoeven, S.; Roelandt, S.; Mayer, F.; Versteirt, V.; Ducheyne, E.; San Martin, G.; Grégoire, J.C.; Stiers, I.; Quoilin, S.; et al. Pandora: A Risk Screening Tool for Pathogens and Parasites; Belgian Biodiversity Platform: Brussels, Belgium, 2014; pp. 1–22. [Google Scholar]
- Schiphouwer, M.E.; Felix, R.P.W.H.; van Duinen, G.A.; de Hoop, L.; de Hullu, P.C.; Matthews, J.; van der Velde, G.; Leuven, R.S.E.W. Risk assessment of the alien smallmouth bass (Micropterus dolomieu). Rep. Environ. Sci. 2017, 527, 1–60. [Google Scholar]
- Rijks, J.M.; Spitzen van-der Sluijs, A.; Leuven, R.S.E.W.; Martel, A.; Kik, M.; Pasmans, F. Risk Analysis of the Common Midwife Toad-Like Virus (CMTV-Like Virus) in The Netherlands; Report 60000784-2013; Netherlands Food and Consumer Product Safety Authority (NVWA), Dutch Ministry of Economic Affairs: Utrecht, The Netherlands, 2012. [Google Scholar]
- Wilson, L.D.; Johnson, J.D.; Mata-Silva, V. A conservation reassessment of the amphibians of Mexico based on the EVS measure. Amphib. Reptile Conserv. 2013, 7, 97–127. [Google Scholar]
- Saucedo, B.; Hughes, J.; Súarez, N.; Haenen, O.; Kik, M.J.L.; van Beurden, S.J. Complete genome sequence of frog virus 3 isolated from a Strawberry poison frog (Oophaga pumilio) imported from Nicaragua into The Netherlands. Genome Announc. 2017, 5, ee00863-17. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.G.; Barkman, T.J.; Chinchar, G.V.; Essani, K. Comparative genomic analyses of frog virus 3, type species of the genus Ranavirus (family Iridoviridae). Virology 2004, 323, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Morrison, E.A.; Garner, S.; Echaubard, P.; Lesbarreres, D.; Kyle, C.J.; Brunetti, C.R. Complete genome analysis of a frog virus 3 (FV3) isolate and sequence comparison with isolates of differing levels of virulence. Virol. J. 2014, 12, 11–46. [Google Scholar] [CrossRef]
- Kolby, J.E.; Smith, K.M.; Berger, L.; Karesh, W.B.; Preston, A.; Pessier, A.P.; Skerratt, L.F. First evidence of amphibian chytrid fungus (Batrachochytrium dendrobatidis) and ranavirus in Hong Kong amphibian trade. PLoS ONE 2014, 3, 1–6. [Google Scholar] [CrossRef]
- Mazzoni, R.; Mesquita, A.J.; Fleury, L.F.F.; Brito, W.; Nunes, I.A.; Robert, J.; Morales, H.; Coelho, A.S.G.; Barthasson, D.L.; Galli, L.; et al. Mass mortality associated with a Frog virus 3-like ranavirus infection in farmed tadpoles Rana catesbeiana from Brazil. Dis. Aquat. Org. 2009, 86, 181–191. [Google Scholar] [CrossRef]
- Une, Y.; Sakuma, A.; Matsueda, H.; Nakai, K.; Murakami, M. Ranavirus outbreak in North American bullfrogs (Rana catesbeiana), Japan, 2008. Emerg. Infect. Dis. 2008, 15, 1146–1147. [Google Scholar] [CrossRef]
- Cunningham, A.A.; Temns, C.A.; Russel, P.H. Immunohistochemical demonstration of ranavirus antigen in the tissues of infected frogs (Rana temporaria) with systemic hameorrhagic or cutaneous ulcerative disease. J. Comp. Pathol. 2008, 138, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Willis, Y.L.; Moyle, P.B.; Basket, T.S. Emergence, breeding, hibernation, movements and transformation of the bullfrog, Rana catesbeiana, in Missouri. Copeia 1956, 1, 30–41. [Google Scholar] [CrossRef]
- Aguilar Ibarra, F. Aspectos Generales sobre las Ranas y su Cultivo. Trab. Div. Inst. Nac. Invest. Biol. Pesq. C 1963, 5, 1–21. [Google Scholar]
- Hardy, L.M.; McDiarmid, R.W. The amphibians and reptiles of Sinaloa, Mexico. Univ. Kansas Publ. Mus. Nat. Hist. 1969, 18, 39–252. [Google Scholar] [CrossRef]
- Nazir, J.; Spengler, M.; Marschang, R.E. Environmental persistence of amphibian and reptilian ranaviruses. Dis. Aquat. Org. 2012, 98, 177–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echaubard, P.; Paulli, B.D.; Trudeau, V.L.; Lesbarrères, D. Ranavirus infection in northern leopard frogs: The timing and number of exposures matter. J. Zool. 2015, 298, 30–36. [Google Scholar] [CrossRef]
- Earl, J.E.; Chaney, J.C.; Sutton, W.B.; Lillard, C.E.; Kouba, A.J.; Langhorne, C.; Krebs, J.; Wilkis, R.P.; Hill, R.D.; Miller, D.L.; et al. Ranavirus could facilitate local extinction of rare amphibian species. Oecologia 2016, 182, 611–623. [Google Scholar] [CrossRef]
- Jerett, I.V.; Whittington, R.J.; Weir, R.P. Pathology of a Bohle-like infection in two Australian frog species (Litoria splendida and Litoria caerulea). J. Comp. Pathol. 2015, 152, 248–259. [Google Scholar] [CrossRef]
- Brand, M.D.; Hill, R.D.; Brenes, R.; Chaney, J.C.; Wilkes, R.P.; Grayfer, L.; Miller, D.L.; Gray, M.J. Water temperature affects susceptibility to ranavirus. Ecohealth 2016, 13, 350–359. [Google Scholar] [CrossRef]
- Mao, J.D.E.; Green, G.; Chinchar, V.G. Molecular characterization of iridoviruses isolated from sympatric amphibians and fish. Virus Res. 1999, 63, 45–52. [Google Scholar] [CrossRef]
- Moody, N.; Owens, L. Experimental demonstration of the pathogenicity of frog virus, Bohle iridovirus, for a fish species, barramundi, Lates calcariger. Dis. Aquat. Org. 1994, 18, 95–102. [Google Scholar] [CrossRef]
- Rodríguez-Meza, D.; Rodríguez Figueroa, G.; Sapozhnikov, D.; Vargas-Ramírez, C.; Vallejo-Soto, A.; Verdugo-Quiñonez, G.; Michel-Rubio, A. Monitoreo de la calidad del agua del acuífero de Guasave, Sinaloa Mexico. Unpublished work, SIP 2008.
- Price, S.J.; Leung, W.T.M.; Christopher, J.O.; Sergeant, C.; Cunningham, A.A.; Ballous, F.; Garner, T.W.J.; Nichols, R. A. Temperature is a key driver of a wildlife epidemic and future warming will increase impact. bioRxiv 2018. [Google Scholar] [CrossRef]
- Jancovich, J.K.; Chinchar, V.G.; Hyatt, A.; Miyazaki, T.; Zhang, Q.Y. Virus Taxonomy: Ninth Report of the International Committee of Taxonomy of Viruses; King, A.M.Q., Adams, M.J., Carstens, E.B., Lefkowitz, E., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2012; pp. 193–210. [Google Scholar]
- Saucedo, B.; Hughes, J.; Spitzen van der Sluijs, A.; Kruithof, N.; Schils, M.; Rijks, J.M.; Jacinto-Maldonado, M.; Suárez, N.; Haenen, O.L.M.; Voorbergen-Laarman, M.; et al. Ranavirus genotypes in The Netherlands and their potential association with virulence in water frogs (Pelophylax spp.). Emerg. Microbes Infect. 2018, 7, 1–56. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A.; Langton, T.E.S.; Bennet, P.W.; Lewing, J.F.; Drury, S.E.N.; Gough, R.E.; MacGregor, E.K. Pathological and microbiological findings from incidents of unusual mortality of the common frog (Rana temporaria). Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1539–1557. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Module | Risk Classification | Risk Score | Certainty | Confidence |
|---|---|---|---|---|
| Emerging/entry | Medium | 0.5 | Medium | 0.5 |
| Emerging/exposure | High | 1 | Medium | 0.75 |
| Environmental score | High | 0.875 | High | 1 |
| Plant score | n/a | 0 | High | 1 |
| Animal score | n/a | 0 | High | 1 |
| Human score | n/a | 0 | High | 1 |
| Other score | Low | 0 | Medium | 0.5 |
| Consequence | High | 0.875 | ||
| Entry exposure | High | 0.707 | ||
| Risk score | High | 0.619 |
| Family | Scientific Name | Common Name | Endemic | IUCN Status | EVS Score | Distance to Farm |
|---|---|---|---|---|---|---|
| Ambystomatidae | Ambystoma rosaceum | Tarahumara salamander | Yes | LC | 14 | 100–200 km |
| Bufonidae | Anaxyrus cognatus | Great plains toad | No | LC | 9 | 100–200 km |
| Bufonidae | Anaxyrus kelloggi | Little mexican toad | Yes | LC | 14 | <50 km |
| Bufonidae | Anaxyrus mexicanus | Mexican spadefoot toad | Yes | NT | 13 | 100–200 km |
| Bufonidae | Anaxyrus punctatus | Red-spotted toad | No | LC | 5 | <50 km |
| Bufonidae | Incilius alvarius | Sonoran desert toad | No | LC | 11 | <50 km |
| Bufonidae | Incilius marmoreus | Marbled toad | Yes | LC | 11 | >200 km |
| Bufonidae | Incilius mazatlanensis | Sinaloan toad | Yes | LC | 12 | <50 km |
| Bufonidae | Incilius occidentalis | Pine toad | Yes | LC | 11 | >200 km |
| Bufonidae | Rhinella marina | Cane toad | No | LC | 3 | <50 km |
| Craugastoridae | Craugastor augusti | Barking frog | No | LC | 8 | <50 km |
| Craugastoridae | Craugastor horbartsmithii | Smith’s pigmy robber frog | Yes | E | 15 | >200 km |
| Craugastoridae | Craugastor occidentalis | Taylor’s barking frog | Yes | NL | 13 | >200 km |
| Craugastoridae | Craugastor vocalis | Pacific stream frog | Yes | LC | 13 | <50 km |
| Eleutherodactylae | Eleutherodactylus interorbitalis | Spectacled chirping frog | Yes | NL | 15 | 100–200 km |
| Eleutherodactylae | Eleutherodactylus nitidus | Shiny peeping frog | Yes | LC | 12 | >200 km |
| Eleutherodactylae | Eleutherodactylus saxatilis | Marbled peeping frog | Yes | E | 17 | >200 km |
| Eleutherodactylae | Eleutherodactylus teretistes | Whistling frog | Yes | NL | 16 | >200 km |
| Hylidae | Agalychnis dacnicolor | Mexican leaf frog | Yes | LC | 13 | <50 km |
| Hylidae | Dryophytes arenicolor | Canyon treefrog | No | LC | 7 | >100 km |
| Hylidae | Dryophytes eximius | Mountain treefrog | Yes | LC | 10 | >200 km |
| Hylidae | Dryophytes wrightorum | Wright’s mountain treefrog | No | LC | 9 | >200 km |
| Hylidae | Exerodonta smaradigna | Emerald treefrog | Yes | LC | 12 | >200 km |
| Hylidae | Sarcohyla bistincta | Mexican Fringed-limbed treefrog | Yes | LC | 9 | >200 km |
| Hylidae | Smilisca baudinii | Common mexican treefrog | No | LC | 3 | <50 km |
| Hylidae | Smilisca fodiens | Lowland burrowing treefrog | No | LC | 8 | <50 km |
| Hylidae | Tlalocohyla smithii | Dwarf mexican treefrog | No | LC | 11 | <50 km |
| Hylidae | Trachycephalus typhonius | Veined treefrog | No | LC | 4 | >200 km |
| Hylidae | Triprion spatulatus | Mexican shovel-headed treefrog | Yes | LC | 13 | >200 km |
| Leptodactylae | Leptodactylus melanotonus | Black-backed frog | No | LC | 6 | <50 km |
| Mycrohylidae | Gastrophryne mazatlanensis | Mazatlan narrowmouth frog | No | NL | 8 | <50 km |
| Mycrohylidae | Hypopachus ustus | Two-spaded narrow-mouthed toad | No | LC | 7 | 100–200 km |
| Mycrohylidae | Hypopachus variolosus | Mexican narrow-mouthed toad | No | LC | 4 | 100–200 km |
| Pelobatidae | Scaphiopus couchii | Couch’s spadefoot toad | No | LC | 3 | <50 km |
| Plethodontidae | Isthmura bellii | Bell’s salamander | Yes | V | 12 | 50–100 km |
| Ranidae | Lithobates catesbeianus | American bullfrog | No | LC | 10 | <50 km |
| Ranidae | Lithobates forreri | Forrer’s leopard frog | No | LC | 3 | <50 km |
| Ranidae | Lithobates magnaocularis | Northwest mexican leopard frog | Yes | LC | 12 | <50 km |
| Ranidae | Lithobates pustulosus | Mexican cascades frog | Yes | LC | 9 | >200 km |
| Ranidae | Lithobates tarahumarae | Tarahumara frog | Yes | V | 8 | 50–100 km |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saucedo, B.; Serrano, J.M.; Jacinto-Maldonado, M.; Leuven, R.S.E.W.; Rocha García, A.A.; Méndez Bernal, A.; Gröne, A.; Van Beurden, S.J.; Escobedo-Bonilla, C.M. Pathogen Risk Analysis for Wild Amphibian Populations Following the First Report of a Ranavirus Outbreak in Farmed American Bullfrogs (Lithobates catesbeianus) from Northern Mexico. Viruses 2019, 11, 26. https://doi.org/10.3390/v11010026
Saucedo B, Serrano JM, Jacinto-Maldonado M, Leuven RSEW, Rocha García AA, Méndez Bernal A, Gröne A, Van Beurden SJ, Escobedo-Bonilla CM. Pathogen Risk Analysis for Wild Amphibian Populations Following the First Report of a Ranavirus Outbreak in Farmed American Bullfrogs (Lithobates catesbeianus) from Northern Mexico. Viruses. 2019; 11(1):26. https://doi.org/10.3390/v11010026
Chicago/Turabian StyleSaucedo, Bernardo, José M. Serrano, Mónica Jacinto-Maldonado, Rob S. E. W. Leuven, Abraham A. Rocha García, Adriana Méndez Bernal, Andrea Gröne, Steven J. Van Beurden, and César M. Escobedo-Bonilla. 2019. "Pathogen Risk Analysis for Wild Amphibian Populations Following the First Report of a Ranavirus Outbreak in Farmed American Bullfrogs (Lithobates catesbeianus) from Northern Mexico" Viruses 11, no. 1: 26. https://doi.org/10.3390/v11010026
APA StyleSaucedo, B., Serrano, J. M., Jacinto-Maldonado, M., Leuven, R. S. E. W., Rocha García, A. A., Méndez Bernal, A., Gröne, A., Van Beurden, S. J., & Escobedo-Bonilla, C. M. (2019). Pathogen Risk Analysis for Wild Amphibian Populations Following the First Report of a Ranavirus Outbreak in Farmed American Bullfrogs (Lithobates catesbeianus) from Northern Mexico. Viruses, 11(1), 26. https://doi.org/10.3390/v11010026