The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda


{kind=link}
Abstract
:1. Defectiveness in Replication: The Bryan High-Titer Strain as the Prototype
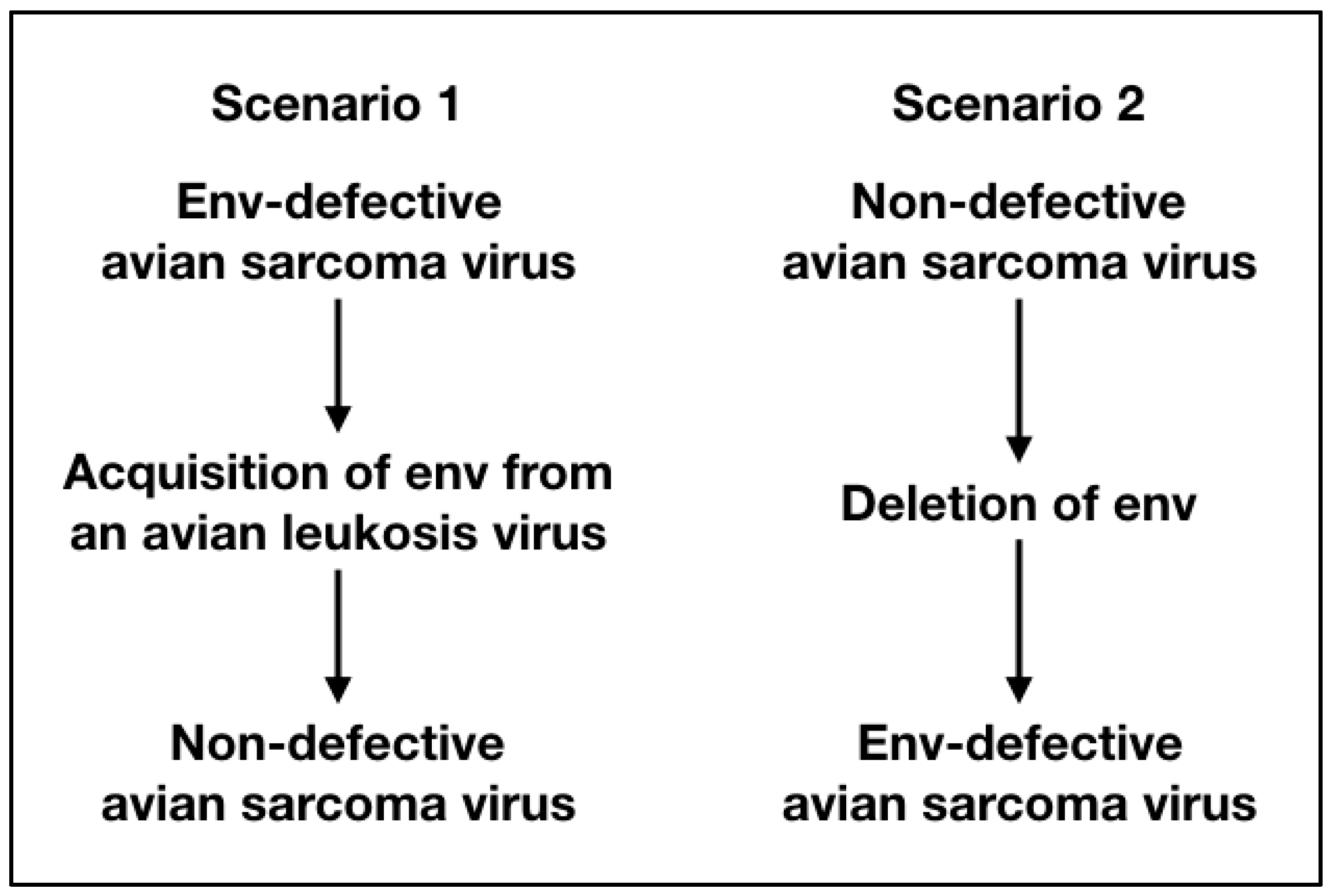
2. Non-Defective Avian Sarcoma Viruses
3. Conditional and Non-Conditional Mutants
4. The Continuing Significance of Oncogenic Retroviruses
5. Idle Speculation
6. Jan Svoboda, the Catalyst
Funding
Conflicts of Interest
References
- Temin, H.M.; Rubin, H. Characteristics of an assay for Rous sarcoma virus and Rous sarcoma cells in tissue culture. Virology 1958, 6, 669–688. [Google Scholar] [CrossRef]
- Hartley, J.W.; Rowe, W.P. Production of altered cell foci in tissue culture by defective Moloney sarcoma virus particles. Proc. Natl. Acad. Sci. USA 1966, 55, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Huebner, R.J.; Hartley, J.W.; Rowe, W.P.; Lane, W.T.; Capps, W.I. Rescue of the defective genome of Moloney sarcoma virus from a noninfectious hamster tumor and the production of pseudotype sarcoma viruses with various murine leukemia viruses. Proc. Natl. Acad. Sci. USA 1966, 56, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Somers, K.D.; Kirsten, W.H. Focus formation by murine sarcoma virus: Enhancement by DEAE-dextran. Virology 1968, 36, 155–157. [Google Scholar] [CrossRef]
- Klement, V.; Rowe, W.P.; Hartley, J.W.; Pugh, W.E. Mixed culture cytopathogenicity: A new test for growth of murine leukemia viruses in tissue culture. Proc. Natl. Acad. Sci. USA 1969, 63, 753–758. [Google Scholar] [CrossRef] [PubMed]
- Klement, V.; Hartley, J.W.; Rowe, W.P.; Huebner, R.J. Recovery of a hamster-specific, focus-forming, and sarcomagenic virus from a “noninfectious” hamster tumor induced by the Kirsten Mouse sarcoma virus. J. Natl. Cancer Inst. 1969, 43, 925–934. [Google Scholar]
- Kirsten, W.H.; Schauf, V.; McCoy, J. Properties of a murine sarcoma virus. Bibl. Haematol. 1970, 36, 246–249. [Google Scholar]
- Rowe, W.P.; Pugh, W.E.; Hartley, J.W. Plaque assay techniques for murine leukemia viruses. Virology 1970, 42, 1136–1139. [Google Scholar] [CrossRef]
- Levy, J.A.; Hartley, J.W.; Rowe, W.P.; Huebner, R.J. Studies of FBJ osteosarcoma virus in tissue culture. I. Biologic characteristics of the “C”-type viruses. J. Natl. Cancer Inst. 1973, 51, 525–539. [Google Scholar]
- Sklar, M.D.; White, B.J.; Rowe, W.P. Initiation of oncogenic transformation of mouse lymphocytes in vitro by Abelson leukemia virus. Proc. Natl. Acad. Sci. USA 1974, 71, 4077–4081. [Google Scholar] [CrossRef]
- Levy, J.A.; Hartley, J.W.; Rowe, W.P.; Huebner, R.J. Studies of FBJ osteosarcoma virus in tissue culture. II. Autoinhibition of focus formation. J. Natl. Cancer Inst. 1975, 54, 615–619. [Google Scholar] [PubMed]
- Poiesz, B.J.; Ruscetti, F.W.; Reitz, M.S.; Kalyanaraman, V.S.; Gallo, R.C. Isolation of a new type C retrovirus (HTLV) in primary uncultured cells of a patient with Sezary T-cell leukaemia. Nature 1981, 294, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.K. Retroviral oncogenes: A historical primer. Nat. Rev. Cancer 2012, 12, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Bryan, W.R.; Moloney, J.B.; Calnan, D. Stable standard preparations of the Rous sarcoma virus preserved by freezing and storage at low temperatures. J. Natl. Cancer Inst. 1954, 15, 315–329. [Google Scholar] [PubMed]
- Bryan, W.R. Enhancement of virus yield of Rous sarcomas. Acta Unio Int. Contra Cancrum 1959, 15, 764–767. [Google Scholar] [PubMed]
- Rubin, H.; Temin, H.M. A radiological study of cell-virus interaction in the Rous sarcoma. Virology 1959, 7, 75–91. [Google Scholar] [CrossRef]
- Temin, H.M. Further evidence for a converted, non-virus-producing state of Rous sarcoma virus-infected cells. Virology 1963, 20, 235–245. [Google Scholar] [CrossRef]
- Hanafusa, H.; Hanafusa, T.; Rubin, H. The defectiveness of Rous sarcoma virus. Proc. Natl. Acad. Sci. USA 1963, 49, 572–580. [Google Scholar] [CrossRef]
- Dougherty, R.M.; Di Stefano, H.S. Virus particles associated with “nonproducer” Rous sarcoma cells. Virology 1965, 27, 351–359. [Google Scholar] [CrossRef]
- Weiss, R. Spontaneous virus production from “non-virus producing” Rous sarcoma cells. Virology 1967, 32, 719–723. [Google Scholar] [CrossRef]
- Klucking, S.; Adkins, H.B.; Young, J.A. Resistance to infection by subgroups B, D, and E avian sarcoma and leukosis viruses is explained by a premature stop codon within a resistance allele of the tvb receptor gene. J. Virol. 2002, 76, 7918–7921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Bacon, L.D.; Cheng, H.H.; Hunt, H.D. Development and validation of a PCR-RFLP assay to evaluate TVB haplotypes coding receptors for subgroup B and subgroup E avian leukosis viruses in White Leghorns. Avian Pathol. 2005, 34, 324–331. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.M.; Bacon, L.D.; Heidari, M.; Muir, W.M.; Groenen, M.A.; Zhang, Y.; Wong, G.K.; Fulton, J.E.; O’Sullivan, N.P.; Albers, G.A.; et al. Genetic variation at the tumour virus B locus in commercial and laboratory chicken populations assessed by a medium-throughput or a high-throughput assay. Avian Pathol. 2007, 36, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, P.K. A virus released by “nonproducing” Rous sarcoma cells. Proc. Natl. Acad. Sci. USA 1967, 58, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Scheele, C.M.; Hanafusa, H. Proteins of helper-dependent RSV. Virology 1971, 45, 401–410. [Google Scholar] [CrossRef]
- Rychlikova, M.; Svoboda, J. Effects of anti-Rous sarcoma virus sera on a hemorrhagic disease of rats. Virology 1960, 10, 545–547. [Google Scholar] [CrossRef]
- Schmidt-Ruppin, K.H. Heterotransplantation of Rous Sarcoma and Rous Sarcoma Virus to Mammals. Oncologia 1964, 17, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Zilber, L.A. Some data on the interaction of Rous sarcoma virus with mammalian cells. In Proceedings of the International Conference on Avian Tumor Viruses, Duke University, Durham, NC, USA, 31 March–3 April 1964; pp. 261–275. [Google Scholar]
- Thurzo, V.; Slabeyciusova, M.; Klimek, M.; Kovarova, V. New filtrable fowl tumor. Cesk. Onkol. 1954, 1, 230–234. [Google Scholar]
- Svoboda, J. Presence of chicken tumour virus in the sarcoma of the adult rat inoculated after birth with Rous sarcoma tissue. Nature 1960, 186, 980–981. [Google Scholar] [CrossRef]
- Simkovic, D.; Smida, J.; Thurzo, V. Release of tumour virus by cells of the B77 fowl virus tumour in vitro. Neoplasma 1961, 8, 495–500. [Google Scholar]
- Thurzo, V.; Smida, J.; Smidova-Kovarova, V.; Simkovic, D. Some properties of the fowl virus tumour B77. Acta Unio Int. Contra Cancrum 1963, 19, 304–305. [Google Scholar] [PubMed]
- Thurzo, V.; Simkovicova, M.; Lizonova, A. Group-Specific Antigens of Ascites Rat Tumors Induced by B77 Virus. Neoplasma 1971, 18, 551–554. [Google Scholar] [PubMed]
- Svec, J.; Hilubinova, K.; Lizorova, V.; Thurzo, V. Mammalian tropism of B77(RBI) virus. Expression of virus genome in the hamster sarcoma cell clones. Neoplasma 1975, 22, 133–145. [Google Scholar] [PubMed]
- Svoboda, J.; Chyle, P.; Simkovic, D.; Hilgert, I. Demonstration of the absence of infectious Rous virus in rat tumour XC, whose structurally intact cells produce Rous sarcoma when transferred to chicks. Folia Biol. 1963, 9, 77–81. [Google Scholar]
- Svoboda, J.; Hejnar, J.; Geryk, J.; Elleder, D.; Vernerova, Z. Retroviruses in foreign species and the problem of provirus silencing. Gene 2000, 261, 181–188. [Google Scholar] [CrossRef]
- Lounkova, A.; Draberova, E.; Senigl, F.; Trejbalova, K.; Geryk, J.; Hejnar, J.; Svoboda, J. Molecular events accompanying rous sarcoma virus rescue from rodent cells and the role of viral gene complementation. J. Virol. 2014, 88, 3505–3515. [Google Scholar] [CrossRef] [PubMed]
- Golde, A.; Vigier, P. Non-defectivity of the Rous virus of the Prague strain. C. R. Acad. Sci. Hebd. Seances Acad. Sci. D 1966, 262, 2793–2796. [Google Scholar] [PubMed]
- Lai, M.M.; Duesberg, P.H.; Horst, J.; Vogt, P.K. Avian tumor virus RNA: A comparison of three sarcoma viruses and their transformation-defective derivatives by oligonucleotide fingerprinting and DNA-RNA hybridization. Proc. Natl. Acad. Sci. USA 1973, 70, 2266–2270. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.M.; Hu, S.S.; Vogt, P.K. Occurrence of partial deletion and substitution of the src gene in the RNA genome of avian sarcoma virus. Proc. Natl. Acad. Sci. USA 1977, 74, 4781–4785. [Google Scholar] [CrossRef] [Green Version]
- Toyoshima, K.; Vogt, P.K. Temperature sensitive mutants of an avian sarcoma virus. Virology 1969, 39, 930–931. [Google Scholar] [CrossRef]
- Friis, R.R.; Toyoshima, K.; Vogt, P.K. Conditional lethal mutants of avian sarcoma viruses. I. Physiology of ts 75 and ts 149. Virology 1971, 43, 375–389. [Google Scholar] [CrossRef]
- Martin, G.S. Rous sarcoma virus: A function required for the maintenance of the transformed state. Nature 1970, 227, 1021–1023. [Google Scholar] [CrossRef] [PubMed]
- Vogt, P.K. Spontaneous segregation of nontransforming viruses from cloned sarcoma viruses. Virology 1971, 46, 939–946. [Google Scholar] [CrossRef]
- Duesberg, P.H.; Vogt, P.K. Differences between the ribonucleic acids of transforming and nontransforming avian tumor viruses. Proc. Natl. Acad. Sci. USA 1970, 67, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, A.; MacCormick, R.; Martin, G.S. Transformation-defective mutants of avian sarcoma viruses: The genetic relationship between conditional and nonconditional mutants. Virology 1976, 70, 206–209. [Google Scholar] [CrossRef]
- Stehelin, D.; Varmus, H.E.; Bishop, J.M.; Vogt, P.K. DNA related to the transforming gene(s) of avian sarcoma viruses is present in normal avian DNA. Nature 1976, 260, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Vennstrom, B.; Sheiness, D.; Zabielski, J.; Bishop, J.M. Isolation and characterization of c-myc, a cellular homolog of the oncogene (v-myc) of avian myelocytomatosis virus strain 29. J. Virol. 1982, 42, 773–779. [Google Scholar]
- Coffin, J.M.; Hughes, S.H.; Varmus, H.E. (Eds.) Retroviruses; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Robert C. Bast, C.M.C.; Hait, W.N.; Hong, W.K.; Kufe, D.W.; Piccart-Gebhart, M.; Pollock, R.E.; Weichselbaum, R.R.; Wang, H.; Holland, J.F. (Eds.) Holland-Frei Cancer Medicine, 9th ed.; Wiley Blackwell: Hoboken, NJ, USA, 2017. [Google Scholar]
- John Mendelsohn, J.W.G.; Howley, P.M.; Israel, M.A.; Thompson, C.B. (Eds.) The Molecular Basis of Cancer, 4th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2014. [Google Scholar]
- Hanafusa, H.; Baltimore, D.; Smoler, D.; Watson, K.F.; Yaniv, A.; Spiegelman, S. Absence of polymerase protein in virions of alpha-type rous sarcoma virus. Science 1972, 177, 1188–1191. [Google Scholar] [CrossRef]
- Baltimore, D.; Verma, I.M.; Drost, S.; Mason, W.S. Temperature-sensitive DNA polymerase from Rous sarcoma virus mutants. Cancer 1974, 34, 1395–1397. [Google Scholar] [CrossRef]
- Friis, R.R.; Mason, W.S.; Chen, Y.C.; Halpern, M.S. A replication defective mutant of Rous sarcoma virus which fails to make a functional reverse transcriptase. Virology 1975, 64, 49–62. [Google Scholar] [CrossRef]
- Verma, I.M. Studies on reverse transcriptase of RNA tumor viruses. I. Localization of thermolabile DNA polymerase and RNase H activities on one polypeptide. J. Virol. 1975, 15, 121–126. [Google Scholar] [PubMed]
- Mason, W.S.; Yeater, C.; Bosch, J.V.; Wyke, J.A.; Friis, R.R. Fourteen temperature-sensitive replication mutants of Rous sarcoma virus. Virology 1979, 99, 226–240. [Google Scholar] [CrossRef]
- Duesberg, P.H.; Wang, L.-H.; Mellon, P.; Mason, W.S.; Vogt, P.K. Towards a complete genetic map of Rous sarcoma virus. In Animal Virology—Proceedings of the Fifth ICN-UCLA Symposium on Molecular Biology, Squaw Valley, 1976; Baltimore, D., Huang, A.S., Fox, C.F., Eds.; Academic Press: New York, NY, USA, 1976; pp. 107–125. [Google Scholar]
- Duesberg, P.H.; Wang, L.H.; Beemon, K.; Kawai, S.; Hanafusa, H. Sequences and functions of Rous sarcoma virus RNA. Hamatol. Bluttransfus 1976, 19, 327–340. [Google Scholar] [PubMed]
- Hayward, W.S.; Neel, B.G.; Astrin, S.M. Activation of a cellular onc gene by promoter insertion in ALV-induced lymphoid leukosis. Nature 1981, 290, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Roelink, H.; Wagenaar, E.; Lopes da Silva, S.; Nusse, R. Wnt-3, a gene activated by proviral insertion in mouse mammary tumors, is homologous to int-1/Wnt-1 and is normally expressed in mouse embryos and adult brain. Proc. Natl. Acad. Sci. USA 1990, 87, 4519–4523. [Google Scholar] [CrossRef] [PubMed]
- Mladenov, Z.; Heine, U.; Beard, D.; Beard, J.W. Strain MC29 avian leukosis virus. Myelocytoma, endothelioma, and renal growths: Pathomorphological and ultrastructural aspects. J. Natl. Cancer Inst. 1967, 38, 251–285. [Google Scholar] [PubMed]
- Duesberg, P.H.; Bister, K.; Vogt, P.K. The RNA of avian acute leukemia virus MC29. Proc. Natl. Acad. Sci. USA 1977, 74, 4320–4324. [Google Scholar] [CrossRef] [PubMed]
- Maki, Y.; Bos, T.J.; Davis, C.; Starbuck, M.; Vogt, P.K. Avian sarcoma virus 17 carries the jun oncogene. Proc. Natl. Acad. Sci. USA 1987, 84, 2848–2852. [Google Scholar] [CrossRef]
- Vogt, P.K. Fortuitous convergences: The beginnings of JUN. Nat. Rev. Cancer 2002, 2, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.W.; Aoki, M.; Fruman, D.; Auger, K.R.; Bellacosa, A.; Tsichlis, P.N.; Cantley, L.C.; Roberts, T.M.; Vogt, P.K. Transformation of chicken cells by the gene encoding the catalytic subunit of PI 3-kinase. Science 1997, 276, 1848–1850. [Google Scholar] [CrossRef] [PubMed]
- Samuels, Y.; Wang, Z.; Bardelli, A.; Silliman, N.; Ptak, J.; Szabo, S.; Yan, H.; Gazdar, A.; Powell, S.M.; Riggins, G.J.; et al. High frequency of mutations of the PIK3CA gene in human cancers. Science 2004, 304, 554. [Google Scholar] [CrossRef] [PubMed]
- Payne, L.N.; Brown, S.R.; Bumstead, N.; Howes, K.; Frazier, J.A.; Thouless, M.E. A novel subgroup of exogenous avian leukosis virus in chickens. J. Gen. Virol. 1991, 72 Pt 4, 801–807. [Google Scholar] [CrossRef] [Green Version]
- Payne, L.N.; Howes, K.; Gillespie, A.M.; Smith, L.M. Host range of Rous sarcoma virus pseudotype RSV(HPRS-103) in 12 avian species: Support for a new avian retrovirus envelope subgroup, designated J. J. Gen. Virol. 1992, 73 Pt 11, 2995–2997. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, K. Avian leukosis virus subgroup J: A rapidly evolving group of oncogenic retroviruses. Res. Vet. Sci. 1999, 67, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Dong, W.; Yu, C.; He, Z.; Lv, Y.; Sun, Y.; Feng, X.; Li, N.; Lee, L.F.; Li, M. Occurrence of avian leukosis virus subgroup J in commercial layer flocks in China. Avian Pathol. 2004, 33, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Li, Q.; Zhang, Y.; Cui, Z.; Chang, S.; Zhao, P. Isolation and characterization of subgroup J Avian Leukosis virus associated with hemangioma in commercial Hy-Line chickens. Poult. Sci. 2018, 97, 2667–2674. [Google Scholar] [CrossRef] [PubMed]
- Bagust, T.J.; Fenton, S.P.; Reddy, M.R. Detection of subgroup J avian leukosis virus infection in Australian meat-type chickens. Aust. Vet. J. 2004, 82, 701–706. [Google Scholar] [CrossRef]
- Malkinson, M.; Banet-Noach, C.; Davidson, I.; Fadly, A.M.; Witter, R.L. Comparison of serological and virological findings from subgroup J avian leukosis virus-infected neoplastic and non-neoplastic flocks in Israel. Avian Pathol. 2004, 33, 281–287. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Tsukamoto, K.; Maeda, M. Multicentric histiocytosis related to avian leukosis virus subgroup J (ALV-J)-infection in meat-type local chickens. J. Vet. Med. Sci. 2014, 76, 89–92. [Google Scholar] [CrossRef]
- Zhao, Z.; Rao, M.; Liao, M.; Cao, W. Phylogenetic Analysis and Pathogenicity Assessment of the Emerging Recombinant Subgroup K of Avian Leukosis Virus in South China. Viruses 2018, 10, 194. [Google Scholar] [CrossRef]
- Sorge, J.; Hughes, S.H. Splicing of intervening sequences introduced into an infectious retroviral vector. J. Mol. Appl. Genet. 1982, 1, 547–559. [Google Scholar] [PubMed]
- Sorge, J.; Ricci, W.; Hughes, S.H. cis-Acting RNA packaging locus in the 115-nucleotide direct repeat of Rous sarcoma virus. J. Virol. 1983, 48, 667–675. [Google Scholar] [PubMed]
- Hughes, S.H.; Greenhouse, J.J.; Petropoulos, C.J.; Sutrave, P. Adaptor plasmids simplify the insertion of foreign DNA into helper-independent retroviral vectors. J. Virol. 1987, 61, 3004–3012. [Google Scholar] [PubMed]
- Hughes, S.H. The RCAS vector system. Folia Biol. 2004, 50, 107–119. [Google Scholar]
- Hughes, S. The RCAS System. Available online: Home.ncifcrf.gov/hivdrp/rcas/index.html (accessed on 12 December 2018).
- Rubin, H. The early history of tumor virology: Rous, RIF, and RAV. Proc. Natl. Acad. Sci. USA 2011, 108, 14389–14396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, S.; Hanafusa, H. Isolation of defective mutant of avian sarcoma virus. Proc. Natl. Acad. Sci. USA 1973, 70, 3493–3497. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Wang, L.H.; Hanafusa, T.; Hanafusa, H. Partial nucleotide sequence of Rous sarcoma virus-29 provides evidence that the original Rous sarcoma virus was replication defective. J. Virol. 1985, 55, 728–735. [Google Scholar]
- Wang, L.H.; Beckson, M.; Anderson, S.M.; Hanafusa, H. Identification of the viral sequence required for the generation of recovered avian sarcoma viruses and characterization of a series of replication-defective recovered avian sarcoma viruses. J. Virol. 1984, 49, 881–891. [Google Scholar]
- Svoboda, J.; Geryk, J.; Karakoz, I.; Rejthar, A. Isolation of two transforming viruses from sarcomas obtained in chickens inoculated intraembryonally with a transformation defective mutant of Prague strain Rous sarcoma virus. Folia Biol. 1985, 31, 135–151. [Google Scholar]
- Svoboda, J.; Kandala, J.C.; Geryk, J.; Pichrtova, J.; Guntaka, R.V. A transformation-competent recombinant between v-src and Rous-associated virus RAV-1. J. Virol. 1990, 64, 1873–1877. [Google Scholar]
- Svoboda, J. Conditions for tumorigenic action of Rous virus in heterologous relationships. Acta Unio Int. Contra Cancrum 1962, 18, 154–158. [Google Scholar] [PubMed]
- Svoboda, J.; Simkovic, D. The relationships between virus and cell in rat sarcomas induced by Rous virus. Acta Unio Int. Contra Cancrum 1963, 19, 302–303. [Google Scholar] [PubMed]
- Svoboda, J.; Vesely, P.; Vrba, M.; Jiranek, L. Biological properties of rat cells transformed in vitro by Rous sarcoma virus. Folia Biol. 1965, 11, 251–257. [Google Scholar]
- Svoboda, J.; Hlozanek, I.; Mach, O. Detection of chicken sarcoma virus after transfection of chicken fibroblasts with DNA isolated from mammalian cells transformed with Rous Virus. Folia Biol. 1972, 18, 149–153. [Google Scholar]
- Svoboda, J. Rous sarcoma virus. Intervirology 1986, 26, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, J. On board a raft or boat in the retrovirus sea. Proc. Natl. Acad. Sci. USA 2016, 113, 3927–3931. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vogt, P.K. The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda. Viruses 2019, 11, 80. https://doi.org/10.3390/v11010080
Vogt PK. The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda. Viruses. 2019; 11(1):80. https://doi.org/10.3390/v11010080
Chicago/Turabian StyleVogt, Peter K. 2019. "The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda" Viruses 11, no. 1: 80. https://doi.org/10.3390/v11010080
APA StyleVogt, P. K. (2019). The Importance of Being Non-Defective: A Mini Review Dedicated to the Memory of Jan Svoboda. Viruses, 11(1), 80. https://doi.org/10.3390/v11010080