Detection and Characterization of Cucumis melo Cryptic Virus, Cucumis melo Amalgavirus 1, and Melon Necrotic Spot Virus in Cucumis melo



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and RNA Sequencing
2.2. RACE Analysis and Validation of the Full Genomes
2.3. Sequence and Bioinformatics Analyses
3. Results
3.1. Identification of Three RNA Viruses in Melon Using Next-Generation Sequencing
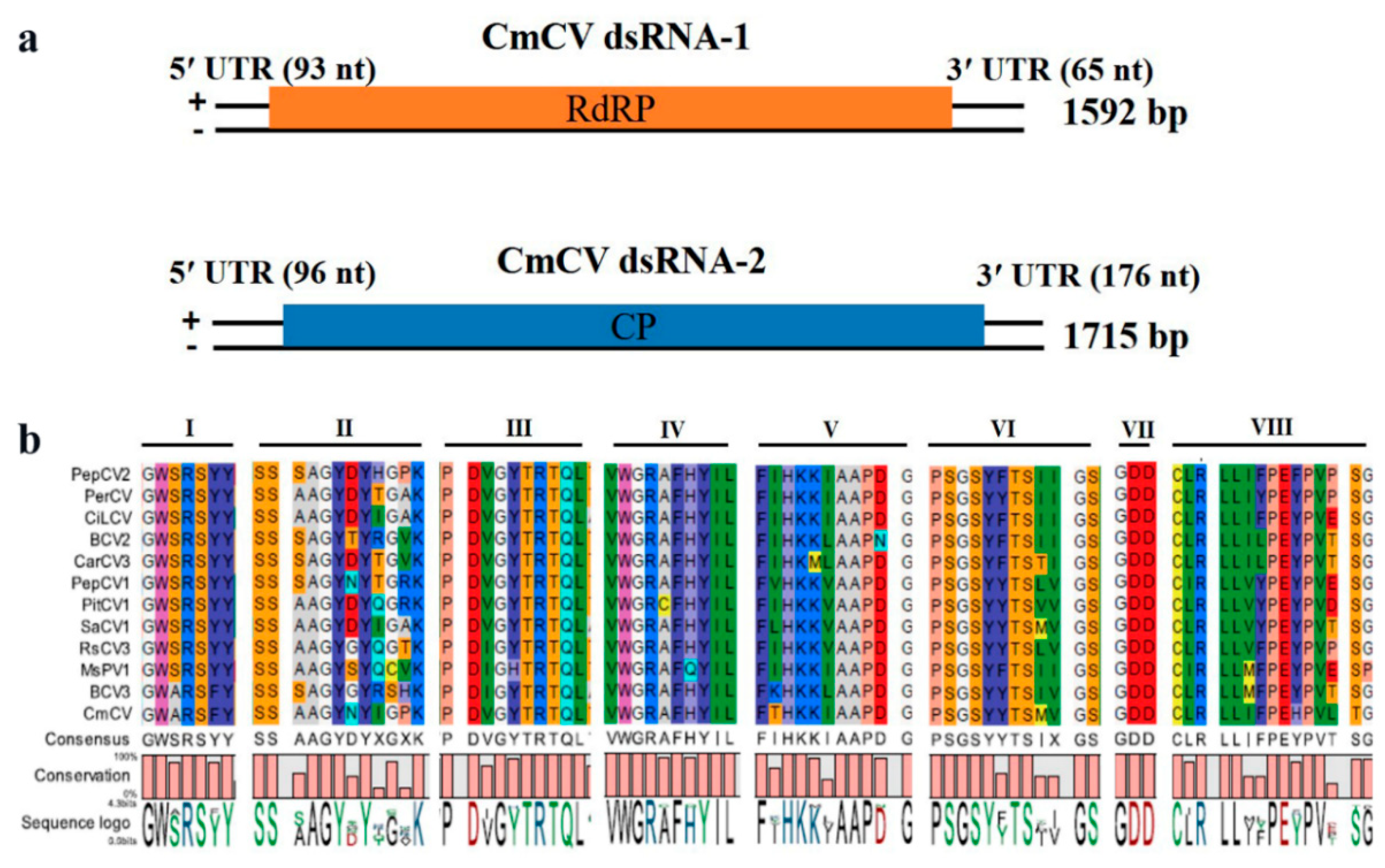
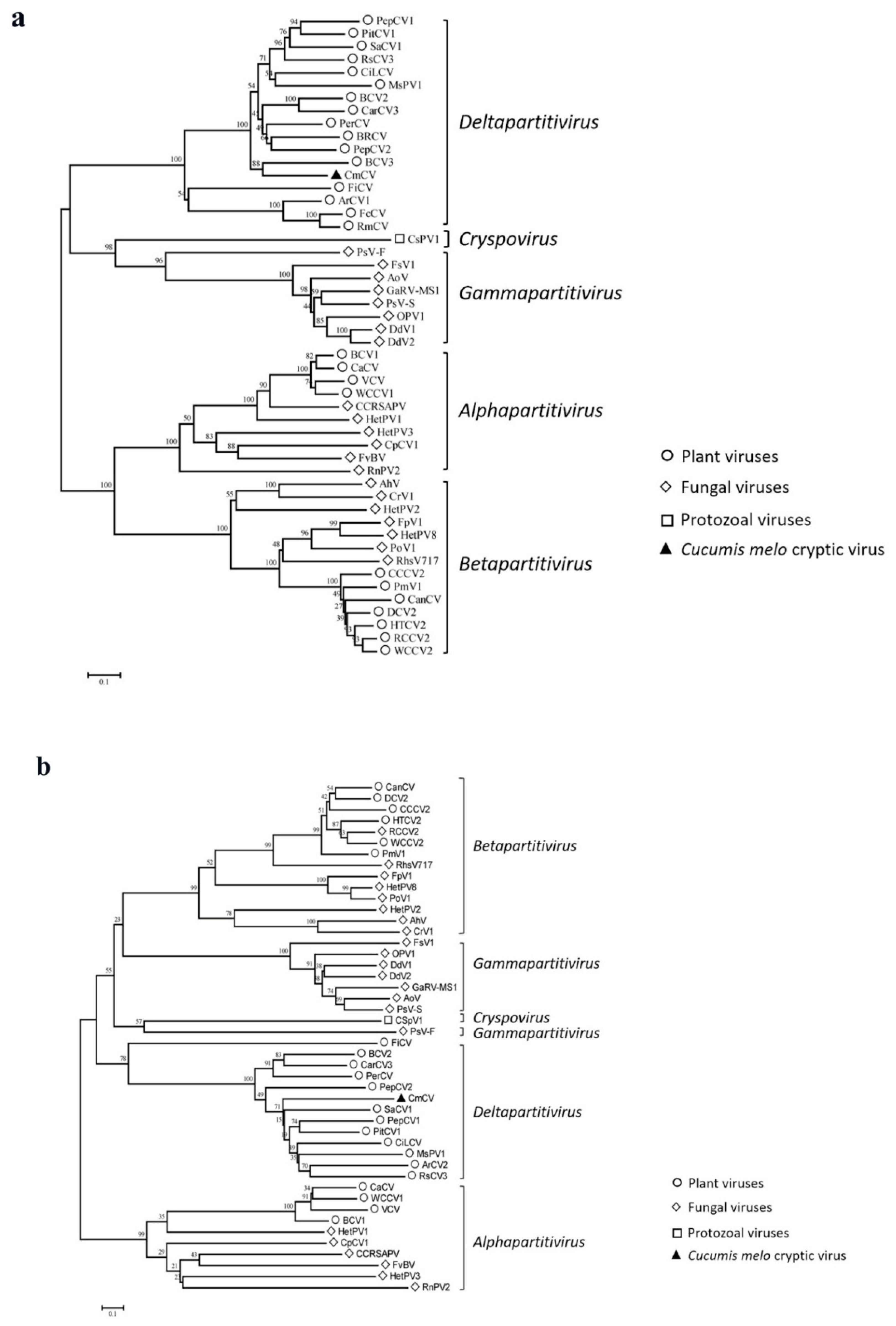
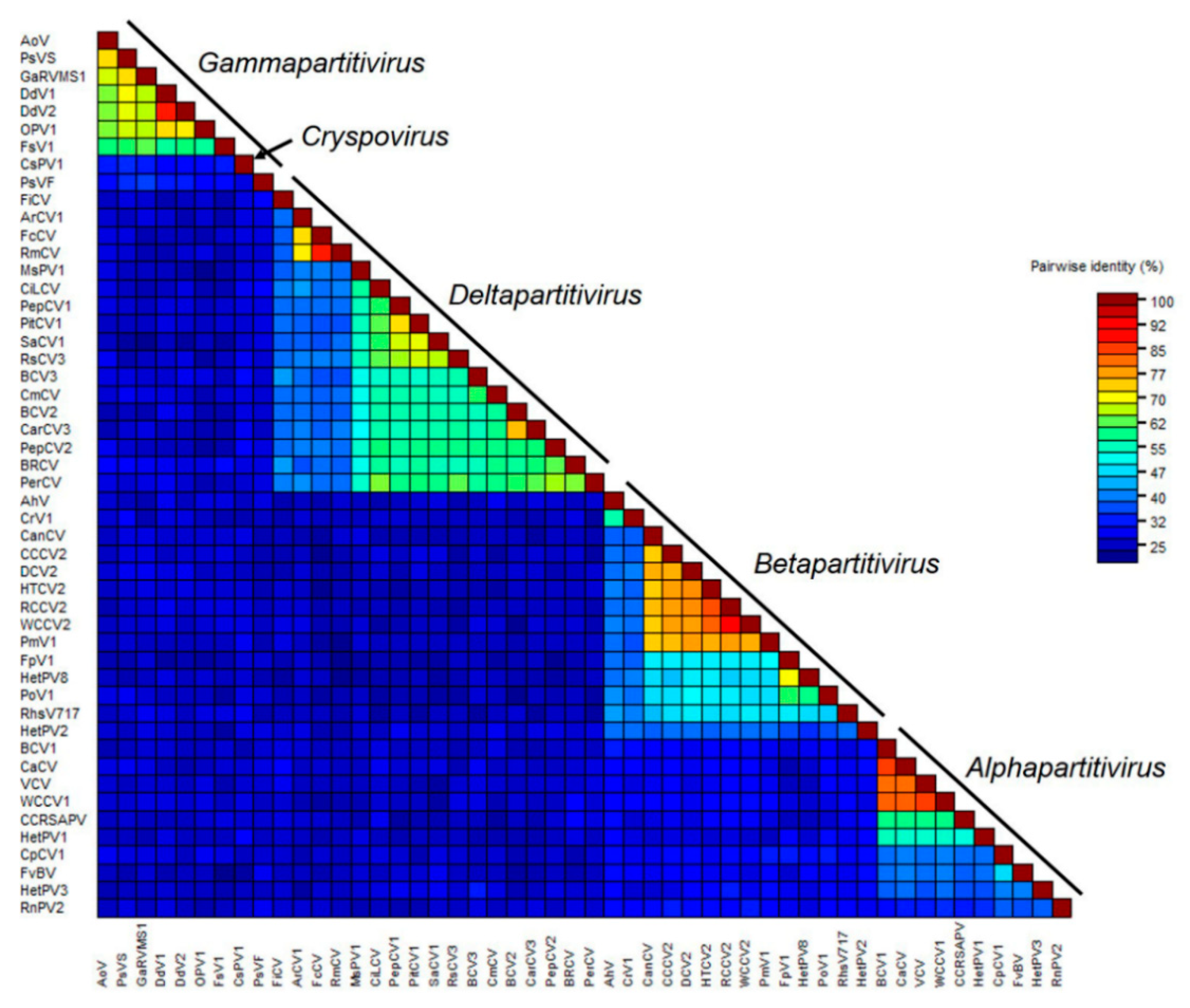
3.2. Characterization of CmCV
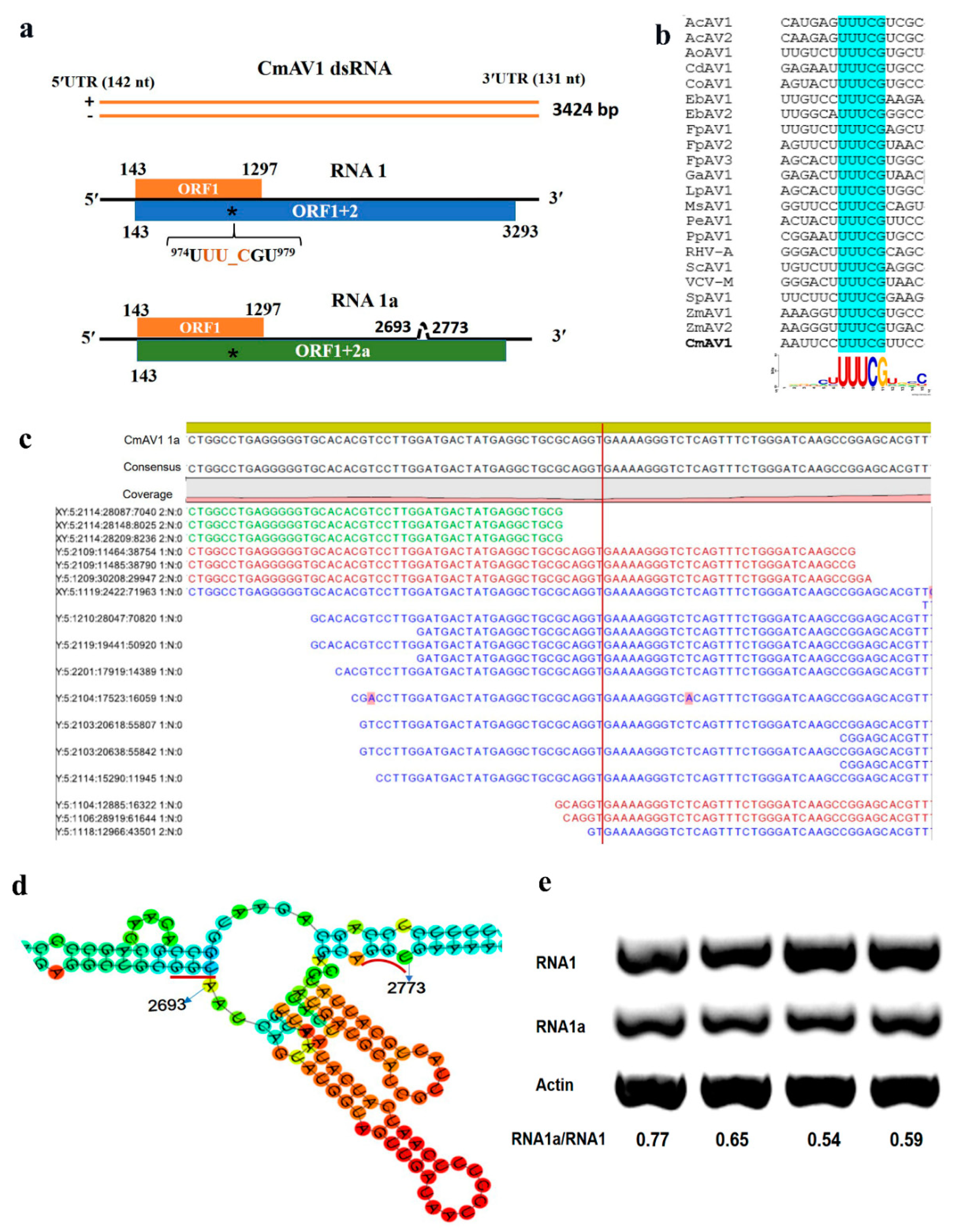
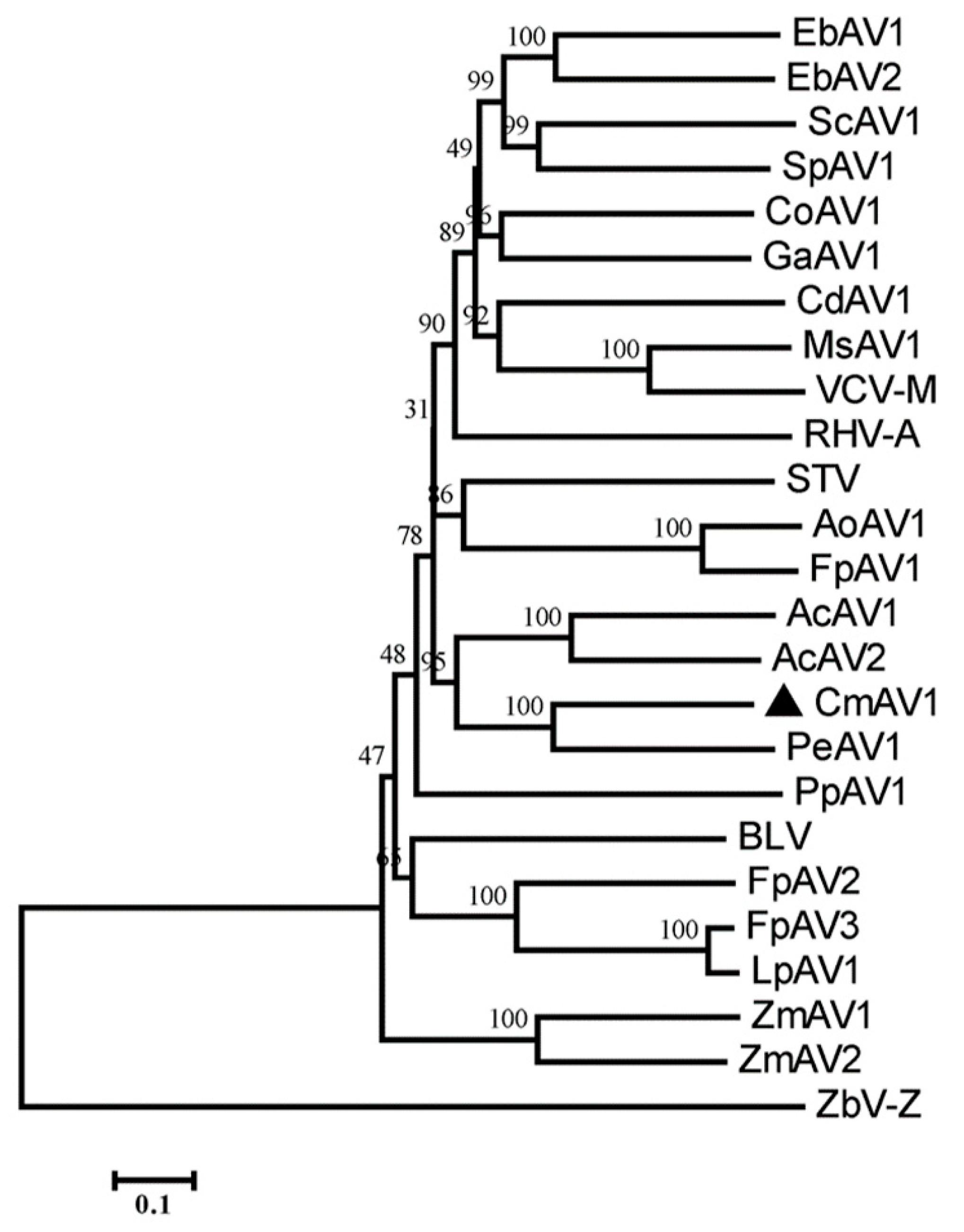
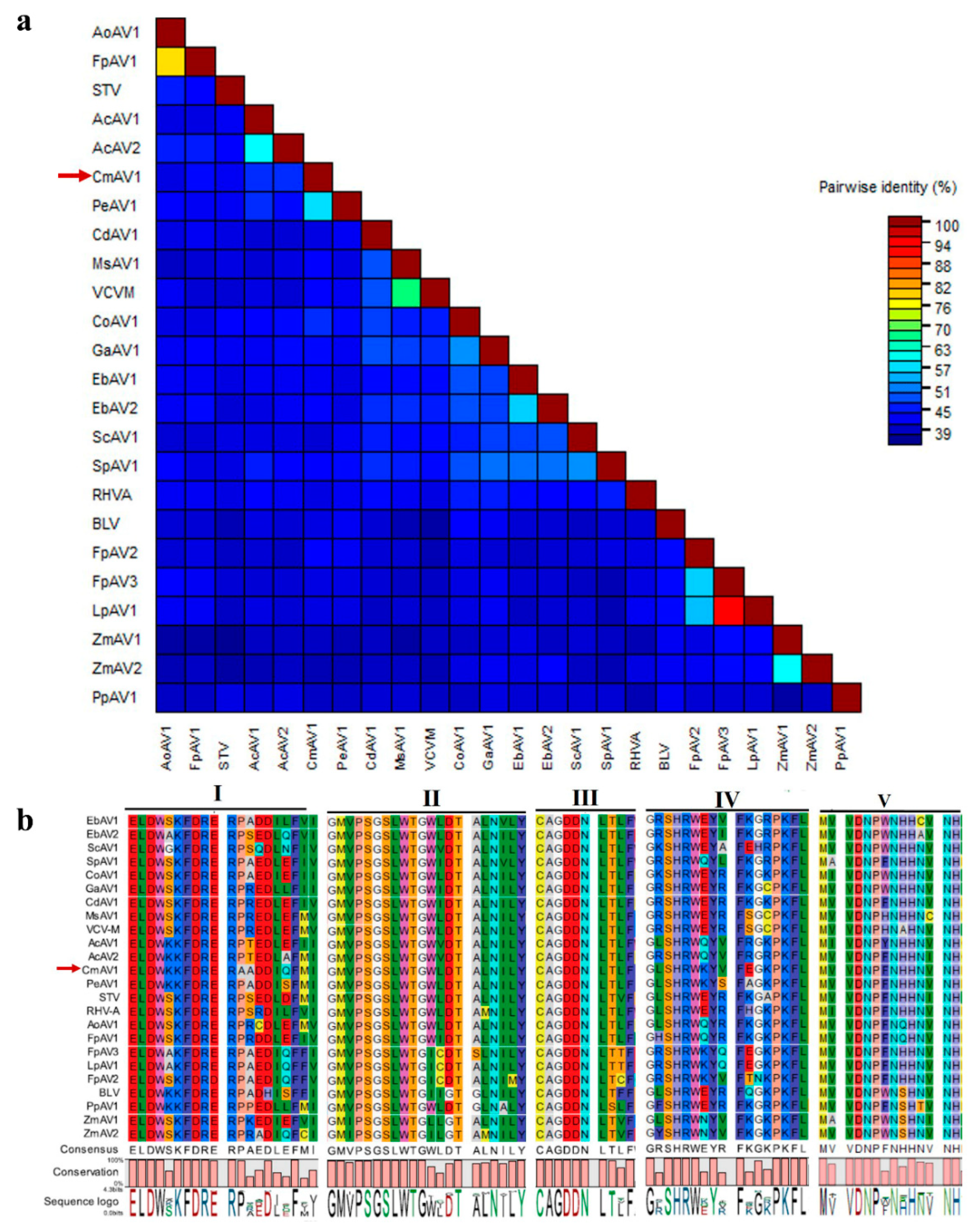
3.3. Characterization of CmAV1
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, M.J.; Antoniw, J.F. DPVweb: A comprehensive database of plant and fungal virus genes and genomes. Nucleic Acids Res. 2006, 34, 382–385. [Google Scholar] [CrossRef] [PubMed]
- Xin, M.; Cao, M.; Liu, W.; Ren, Y.; Lu, C.; Wang, X. The genomic and biological characterization of citrullus lanatus cryptic virus infecting watermelon in China. Virus Res. 2017, 232, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Nibert, M.L.; Ghabrial, S.A.; Maiss, E.; Lesker, T.; Vainio, E.J.; Jiang, D.; Suzuki, N. Taxonomic reorganization of family Partitiviridae and other recent progress in partitivirus research. Virus Res. 2014, 188, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Vainio, E.J.; Chiba, S.; Ghabrial, S.A.; Maiss, E.; Roossinck, M.; Sabanadzovic, S.; Suzuki, N.; Xie, J.; Nibert, M. ICTV Report Consortium. ICTV virus taxonomy profile: Partitiviridae. J. Gen. Virol. 2017, 99, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Boccardo, G.; Lisa, V.; Luisoni, E.; Milne, R.G. Cryptic plant viruses. Adv. Virus Res. 1987, 32, 171–214. [Google Scholar]
- Xiao, X.; Cheng, J.; Tang, J.; Fu, Y.; Jiang, D.; Baker, T.S.; Ghabrial, S.A.; Xie, J. A novel partitivirus that confers hypovirulence on plant pathogenic fungi. J. Virol. 2014, 88, 10120–10133. [Google Scholar] [CrossRef]
- Xie, J.; Jiang, D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases. Annu. Rev. Phytopathol. 2014, 52, 45–68. [Google Scholar] [CrossRef]
- Zhong, J.; Chen, D.; Lei, X.H.; Zhu, H.J.; Zhu, J.Z.; Gao, B.D. Detection and characterization of a novel gammapartitivirus in the phytopathogenic fungus Colletotrichum acutatum strain HNZJ001. Virus Res. 2014, 190, 104–109. [Google Scholar] [CrossRef]
- Righetti, L.; Paris, R.; Ratti, C.; Calassanzio, M.; Onofri, C.; Calzolari, D.; Menzel, W.; Knierim, D.; Magagnini, G.; Pacifico, D. Not the one, but the only one: About cannabis cryptic virus in plants showing ‘hemp streak’ disease symptoms. Eur. J. Plant Pathol. 2018, 150, 575–588. [Google Scholar] [CrossRef]
- Krupovic, M.; Dolja, V.V.; Koonin, E.V. Plant viruses of the Amalgaviridae family evolved via recombination between viruses with double-stranded and negative-strand rna genomes. Biol. Direct 2015, 10, 12. [Google Scholar] [CrossRef]
- Isogai, M.; Nakamura, T.; Ishii, K.; Watanabe, M.; Yamagishi, N.; Yoshikawa, N. Histochemical detection of blueberry latent virus in highbush blueberry plant. J. Gen. Plant Pathol. 2011, 77, 304–306. [Google Scholar] [CrossRef]
- Nibert, M.L.; Pyle, J.D.; Firth, A.E. A +1 ribosomal frameshifting motif prevalent among plant amalgaviruses. Virology 2016, 498, 201–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.; Goh, C.J.; Kim, H.; Hahn, Y. Identification of two novel amalgaviruses in the common eelgrass (Zostera marina) and in silico analysis of the amalgavirus +1 programmed ribosomal frameshifting sites. Plant Pathol. J. 2018, 34, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Firth, A.E.; Jagger, B.W.; Wise, H.M.; Nelson, C.C.; Parsawar, K.; Wills, N.M.; Napthine, S.; Taubenberger, J.K.; Digard, P.; Atkins, J.F. Ribosomal frameshifting used in influenza A virus expression occurs within the sequence UCC_UUU_CGU and is in the +1 direction. Open Biol. 2012, 2, 120109. [Google Scholar] [CrossRef] [PubMed]
- Depierreux, D.; Vong, M.; Nibert, M.L. Nucleotide sequence of Zygosaccharomyces bailii virus Z: Evidence for +1 programmed ribosomal frameshifting and for assignment to family Amalgaviridae. Virus Res. 2016, 217, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.R.; Zhou, J.; Tzanetakis, I.E. Blueberry latent virus: An amalgam of the Partitiviridae and Totiviridae. Virus Res. 2011, 155, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Martin, R. A virus associated with blueberry fruit drop disease. Acta Hortic. 2006, 715, 497–501. [Google Scholar] [CrossRef]
- Sabanadzovic, S.; Valverde, R.J.; Martin, R.; Tzanetakis, I. Southern tomato virus: The link between the families Totiviridae and Partitiviridae. Virus Res. 2009, 140, 130–137. [Google Scholar] [CrossRef]
- Campbell, R.N.; Wipfscheibel, C.; Lecoq, H. Vector-assisted seed transmission of melon necrotic spot virus in melon. Phytopathology 1997, 86, 1294–1298. [Google Scholar] [CrossRef]
- Mallor, C.; Luisarteaga, M.; Alvarez, J.M.; Montaner, C.; Floris, E. Resistance to melon necrotic spot virus in Cucumis melo L. ‘Doublon’ artificially inoculated by the fungus vector Olpidium bornovanus. Crop Prot. 2006, 25, 426–431. [Google Scholar] [CrossRef]
- Gu, Q.S.; Bao, W.H.; Tian, Y.P.; Prins, M.; Yang, H.X.; Lu, J.; Liu, L.F.; Peng, B. Melon necrotic spot virus newly reported in China. Plant Pathol. 2010, 57, 765. [Google Scholar] [CrossRef]
- Chung, B.Y.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An overlapping essential gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef] [PubMed]
- Bouton, C.; Geldreich, A.; Ramel, L.; Ryabova, L.A.; Dimitrova, M.; Keller, M. Cauliflower mosaic virus transcriptome reveals a complex alternative splicing pattern. PLoS ONE 2015, 10, e0132665. [Google Scholar] [CrossRef]
- Zhang, C.X.; Cascone, P.J.; Simon, A.E. Recombination between satellite and genomic RNAs of turnip crinkle virus. Virology 1991, 184, 791–794. [Google Scholar] [CrossRef]
- Panavas, T.; Nagy, P.D. Yeast as a model host to study replication and recombination of defective interfering RNA of tomato bushy stunt virus. Virology 2003, 314, 315–325. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Thwaites, R.; Mumford, R.; Jackeviciene, E.; Navalinskiene, M.; Samuitiene, M.; Boonham, N. Next-generation sequencing and metagenomic analysis: A universal diagnostic tool in plant pathology. Mol. Plant Pathol. 2010, 10, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A.; Flores, R.; Candresse, T.; Barba, M. Next-generation sequencing and genome editing in plant virology. Front Microbiol. 2016, 7, 1325. [Google Scholar] [CrossRef] [PubMed]
- Massart, S.; Candresse, T.; Gil, J.; Lacomme, C.; Predjana, L.; Ravnikar, M.; Reynard, J.; Rumbou, A.; Saldarelli, P.; Skoric, D. A framework for the evaluation of biosecurity, commercial, regulatory, and scientific impacts of plant viruses and viroids identified by NGS technologies. Front Microbiol. 2017, 8, 45. [Google Scholar] [CrossRef] [PubMed]
- Svanella-Dumas, L.; Theil, S.; Barret, M.; Candresse, T. Complete genomic sequence of Raphanus sativus cryptic virus 4 (RsCV4), a novel alphapartitivirus from radish. Arch. Virol. 2018, 163, 1097–1100. [Google Scholar] [CrossRef]
- Osaki, H.; Sasaki, A. A novel alphapartitivirus detected in Japanese pear. Virus Genes 2017, 54, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Hacker, C.V.; Brasier, C.M.; Buck, K.W. Determination of the 5′- and 3′-terminal sequences completes the sequences of the two double-stranded RNAs of Penicillium stoloniferum virus S. Virus Genes 2006, 32, 137–138. [Google Scholar] [CrossRef]
- Lesker, T.; Rabenstein, F.; Maiss, E. Molecular characterization of five betacryptoviruses infecting four clover species and dill. Arch. Virol. 2013, 158, 1943–1952. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Tan, G.; Xiong, G.; Li, M.; Fang, S.; Islam, S.U.; Zhang, S.; Li, F. A new putative deltapartitivirus recovered from Dianthus amurensis. Arch. Virol. 2017, 162, 2897–2901. [Google Scholar] [CrossRef]
- Bujarski, J.J. Genetic recombination in plant-infecting messenger-sense RNA viruses: Overview and research perspectives. Front. Plant Sci. 2013, 4, 68. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.P.; Nagy, P.D. Mechanism of RNA recombination in carmo- and tombusviruses: Evidence for template switching by the RNA-dependent RNA polymerase in vitro. J. Virol. 2003, 77, 12033–12047. [Google Scholar] [CrossRef]
- White, K.A.; Morris, T.J. Nonhomologous RNA recombination in tombusviruses: Generation and evolution of defective interfering RNAs by stepwise deletions. J. Virol. 1994, 68, 14–24. [Google Scholar] [PubMed]
- Lewandowski, D.J.; Dawson, W.O. Deletion of internal sequences results in tobacco mosaic virus defective RNAs that accumulate to high levels without interfering with replication of the helper virus. Virology 1998, 251, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.; Liu, R.; Yu, J.; Ran, Z.; Newkirk, S.J.; An, W.; Li, F.; Wang, D. Identification and characterization of viral defective RNA genomes in influenza B virus. J. Gen. Virol. 2018, 99, 475–488. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Full Name | Abbrev. | Segment | Accession | Length | ORF | Position | Length (aa) |
|---|---|---|---|---|---|---|---|
| Cucumis melon cryptic virus | CmCV | RNA1 | MH479772 | 1592 bp | RdRp | 94–1527 | 477 |
| RNA2 | MH479773 | 1715 bp | CP | 97–1539 | 480 | ||
| Cucumis melon amalgavirus 1 | CmAV1 | RNA1 | MH479774 | 3424 bp | ORF1 (putative CP) | 143–1297 | 384 |
| ORF1 + 2 (fusion protein, RdRp) | 143–973, 975–3293 | 1049 | |||||
| RNA1a | ORF1 + 2a | 143–973, 975–2692, | 1022 | ||||
| 2774–3293 | |||||||
| melon necrotic spot virus | MNSV | RNA1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, B.; Cao, M.; Wang, K.; Wang, X.; Zhou, X. Detection and Characterization of Cucumis melo Cryptic Virus, Cucumis melo Amalgavirus 1, and Melon Necrotic Spot Virus in Cucumis melo. Viruses 2019, 11, 81. https://doi.org/10.3390/v11010081
Zhan B, Cao M, Wang K, Wang X, Zhou X. Detection and Characterization of Cucumis melo Cryptic Virus, Cucumis melo Amalgavirus 1, and Melon Necrotic Spot Virus in Cucumis melo. Viruses. 2019; 11(1):81. https://doi.org/10.3390/v11010081
Chicago/Turabian StyleZhan, Binhui, Mengji Cao, Kaina Wang, Xifeng Wang, and Xueping Zhou. 2019. "Detection and Characterization of Cucumis melo Cryptic Virus, Cucumis melo Amalgavirus 1, and Melon Necrotic Spot Virus in Cucumis melo" Viruses 11, no. 1: 81. https://doi.org/10.3390/v11010081
APA StyleZhan, B., Cao, M., Wang, K., Wang, X., & Zhou, X. (2019). Detection and Characterization of Cucumis melo Cryptic Virus, Cucumis melo Amalgavirus 1, and Melon Necrotic Spot Virus in Cucumis melo. Viruses, 11(1), 81. https://doi.org/10.3390/v11010081