Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility

 , ,
, , 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Challenge Viruses
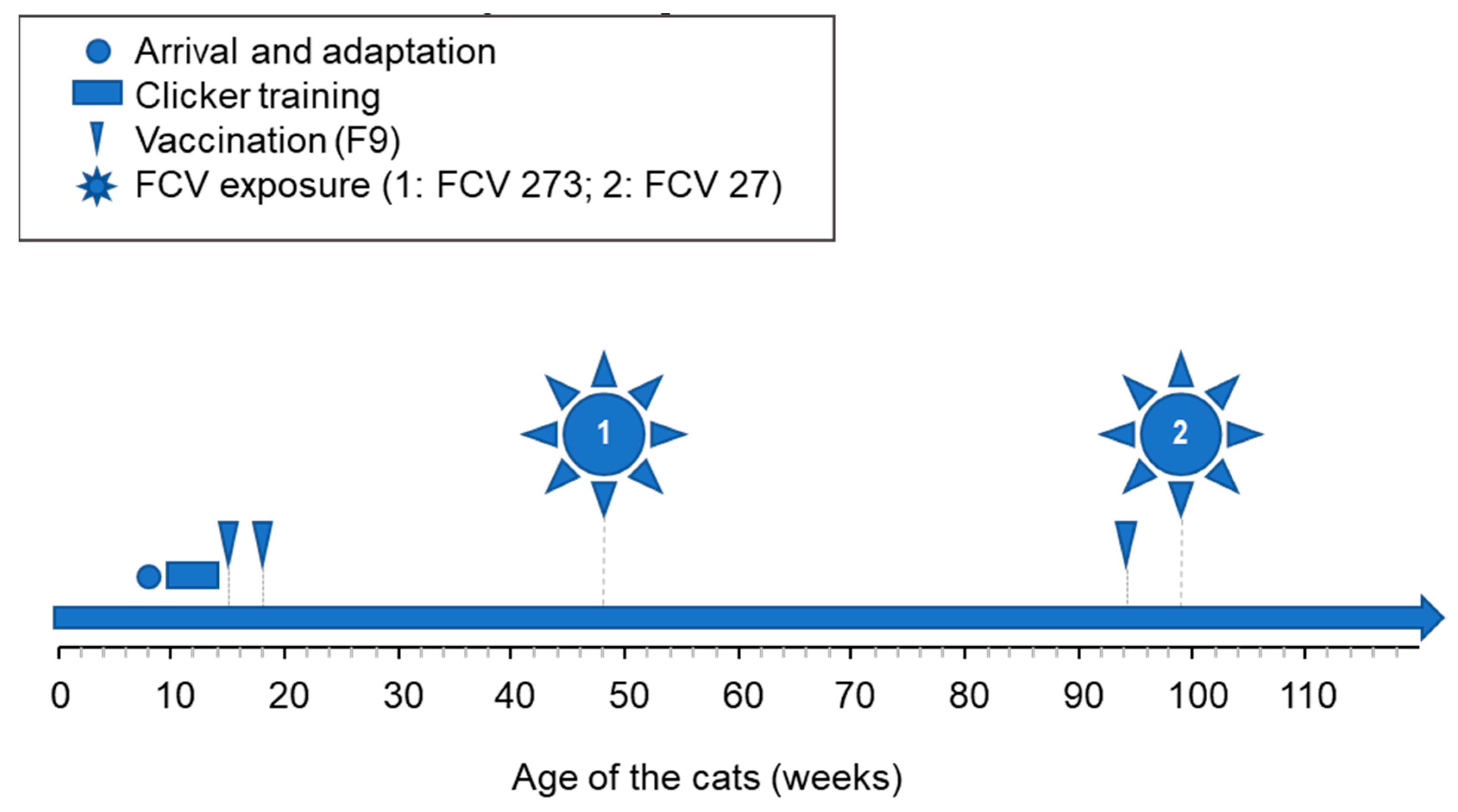
2.3. Vaccination and Experimental Infection of the Cats
2.4. Sample Collection from Cats and Processing
2.5. Environmental Sample Collection and Processing
2.6. TNA Extraction and PCR
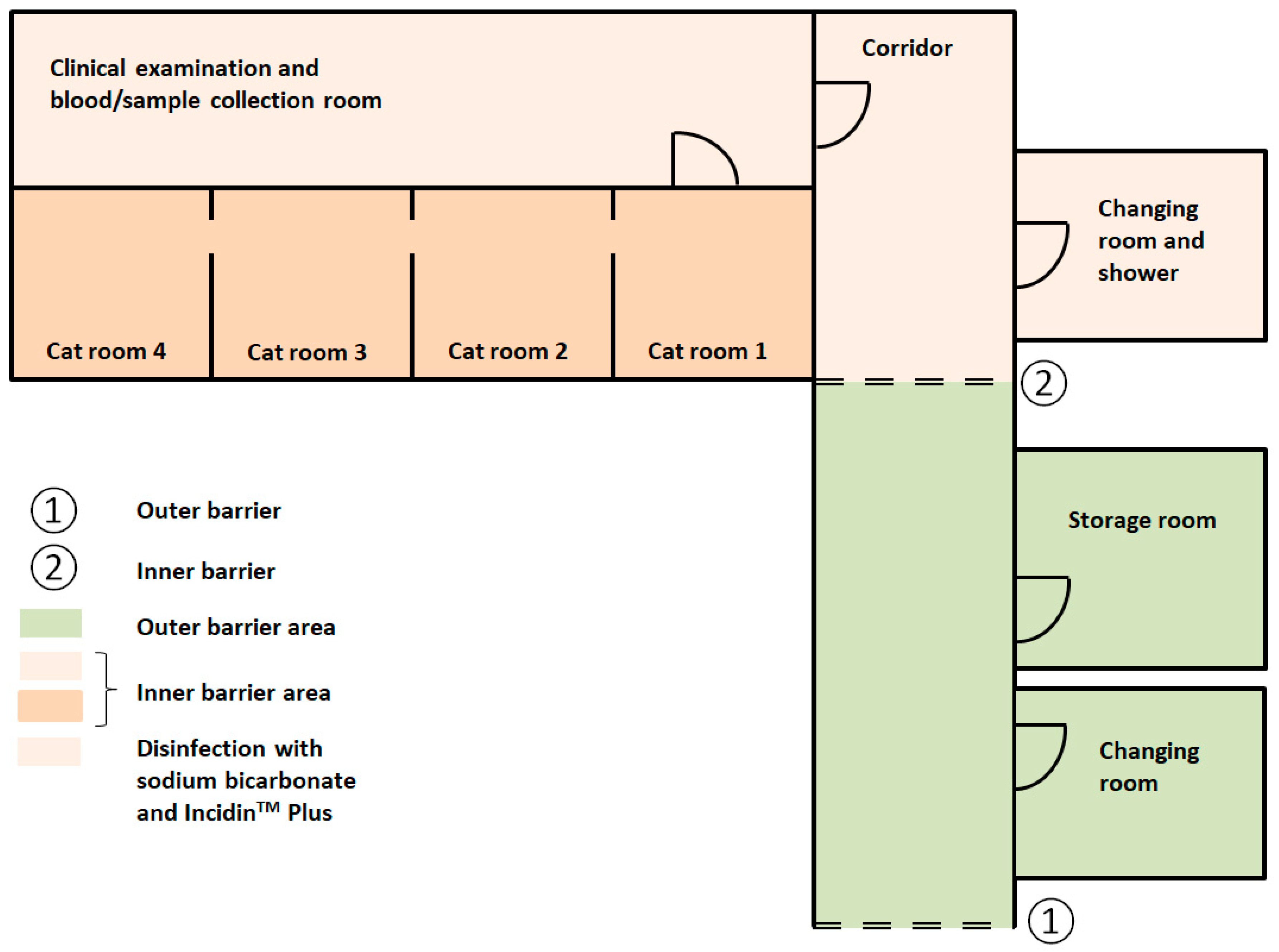
2.7. Hygienic Measures to Enter and Exit the Cat Facility
2.8. Cleaning of the Cat Rooms, the Equipment and the Inner and the Outer Barrier Area
2.9. Statistics
3. Results
3.1. Characterization of the Challenge Viruses FCV 273 and FCV 27
3.2. All Cats Shed FCV after the 1st Experimental Infection with FCV 273
3.3. FCV RNA Is Present on All Items Tested after the 1st Experimental Infection of Cats with FCV
3.4. No Replication Competent FCV Detected on Any of the Tested Items after the 1st FCV Infection
3.5. FCV RNA Detected on the Fur of a Cat after the 1st FCV Infection
3.6. Some Cats were Shedding FCV after the 2nd FCV Challenge
3.7. No FCV was Found in the Environment and the Cat Fur after the 2nd FCV Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Radford, A.D.; Coyne, K.P.; Dawson, S.; Porter, C.J.; Gaskell, R.M. Feline calicivirus. Vet. Res. 2007, 38, 319–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyne, K.P.; Dawson, S.; Radford, A.D.; Cripps, P.J.; Porter, C.J.; McCracken, C.M.; Gaskell, R.M. Long-term analysis of feline calicivirus prevalence and viral shedding patterns in naturally infected colonies of domestic cats. Vet. Microbiol. 2006, 118, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Willi, B.; Meli, M.L.; Boretti, F.S.; Hartnack, S.; Dreyfus, A.; Lutz, H.; Hofmann-Lehmann, R. Feline calicivirus and other respiratory pathogens in cats with Feline calicivirus-related symptoms and in clinically healthy cats in Switzerland. BMC Vet. Res. 2015, 11, 282. [Google Scholar] [CrossRef] [PubMed]
- Radford, A.D.; Addie, D.; Belak, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; et al. Feline calicivirus infection. ABCD guidelines on prevention and management. J. Feline Med. Surg. 2009, 11, 556–564. [Google Scholar] [CrossRef]
- Pedersen, N.C.; Elliott, J.B.; Glasgow, A.; Poland, A.; Keel, K. An isolated epizootic of hemorrhagic-like fever in cats caused by a novel and highly virulent strain of feline calicivirus. Vet. Microbiol. 2000, 73, 281–300. [Google Scholar] [CrossRef]
- Foley, J.; Hurley, K.; Pesavento, P.A.; Poland, A.; Pedersen, N.C. Virulent systemic feline calicivirus infection: Local cytokine modulation and contribution of viral mutants. J. Feline Med. Surg. 2006, 8, 55–61. [Google Scholar] [CrossRef]
- Willi, B.; Spiri, A.M.; Meli, M.L.; Samman, A.; Hoffmann, K.; Sydler, T.; Cattori, V.; Graf, F.; Diserens, K.A.; Padrutt, I.; et al. Molecular characterization and virus neutralization patterns of severe, non-epizootic forms of feline calicivirus infections resembling virulent systemic disease in cats in Switzerland and in Liechtenstein. Vet. Microbiol. 2016, 182, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Clay, S.; Maherchandani, S.; Malik, Y.S.; Goyal, S.M. Survival on uncommon fomites of feline calicivirus, a surrogate of noroviruses. Am. J. Infect. Control 2006, 34, 41–43. [Google Scholar] [CrossRef]
- Duizer, E.; Bijkerk, P.; Rockx, B.; De Groot, A.; Twisk, F.; Koopmans, M. Inactivation of caliciviruses. Appl. Environ. Microbiol. 2004, 70, 4538–4543. [Google Scholar] [CrossRef]
- Doultree, J.C.; Druce, J.D.; Birch, C.J.; Bowden, D.S.; Marshall, J.A. Inactivation of feline calicivirus, a Norwalk virus surrogate. J. Hosp. Infect. 1999, 41, 51–57. [Google Scholar] [CrossRef]
- Di Martino, B.; Ceci, C.; Di Profio, F.; Marsilio, F. In vitro inactivation of feline calicivirus (FCV) by chemical disinfectants: Resistance variation among field strains. Arch. Virol. 2010, 155, 2047–2051. [Google Scholar] [CrossRef] [PubMed]
- Scott, F.W. Virucidal disinfectants and feline viruses. Am. J. Vet. Res. 1980, 41, 410–414. [Google Scholar] [PubMed]
- Schorr-Evans, E.M.; Poland, A.; Johnson, W.E.; Pedersen, N.C. An epizootic of highly virulent feline calicivirus disease in a hospital setting in New England. J. Feline Med. Surg. 2003, 5, 217–226. [Google Scholar] [CrossRef]
- Hurley, K.E.; Pesavento, P.A.; Pedersen, N.C.; Poland, A.M.; Wilson, E.; Foley, J.E. An outbreak of virulent systemic feline calicivirus disease. J. Am. Vet. Med. Assoc. 2004, 224, 241–249. [Google Scholar] [CrossRef]
- Carducci, A.; Verani, M.; Lombardi, R.; Casini, B.; Privitera, G. Environmental survey to assess viral contamination of air and surfaces in hospital settings. J. Hosp. Infect. 2011, 77, 242–247. [Google Scholar] [CrossRef]
- Geret, C.; Riond, B.; Cattori, V.; Meli, M.; Hofmann-Lehmann, R.; Lutz, H. Housing and care of laboratory cats: From requirements to practice. Schweiz. Arch. Tierheilkd. 2011, 153, 157–164. [Google Scholar] [CrossRef]
- Museux, K.; Boretti, F.S.; Willi, B.; Riond, B.; Hoelzle, K.; Hoelzle, L.E.; Wittenbrink, M.M.; Tasker, S.; Wengi, N.; Reusch, C.E.; et al. In vivo transmission studies of ‘Candidatus Mycoplasma turicensis’ in the domestic cat. Vet. Res. 2009, 40, 45. [Google Scholar] [CrossRef]
- Helps, C.R.; Lait, P.; Damhuis, A.; Bjornehammar, U.; Bolta, D.; Brovida, C.; Chabanne, L.; Egberink, H.; Ferrand, G.; Fontbonne, A.; et al. Factors associated with upper respiratory tract disease caused by feline herpesvirus, feline calicivirus, Chlamydophila felis and Bordetella bronchiseptica in cats: Experience from 218 European catteries. Vet. Rec. 2005, 156, 669–673. [Google Scholar] [CrossRef]
- Lutz, H.; Pedersen, N.C.; Durbin, R.; Theilen, G.H. Monoclonal antibodies to three epitopic regions of feline leukemia virus p27 and their use in enzyme-linked immunosorbent assay of p27. J. Immunol. Methods 1983, 56, 209–220. [Google Scholar] [CrossRef]
- Soderlund, R.; Bolske, G.; Holst, B.S.; Aspan, A. Development and evaluation of a real-time polymerase chain reaction method for the detection of Mycoplasma felis. J. Vet. Diagn. Investig. 2011, 23, 890–893. [Google Scholar] [CrossRef]
- Troyer, R.M.; Beatty, J.A.; Stutzman-Rodriguez, K.R.; Carver, S.; Lozano, C.C.; Lee, J.S.; Lappin, M.R.; Riley, S.P.; Serieys, L.E.; Logan, K.A.; et al. Novel gammaherpesviruses in North American domestic cats, bobcats, and pumas: Identification, prevalence, and risk factors. J. Virol. 2014, 88, 3914–3924. [Google Scholar] [CrossRef] [PubMed]
- Novacco, M.; Kohan, N.R.; Stirn, M.; Meli, M.L.; Diaz-Sanchez, A.A.; Boretti, F.S.; Hofmann-Lehmann, R. Prevalence, Geographic Distribution, Risk Factors and Co-Infections of Feline Gammaherpesvirus Infections in Domestic Cats in Switzerland. Viruses 2019, 11, 721. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Lehmann, R.; Fehr, D.; Grob, M.; Elgizoli, M.; Packer, C.; Martenson, J.S.; O’Brien, S.J.; Lutz, H. Prevalence of antibodies to feline parvovirus, calicivirus, herpesvirus, coronavirus, and immunodeficiency virus and of feline leukemia virus antigen and the interrelationship of these viral infections in free-ranging lions in east Africa. Clin. Diagn. Lab. Immunol. 1996, 3, 554–562. [Google Scholar]
- Osterhaus, A.D.M.E.; Horzinek, M.C.; Reynolds, D.J. Seroepidemiology of feline infectious peritonitis virus-infections using transmissible gastroenteritis virus as antigen. Zentralbl. Veterinarmed. B 1977, 24, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.W.; Olmsted, R.A.; Martenson, J.S.; Obrien, S.J. Exposure to FIV and FIPV in wild and captive cheetahs. Zoo Biol. 1993, 12, 135–142. [Google Scholar] [CrossRef]
- Schweizerische Eidgenossenschaft. Schweizer Tierschutzgesetz (TSchG). Available online: https://www.admin.ch/opc/de/classified-compilation/20022103/index.html (accessed on 16 July 2019).
- Spiri, A.M.; Theze, J.; Meli, M.L.; Cattori, V.; Berger, A.; Steinrigl, A.; Pybus, O.G.; Hofmann-Lehmann, R.; Willi, B. Genetic diversity and phenotypic associations of feline caliciviruses from cats in Switzerland. J. Gen. Virol. 2016, 97, 3253–3266. [Google Scholar] [CrossRef] [PubMed]
- Sosnovtsev, S.V.; Prikhod’ko, E.A.; Belliot, G.; Cohen, J.I.; Green, K.Y. Feline calicivirus replication induces apoptosis in cultured cells. Virus Res. 2003, 94, 1–10. [Google Scholar] [CrossRef]
- Helps, C.; Lait, P.; Tasker, S.; Harbour, D. Melting curve analysis of feline calicivirus isolates detected by real-time reverse transcription PCR. J. Virol. Methods 2002, 106, 241–244. [Google Scholar] [CrossRef]
- Abd-Eldaim, M.M.; Wilkes, R.P.; Thomas, K.V.; Kennedy, M.A. Development and validation of a TaqMan real-time reverse transcription-PCR for rapid detection of feline calicivirus. Arch. Virol. 2009, 154, 555–560. [Google Scholar] [CrossRef]
- Meli, M.L.; Berger, A.; Willi, B.; Spiri, A.M.; Riond, B.; Hofmann-Lehmann, R. Molecular detection of feline calicivirus in clinical samples: A study comparing its detection by RT-qPCR directly from swabs and after virus isolation. J. Virol. Methods 2018, 251, 54–60. [Google Scholar] [CrossRef]
- Addie, D.D.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Horzinek, M.C.; Hosie, M.J.; Lloret, A.; Lutz, H.; et al. Disinfectant choices in veterinary practices, shelters and households: ABCD guidelines on safe and effective disinfection for feline environments. J. Feline Med. Surg. 2015, 17, 594–605. [Google Scholar] [CrossRef] [PubMed]
- Ecolab Healthcare. Incidin™ Plus, Flächendesinfektionsmittel mit Glucoprotamin, Aldehydfrei und Ohne QAV. Available online: https://www.ecolabhealthcare.de/website/seiten/produkte/flaechendesinfektion/konzentrate_aldehydfrei/incidin_plus.php (accessed on 17 July 2019).
- Lee, K.M.; Gillespie, J.H. Thermal and pH stability of feline calicivirus. Infect. Immun. 1973, 7, 678–679. [Google Scholar] [PubMed]
- Buckley, D. Survival, Recovery, and Inactivation of Human Norovirus Surrogates, Feline Calicivirus and Murine Norovirus, on Carpets. All Dissertations 2017, 1979. Available online: https://tigerprints.clemson.edu/all_dissertations/1979 (accessed on 17 July 2019).
- Rice, C.C.; Kruger, J.M.; Venta, P.J.; Vilnis, A.; Maas, K.A.; Dulin, J.A.; Maes, R.K. Genetic characterization of 2 novel feline caliciviruses isolated from cats with idiopathic lower urinary tract disease. J. Vet. Intern. Med. 2002, 16, 293–302. [Google Scholar] [CrossRef]
- Larson, J.; Kruger, J.M.; Wise, A.G.; Kaneene, J.B.; Miller, R.; Fitzgerald, S.D.; Kiupel, M.; Maes, R.K. Nested case-control study of feline calicivirus viruria, oral carriage, and serum neutralizing antibodies in cats with idiopathic cystitis. J. Vet. Intern. Med. 2011, 25, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Kruger, J.M.; Osborne, C.A.; Venta, P.J.; Sussman, M.D. Viral infections of the feline urinary tract. Vet. Clin. N. Am. Small Anim. Pract. 1996, 26, 281–296. [Google Scholar] [CrossRef]
- Addie, D.; Houe, L.; Maitland, K.; Passantino, G.; Decaro, N. Effect of cat litters on feline coronavirus infection of cell culture and cats. J. Feline Med. Surg. 2019. [Google Scholar] [CrossRef]
- Gassilloud, B.; Schwartzbrod, L.; Gantzer, C. Presence of viral genomes in mineral water: A sufficient condition to assume infectious risk? Appl. Environ. Microbiol. 2003, 69, 3965–3969. [Google Scholar] [CrossRef]
- Nuanualsuwan, S.; Cliver, D.O. Capsid functions of inactivated human picornaviruses and feline calicivirus. Appl. Environ. Microbiol. 2003, 69, 350–357. [Google Scholar] [CrossRef]
- Malik, Y.S.; Goyal, S.M. Virucidal efficacy of sodium bicarbonate on a food contact surface against feline calicivirus, a norovirus surrogate. Int. J. Food Microbiol. 2006, 109, 160–163. [Google Scholar] [CrossRef]
- Yang, H.; Kendall, P.A.; Medeiros, L.; Sofos, J.N. Inactivation of Listeria monocytogenes, Escherichia coli O157:H7, and Salmonella typhimurium with compounds available in households. J. Food Prot. 2009, 72, 1201–1208. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Days Post FCV Challenge I | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cat ID | −4 | 3 | 9 | 15 | 22 | 29 | 36 | 43 | 50 | 56 | 63 | 71 | 78 | 85 | 92 | 99 | 106 | 371 |
| Cat JJG4 | ||||||||||||||||||
| Cat JJG6 | ||||||||||||||||||
| Cat JJH3 | ||||||||||||||||||
| Cat JJI1 | ||||||||||||||||||
| Cat JJI2 | ||||||||||||||||||
| Cat JJF1 | ||||||||||||||||||
| Cat JJG3 | ||||||||||||||||||
| Cat JJH2 | ||||||||||||||||||
| Cat JJI3 | ||||||||||||||||||
| Cat JJI4 | ||||||||||||||||||
| Days after Challenge | −4 | 1 | 3 | 5 | 7 | 8 | 9 | 11 | 13 | 15 | 22 | 29 | 36 | 43 | 50 | 56 | 63 | 71 | 78 | 85 | 92 | 99 | 106 | 371 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Water bowl | 34.4 | 35.9 | 36.0 | 34.1 | 34.2 | 33.5 | 36.4 | 36.0 | 33.8 | 38.5 | nt | nt | 38.5 | |||||||||||
| Water | 38.9 | 37.0 | 38.6 | 36.8 | nt | nt | ||||||||||||||||||
| Litter tray 1 | 37.9 | 39.0 | 37.2 | 38.7 | 37.9 | nt | nt | |||||||||||||||||
| Litter tray 2 | 37.2 | 32.3 | 38.5 | nt | nt | |||||||||||||||||||
| Transfer area | 34.3 | 36.1 | 35.5 | 34.5 | 35.4 | 33.1 | 33.7 | 33.9 | 36.8 | 36.1 | 35.3 | 36.4 | 35.7 | 37.4 | 37.7 | 38.9 | nt | nt | 39.4 | 38.2 | ||||
| Food bowl 1 | 33.4 | 35.0 | 33.6 | 36.4 | 34.6 | 34.1 | 34.3 | 38.7 | 37.4 | 37.3 | 37.0 | 39.9 | nt | nt | 37.9 | |||||||||
| Food bowl 2 | 30.9 | 33.7 | 33.7 | 36.8 | 34.9 | 36.4 | 33.6 | 35.4 | 37.0 | 38.0 | 38.6 | 37.9 | 38.6 | nt | nt | |||||||||
| Ventilation filter | 34.8 | 32.6 | 39.2 | 34.2 | 35.9 | 36.0 | 36.0 | 36.2 | 36.7 | 37.4 | 37.9 | 36.1 | 37.1 | 38.1 | nt | nt | ||||||||
| Floor food storage room | nt | nt | ||||||||||||||||||||||
| Hair cat JJH3 | nt | nt | nt | nt | nt | nt | nt | nt | nt | nt | nt | nt | nt | nt | nt | nt | 38.7 | nt |
| Days Post FCV Challenge II | ||||||
|---|---|---|---|---|---|---|
| Cat ID | −4 | 3 | 9 | 15 | 22 | 42 |
| Cat JJG4 | ||||||
| Cat JJG6 | ||||||
| Cat JJH3 | ||||||
| Cat JJI1 | ||||||
| Cat JJI2 | ||||||
| Cat JJF1 | ||||||
| Cat JJG3 | ||||||
| Cat JJH2 | ||||||
| Cat JJI3 | ||||||
| Cat JJI4 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spiri, A.M.; Meli, M.L.; Riond, B.; Herbert, I.; Hosie, M.J.; Hofmann-Lehmann, R. Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility. Viruses 2019, 11, 958. https://doi.org/10.3390/v11100958
Spiri AM, Meli ML, Riond B, Herbert I, Hosie MJ, Hofmann-Lehmann R. Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility. Viruses. 2019; 11(10):958. https://doi.org/10.3390/v11100958
Chicago/Turabian StyleSpiri, Andrea Monika, Marina Luisa Meli, Barbara Riond, Imogen Herbert, Margaret J. Hosie, and Regina Hofmann-Lehmann. 2019. "Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility" Viruses 11, no. 10: 958. https://doi.org/10.3390/v11100958
APA StyleSpiri, A. M., Meli, M. L., Riond, B., Herbert, I., Hosie, M. J., & Hofmann-Lehmann, R. (2019). Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility. Viruses, 11(10), 958. https://doi.org/10.3390/v11100958