Characterization of Two Pseudomonas aeruginosa Viruses vB_PaeM_SCUT-S1 and vB_PaeM_SCUT-S2


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Bacteriophage Isolation, Propagation, and Purification
2.3. Host Range Analysis
2.4. Electron Microscopy
2.5. Temperature and pH Stability
2.6. Genome DNA Extraction and Sequencing
2.7. Genome Assembly, Annotation, and Comparison
2.8. Proteomics Analysis
2.9. One-Step Growth Curve and Lysis Kinetics
2.10. Biofilm Inhibition Assays
2.11. Accession Number
3. Results
3.1. Isolation and Characterization of the Phages
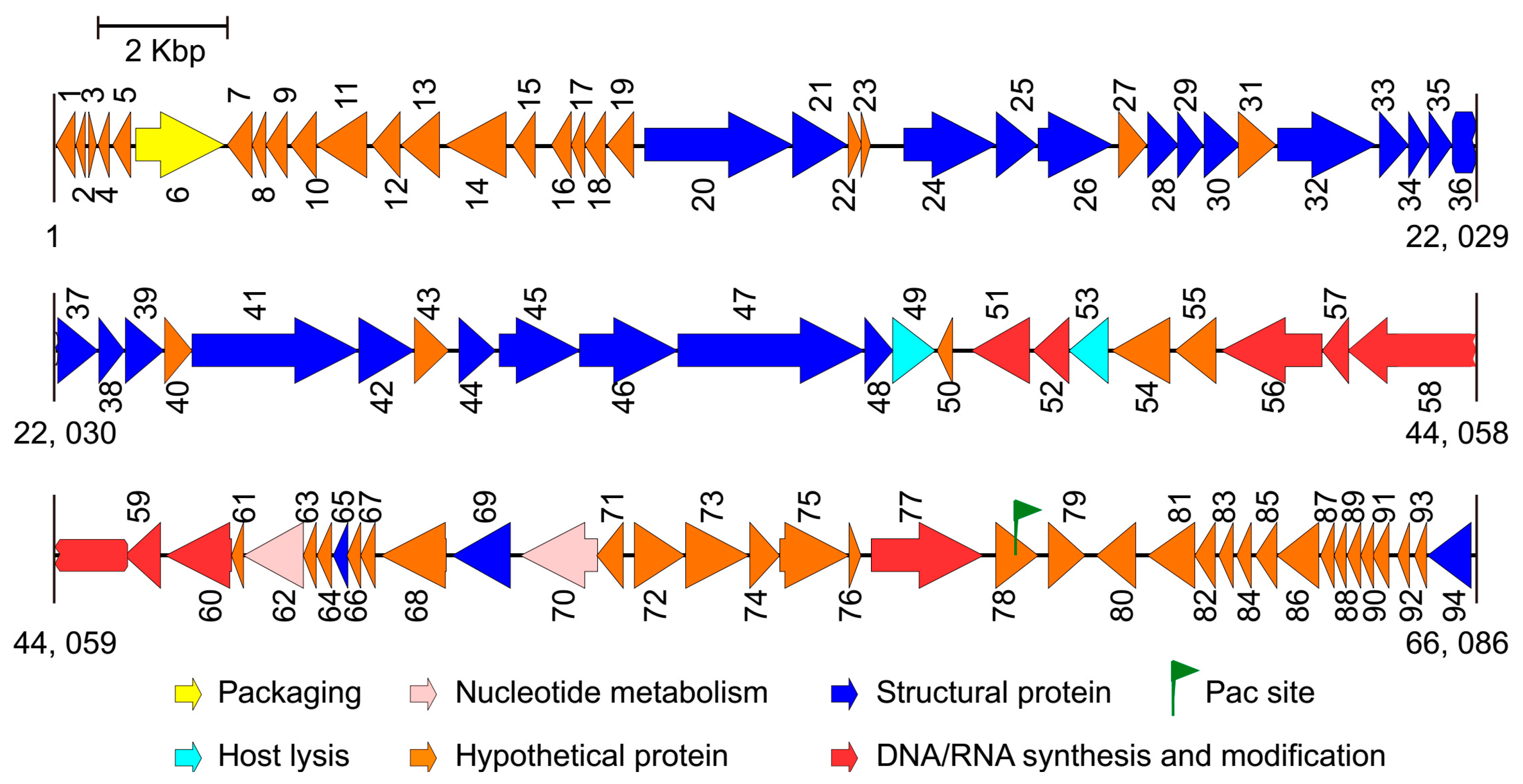
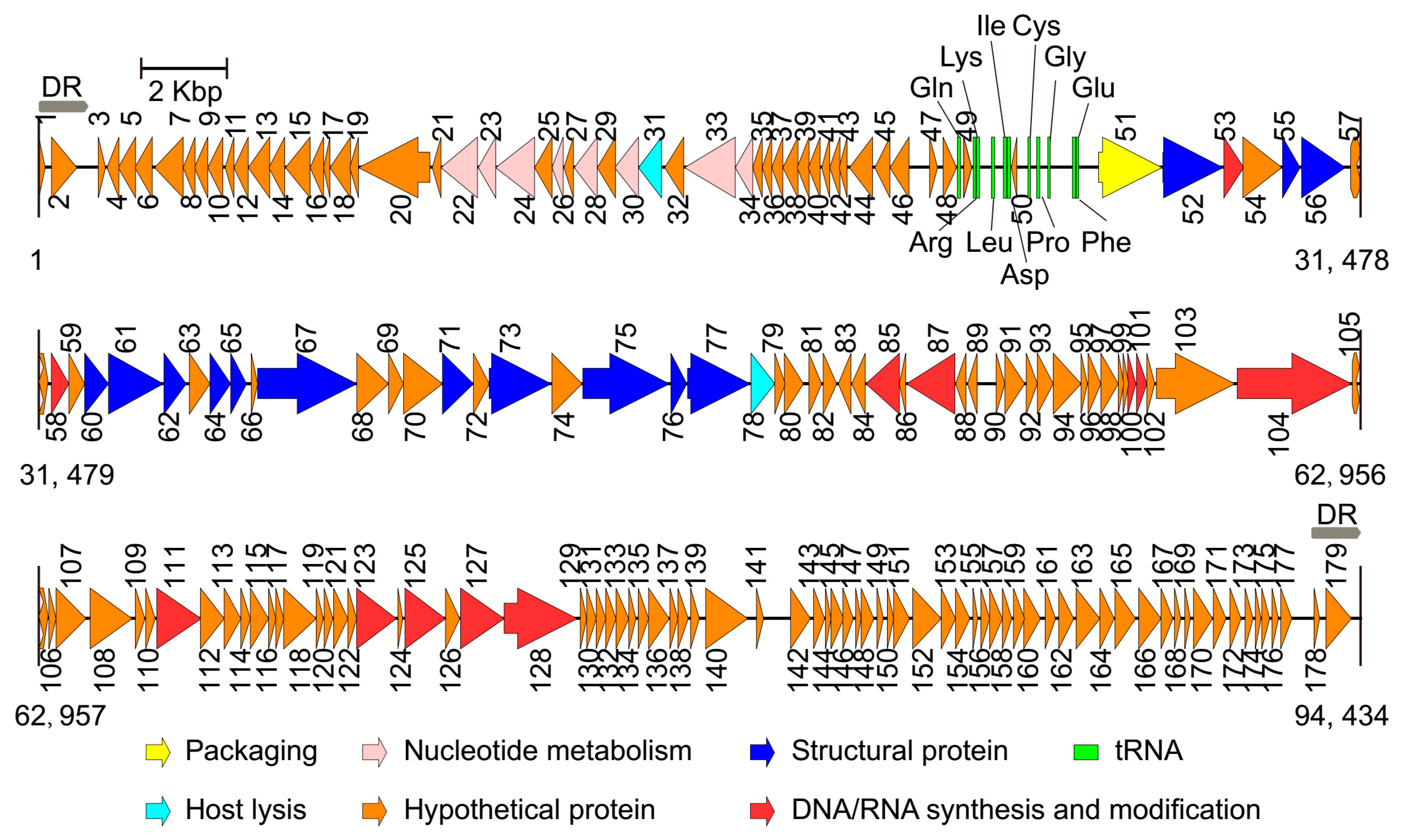
3.2. Basic Characteristics of the Genomes
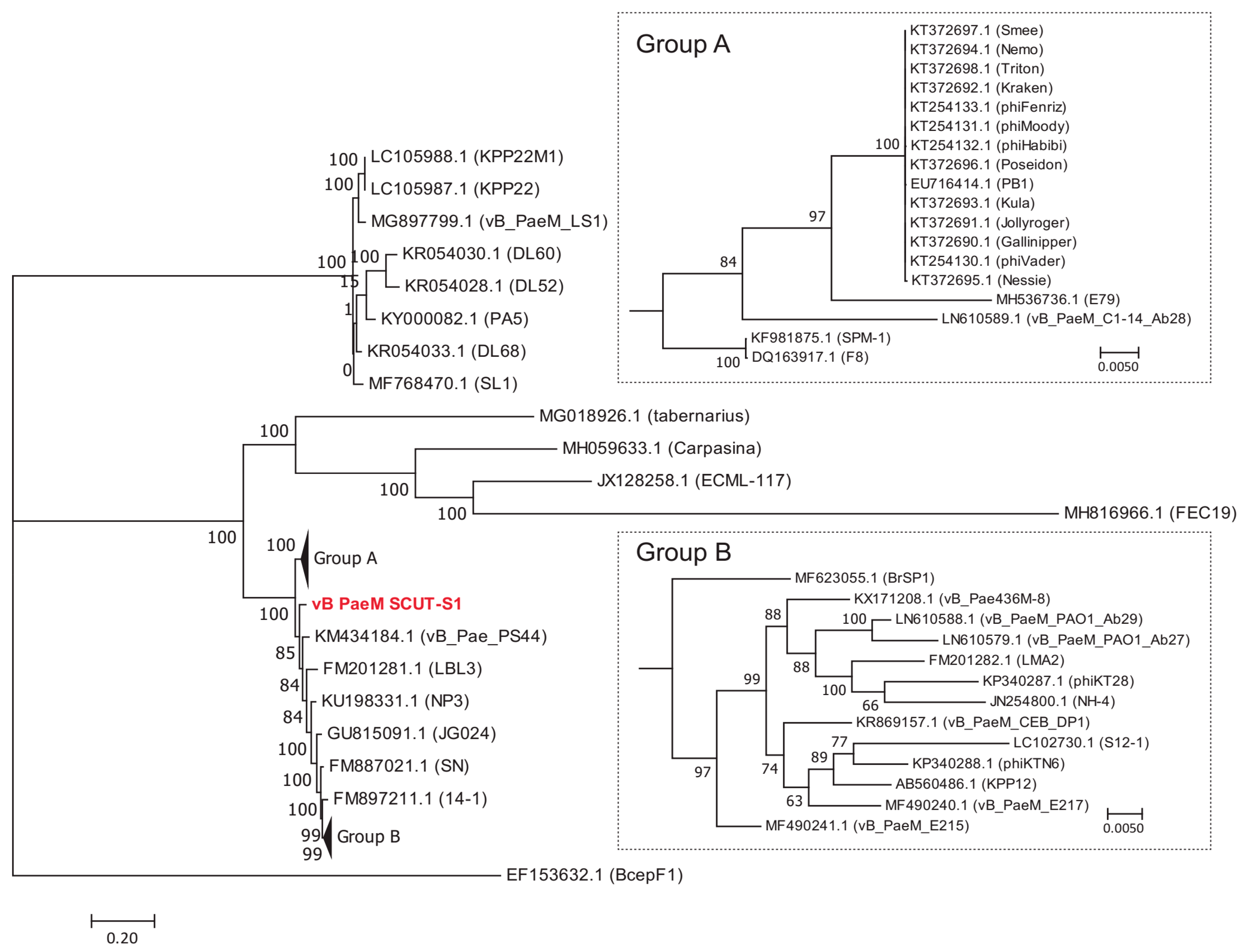
3.3. Comparative Genome Analysis
3.4. Structural Proteins of the Two Phages
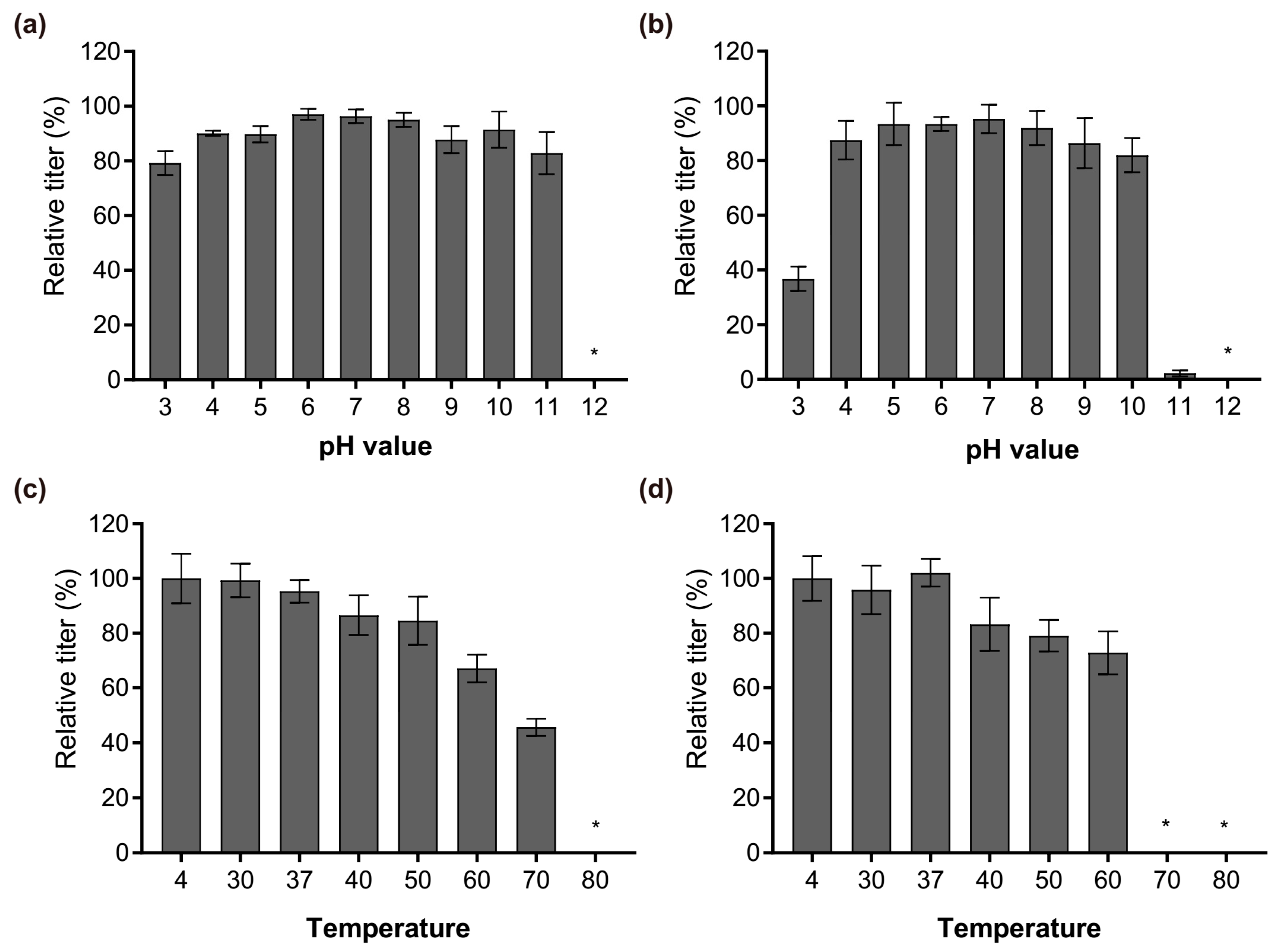
3.5. Stability Analysis
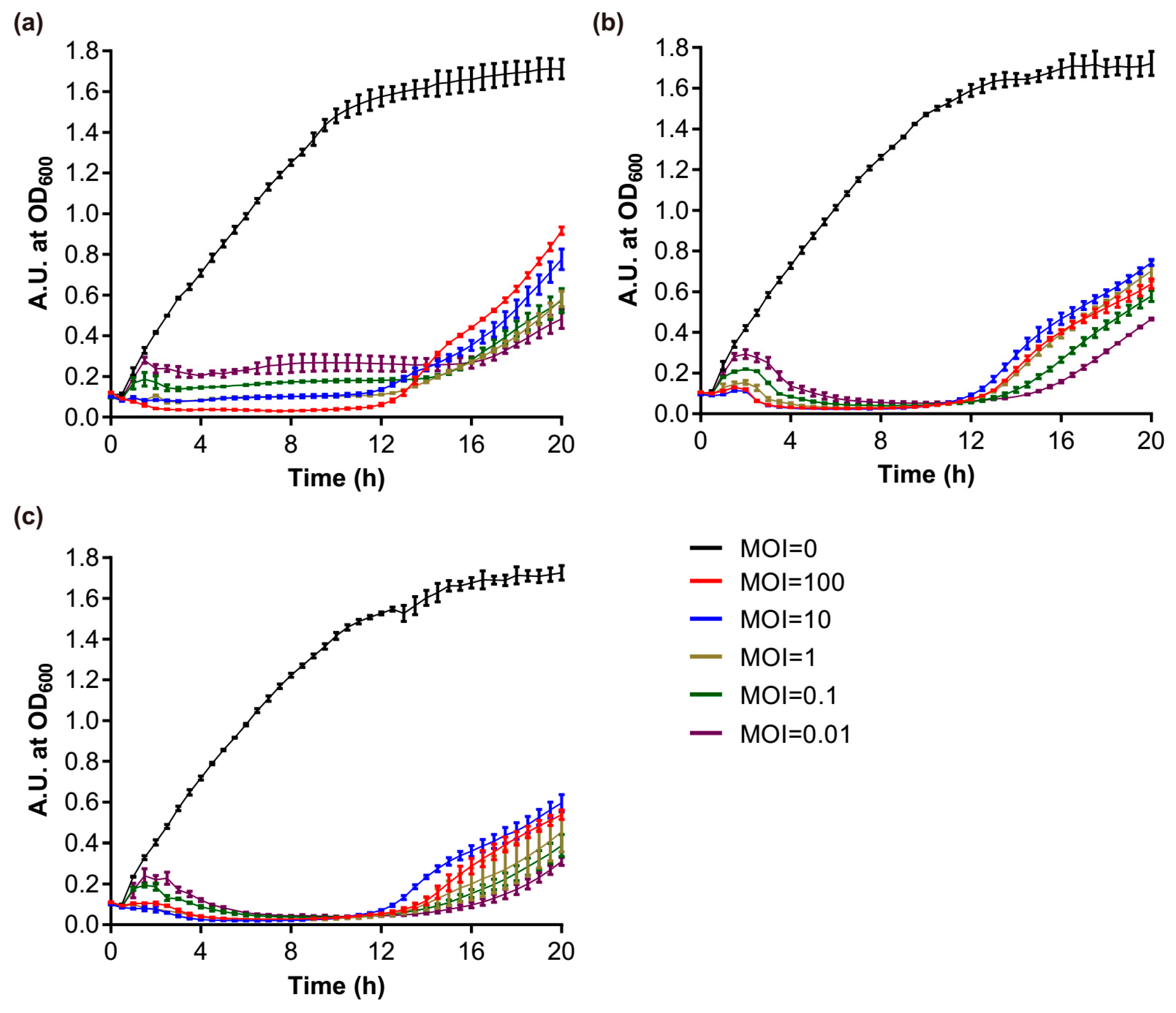
3.6. Growth Characteristics and Lysis Kinetics
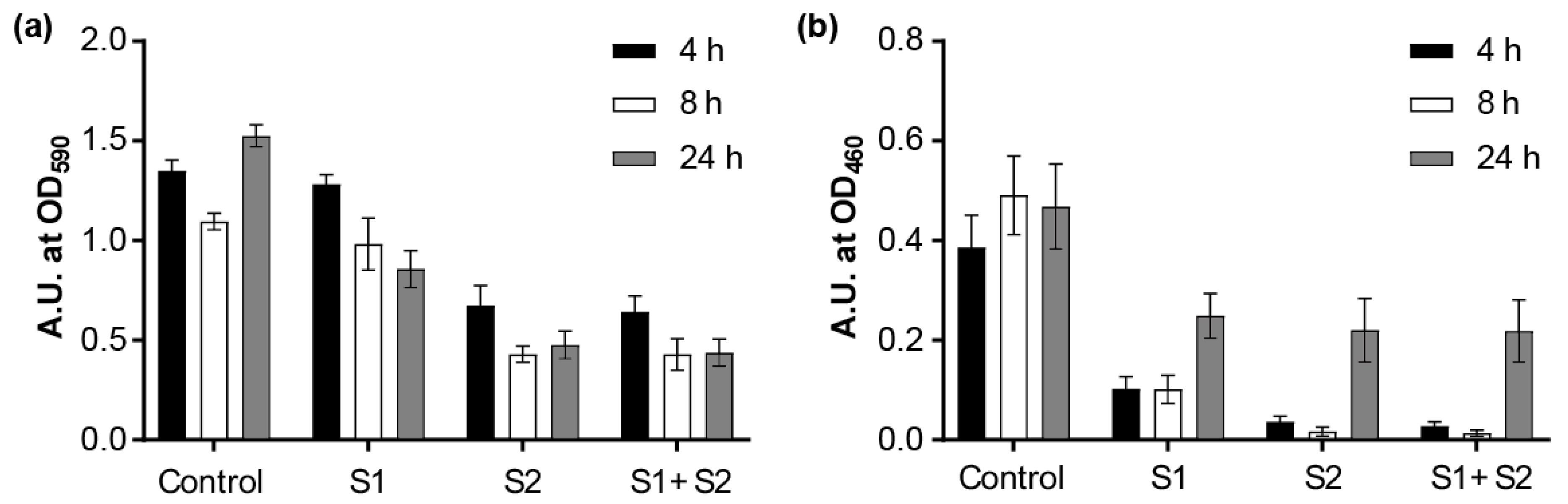
3.7. Biofilm Eradication
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No eskape! An update from the infectious diseases society of america. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the eskape pathogens. Expert Rev. Anti Infect. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in eskape pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef]
- Emerson, J.; Rosenfeld, M.; McNamara, S.; Ramsey, B.; Gibson, R.L. Pseudomonas aeruginosa and other predictors of mortality and morbidity in young children with cystic fibrosis. Pediatric Pulmonol. 2002, 34, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of pseudomonas aeruginosa infection: Lessons from a versatile opportunist. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar] [CrossRef]
- Mesaros, N.; Nordmann, P.; Plésiat, P.; Roussel-Delvallez, M.; Van Eldere, J.; Glupczynski, Y.; Van Laethem, Y.; Jacobs, F.; Lebecque, P.; Malfroot, A.; et al. Pseudomonas aeruginosa: Resistance and therapeutic options at the turn of the new millennium. Clin. Microbiol. Infect. 2007, 13, 560–578. [Google Scholar] [CrossRef]
- Lister, P.D.; Wolter, D.J.; Hanson, N.D. Antibacterial-resistant pseudomonas aeruginosa: Clinical impact and complex regulation of chromosomally encoded resistance mechanisms. Clin. Microbiol. Rev. 2009, 22, 582–610. [Google Scholar] [CrossRef]
- Chatterjee, M.; Anju, C.P.; Biswas, L.; Anil Kumar, V.; Gopi Mohan, C.; Biswas, R. Antibiotic resistance in pseudomonas aeruginosa and alternative therapeutic options. Int. J. Med. Microbiol. 2016, 306, 48–58. [Google Scholar] [CrossRef]
- Wagner, S.; Sommer, R.; Hinsberger, S.; Lu, C.; Hartmann, R.W.; Empting, M.; Titz, A. Novel strategies for the treatment of pseudomonas aeruginosa infections. J. Med. Chem. 2016, 59, 5929–5969. [Google Scholar] [CrossRef] [PubMed]
- Reardon, S. Phage therapy gets revitalized. Nature 2014, 510, 15–16. [Google Scholar] [CrossRef]
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2014, 1, 111–114. [Google Scholar] [CrossRef]
- Dublanchet, A.; Fruciano, E. A short history of phage therapy. Med. Et Mal. Infect. 2008, 38, 415–420. [Google Scholar] [CrossRef]
- Summers, W.C. Bacteriophage research: Early history. In Bacteriophages: Biology and Applications; CRC Press: Boca Raton, FL, USA, 2005; pp. 5–27. [Google Scholar]
- Chan, B.K.; Abedon, S.T.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef]
- Aziz, R.K.; Ackermann, H.W.; Petty, N.K.; Kropinski, A.M. Essential steps in characterizing bacteriophages: Biology, taxonomy, and genome analysis. Methods Mol. Biol. 2018, 1681, 197–215. [Google Scholar]
- McVay, C.S.; Velasquez, M.; Fralick, J.A. Phage therapy of pseudomonas aeruginosa infection in a mouse burn wound model. Antimicrob. Agents Chemother. 2007, 51, 1934–1938. [Google Scholar] [CrossRef]
- Kutter, E.; De Vos, D.; Gvasalia, G.; Alavidze, Z.; Gogokhia, L.; Kuhl, S.; Abedon, S. Phage therapy in clinical practice: Treatment of human infections. Curr. Pharm. Biotechnol. 2010, 11, 69–86. [Google Scholar] [CrossRef]
- De Smet, J.; Hendrix, H.; Blasdel, B.G.; Danis-Wlodarczyk, K.; Lavigne, R. Pseudomonas predators: Understanding and exploiting phage–host interactions. Nat. Rev. Microbiol. 2017, 15, 517–530. [Google Scholar] [CrossRef]
- Weber-Dabrowska, B.; Jonczyk-Matysiak, E.; Zaczek, M.; Lobocka, M.; Lusiak-Szelachowska, M.; Gorski, A. Bacteriophage procurement for therapeutic purposes. Front. Microbiol. 2016, 7, 1177. [Google Scholar] [CrossRef]
- Azeredo, J.; Sillankorva, S.; Pires, D.P. Pseudomonas bacteriophage isolation and production. Methods Mol. Biol. 2014, 1149, 23–32. [Google Scholar]
- Garbe, J.; Wesche, A.; Bunk, B.; Kazmierczak, M.; Selezska, K.; Rohde, C.; Sikorski, J.; Rohde, M.; Jahn, D.; Schobert, M. Characterization of jg024, a pseudomonas aeruginosa pb1-like broad host range phage under simulated infection conditions. BMC Microbiol. 2010, 10, 301. [Google Scholar] [CrossRef]
- Garbe, J.; Bunk, B.; Rohde, M.; Schobert, M. Sequencing and characterization of pseudomonas aeruginosa phage jg004. BMC Microbiol. 2011, 11, 102. [Google Scholar] [CrossRef]
- Russell, J.F.S.D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Kutter, E. Phage host range and efficiency of plating. Methods Mol. Biol. 2009, 501, 141–149. [Google Scholar]
- Essoh, C.; Blouin, Y.; Loukou, G.; Cablanmian, A.; Lathro, S.; Kutter, E.; Thien, H.V.; Vergnaud, G.; Pourcel, C. The susceptibility of pseudomonas aeruginosa strains from cystic fibrosis patients to bacteriophages. PLoS ONE 2013, 8, e60575. [Google Scholar] [CrossRef]
- Park, M.; Lee, J.H.; Shin, H.; Kim, M.; Choi, J.; Kang, D.H.; Heu, S.; Ryu, S. Characterization and comparative genomic analysis of a novel bacteriophage, sfp10, simultaneously inhibiting both salmonella enterica and escherichia coli o157:H7. Appl. Environ. Microbiol. 2012, 78, 58–69. [Google Scholar] [CrossRef]
- Thurber, R.V.; Haynes, M.; Breitbart, M.; Wegley, L.; Rohwer, F. Laboratory procedures to generate viral metagenomes. Nat. Protoc. 2009, 4, 470–483. [Google Scholar] [CrossRef]
- Jackman, S.D.; Vandervalk, B.P.; Mohamadi, H.; Chu, J.; Yeo, S.; Hammond, S.A.; Jahesh, G.; Khan, H.; Coombe, L.; Warren, R.L.; et al. Abyss 2.0: Resource-efficient assembly of large genomes using a bloom filter. Genome Res. 2017, 27, 768–777. [Google Scholar] [CrossRef]
- Chevreux, B.W.; Wetter, T.; Suhai, S. Genome sequence assembly using trace signals and additional sequence information. In Proceedings of the German Conference on Bioinformatics (GCB), Hannover, Germany, 4–6 October 1999; Volume 99, pp. 45–56. [Google Scholar]
- Garneau, J.R.; Depardieu, F.; Fortier, L.C.; Bikard, D.; Monot, M. Phageterm: A tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. Rasttk: A modular and extensible implementation of the rast algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The seed and the rapid annotation of microbial genomes using subsystems technology (rast). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The rast server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Delcher, A.L.; Bratke, K.A.; Powers, E.C.; Salzberg, S.L. Identifying bacterial genes and endosymbiont DNA with glimmer. Bioinformatics 2007, 23, 673–679. [Google Scholar] [CrossRef]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. Genemarks: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef]
- Lowe, T.M.; Chan, P.P. Trnascan-se on-line: Integrating search and context for analysis of transfer rna genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Nakamura, T.; Yamada, K.D.; Tomii, K.; Katoh, K. Parallelization of mafft for large-scale multiple sequence alignments. Bioinformatics 2018, 34, 2490–2492. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Stamatakis, A. Raxml version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. Mega7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Boulanger, P. Purification of bacteriophages and sds-page analysis of phage structural proteins from ghost particles. Methods Mol. Biol. 2009, 502, 227–238. [Google Scholar]
- Kropinski, A.M. Practical advice on the one-step growth curve. Methods Mol. Biol. 2018, 1681, 41–47. [Google Scholar]
- Danis-Wlodarczyk, K.; Olszak, T.; Arabski, M.; Wasik, S.; Majkowska-Skrobek, G.; Augustyniak, D.; Gula, G.; Briers, Y.; Jang, H.B.; Vandenheuvel, D.; et al. Characterization of the newly isolated lytic bacteriophages ktn6 and kt28 and their efficacy against pseudomonas aeruginosa biofilm. PLoS ONE 2015, 10, e0127603. [Google Scholar]
- Coffey, B.M.; Anderson, G.G. Biofilm formation in the 96-well microtiter plate. Methods Mol. Biol. 2014, 1149, 631–641. [Google Scholar]
- Smith, K.; Perez, A.; Ramage, G.; Gemmell, C.G.; Lang, S. Comparison of biofilm-associated cell survival following in vitro exposure of meticillin-resistant staphylococcus aureus biofilms to the antibiotics clindamycin, daptomycin, linezolid, tigecycline and vancomycin. Int. J. Antimicrob. Agents 2009, 33, 374–378. [Google Scholar] [CrossRef]
- Chaturongakul, S.; Ounjai, P. Phage–host interplay: Examples from tailed phages and gram-negative bacterial pathogens. Front. Microbiol. 2014, 5, 442. [Google Scholar] [CrossRef]
- Parmar, K.M.; Gaikwad, S.L.; Dhakephalkar, P.K.; Kothari, R.; Singh, R.P. Intriguing interaction of bacteriophage-host association: An understanding in the era of omics. Front. Microbiol. 2017, 8, 559. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Lemire, S.; Pires, D.P.; Lu, T.K. Engineering modular viral scaffolds for targeted bacterial population editing. Cell Syst. 2015, 1, 187–196. [Google Scholar] [CrossRef]
- Le, S.; He, X.; Tan, Y.; Huang, G.; Zhang, L.; Lux, R.; Shi, W.; Hu, F. Mapping the tail fiber as the receptor binding protein responsible for differential host specificity of pseudomonas aeruginosa bacteriophages pap1 and jg004. PLoS ONE 2013, 8, e68562. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Phage evolution and ecology. Adv. Appl. Microbiol. 2009, 67, 1–45. [Google Scholar]
- Deghorain, M.; Van Melderen, L. The staphylococci phages family: An overview. Viruses 2012, 4, 3316–3335. [Google Scholar] [CrossRef] [PubMed]
- Essoh, C.; Latino, L.; Midoux, C.; Blouin, Y.; Loukou, G.; Nguetta, S.P.; Lathro, S.; Cablanmian, A.; Kouassi, A.K.; Vergnaud, G.; et al. Investigation of a large collection of pseudomonas aeruginosa bacteriophages collected from a single environmental source in abidjan, cote d’ivoire. PLoS ONE 2015, 10, e0130548. [Google Scholar] [CrossRef]
- Young, R.; Gill, J.J. Microbiology. Phage therapy redux—What is to be done? Science 2015, 350, 1163–1164. [Google Scholar] [CrossRef]
- Rohde, C.; Resch, G.; Pirnay, J.P.; Blasdel, B.G.; Debarbieux, L.; Gelman, D.; Gorski, A.; Hazan, R.; Huys, I.; Kakabadze, E.; et al. Expert opinion on three phage therapy related topics: Bacterial phage resistance, phage training and prophages in bacterial production strains. Viruses 2018, 10, 178. [Google Scholar] [CrossRef] [PubMed]
- Betts, A.; Vasse, M.; Kaltz, O.; Hochberg, M.E. Back to the future: Evolving bacteriophages to increase their effectiveness against the pathogen pseudomonas aeruginosa pao1. Evol. Appl. 2013, 6, 1054–1063. [Google Scholar]
- Uchiyama, J.; Suzuki, M.; Nishifuji, K.; Kato, S.I.; Miyata, R.; Nasukawa, T.; Yamaguchi, K.; Takemura-Uchiyama, I.; Ujihara, T.; Shimakura, H.; et al. Analyses of short-term antagonistic evolution of pseudomonas aeruginosa strain pao1 and phage kpp22 (myoviridae family, pb1-like virus genus). Appl. Environ. Microbiol. 2016, 82, 4482–4491. [Google Scholar] [CrossRef] [PubMed]
- Rossitto, M.; Fiscarelli, E.V.; Rosati, P. Challenges and promises for planning future clinical research into bacteriophage therapy against pseudomonas aeruginosa in cystic fibrosis. An argumentative review. Front. Microbiol. 2018, 9, 775. [Google Scholar] [CrossRef]
- Vieira, A.; Silva, Y.J.; Cunha, A.; Gomes, N.C.; Ackermann, H.W.; Almeida, A. Phage therapy to control multidrug-resistant pseudomonas aeruginosa skin infections: In vitro and ex vivo experiments. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 3241–3249. [Google Scholar] [CrossRef] [PubMed]
- Alemayehu, D.; Casey, P.G.; McAuliffe, O.; Guinane, C.M.; Martin, J.G.; Shanahan, F.; Coffey, A.; Ross, R.P.; Hill, C. Bacteriophages phimr299-2 and phinh-4 can eliminate pseudomonas aeruginosa in the murine lung and on cystic fibrosis lung airway cells. mBio 2012, 3, e00029-12. [Google Scholar] [CrossRef]
- Beeton, M.L.; Alves, D.R.; Enright, M.C.; Jenkins, A.T. Assessing phage therapy against pseudomonas aeruginosa using a galleria mellonella infection model. Int. J. Antimicrob Agents 2015, 46, 196–200. [Google Scholar] [CrossRef]
- Debarbieux, L.; Leduc, D.; Maura, D.; Morello, E.; Criscuolo, A.; Grossi, O.; Balloy, V.; Touqui, L. Bacteriophages can treat and prevent pseudomonas aeruginosa lung infections. J. Infect. Dis. 2010, 201, 1096–1104. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Lavigne, R.; Debarbieux, L. Predicting in vivo efficacy of therapeutic bacteriophages used to treat pulmonary infections. Antimicrob Agents Chemother. 2013, 57, 5961–5968. [Google Scholar] [CrossRef]
- Ceyssens, P.J.; Miroshnikov, K.; Mattheus, W.; Krylov, V.; Robben, J.; Noben, J.P.; Vanderschraeghe, S.; Sykilinda, N.; Kropinski, A.M.; Volckaert, G.; et al. Comparative analysis of the widespread and conserved pb1-like viruses infecting pseudomonas aeruginosa. Environ. Microbiol. 2009, 11, 2874–2883. [Google Scholar] [CrossRef]
- Latz, S.; Kruttgen, A.; Hafner, H.; Buhl, E.M.; Ritter, K.; Horz, H.P. Differential effect of newly isolated phages belonging to pb1-like, phikz-like and luz24-like viruses against multi-drug resistant pseudomonas aeruginosa under varying growth conditions. Viruses 2017, 9, 315. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Predicted Function | Gene No. | Mol. Mass (kDa) | No. of Peptides | Coverage |
|---|---|---|---|---|---|
| 1 | putative tail protein containing transglycosylase | ORF41 | 94.39 | 72 | 81% |
| 2 | putative tail fiber protein | ORF47 | 103.16 | 44 | 63% |
| 3 | putative minor head protein | ORF20 | 84.53 | 44 | 59% |
| 4 | putative structural protein | ORF46 | 54.81 | 23 | 60% |
| 5 | putative major structural protein | ORF26 | 41.59 | 20 | 49% |
| 6 | putative structural protein | ORF32 | 53.82 | 19 | 47% |
| 7 | putative minor head protein | ORF21 | 31.72 | 16 | 56% |
| 8 | putative structural protein | ORF24 | 52.10 | 14 | 38% |
| 9 | putative tail fiber protein | ORF33 | 15.91 | 12 | 70% |
| 10 | putative structural protein | ORF42 | 32.60 | 12 | 58% |
| 11 | putative structural protein | ORF31 | 21.48 | 12 | 77% |
| 12 | putative baseplate protein | ORF45 | 43.51 | 11 | 41% |
| 13 | putative endolysin | ORF49 | 24.35 | 11 | 48% |
| 14 | putative structural protein | ORF25 | 21.57 | 9 | 59% |
| 15 | putative structural protein | ORF38 | 19.94 | 9 | 60% |
| 16 | putative structural protein | ORF54 | 32.21 | 8 | 25% |
| 17 | putative structural protein | ORF37 | 17.80 | 7 | 67% |
| 18 | putative structural protein | ORF28 | 16.96 | 7 | 30% |
| 19 | putative structural protein | ORF22 | 7.46 | 7 | 77% |
| 20 | putative tail fiber protein | ORF39 | 21.70 | 6 | 44% |
| 21 | putative structural protein | ORF36 | 14.46 | 6 | 50% |
| 22 | putative structural protein | ORF35 | 12.74 | 5 | 29% |
| 23 | unknown function protein | ORF84 | 8.61 | 4 | 47% |
| 24 | putative holin | ORF53 | 21.13 | 4 | 25% |
| 25 | putative structural protein | ORF27 | 16.37 | 4 | 22% |
| 26 | putative DNA helicase | ORF56 | 59.43 | 4 | 7% |
| 27 | putative structural protein | ORF34 | 12.36 | 3 | 42% |
| 28 | putative ATP-dependent exonuclease V | ORF70 | 45.54 | 3 | 7% |
| 29 | putative baseplate protein | ORF44 | 24.17 | 3 | 13% |
| 30 | unknown function protein | ORF72 | 29.47 | 3 | 10% |
| 31 | unknown function protein | ORF11 | 15.52 | 2 | 21% |
| No. | Predicted Function | Gene No. | Mol. Mass (kDa) | No. of Peptides | Coverage |
|---|---|---|---|---|---|
| 1 | putative tape measure protein | ORF67 | 85.86 | 55 | 66% |
| 2 | putative structural protein | ORF52 | 54.49 | 38 | 77% |
| 3 | putative structural protein | ORF61 | 46.35 | 31 | 51% |
| 4 | major capsid protein | ORF56 | 39.41 | 30 | 72% |
| 5 | putative tail fiber protein | ORF75 | 71.97 | 29 | 69% |
| 6 | putative tail fiber protein | ORF77 | 53.15 | 23 | 58% |
| 7 | putative baseplate component | ORF73 | 52.37 | 17 | 36% |
| 8 | putative structural protein | ORF74 | 26.64 | 17 | 80% |
| 9 | putative structural protein | ORF55 | 14.85 | 14 | 90% |
| 10 | putative baseplate protein | ORF71 | 26.84 | 11 | 53% |
| 11 | putative structural protein | ORF60 | 21.27 | 10 | 43% |
| 12 | putative structural protein | ORF70 | 34.17 | 10 | 35% |
| 13 | putative structural protein | ORF68 | 28.60 | 10 | 49% |
| 14 | putative structural protein | ORF57 | 18.24 | 9 | 40% |
| 15 | putative structural protein | ORF62 | 18.95 | 8 | 55% |
| 16 | putative structural protein | ORF63 | 18.29 | 8 | 40% |
| 17 | putative structural protein | ORF72 | 14.07 | 7 | 32% |
| 18 | putative structural protein | ORF59 | 14.46 | 7 | 73% |
| 19 | unknown function protein | ORF108 | 37.10 | 6 | 17% |
| 20 | putative RNA polymerase | ORF58 | 15.51 | 5 | 15% |
| 21 | putative structural protein | ORF80 | 16.06 | 4 | 32% |
| 22 | putative endolysin | ORF78 | 20.89 | 4 | 20% |
| 23 | putative ribonucleoside-diphosphate reductase alpha chain | ORF128 | 67.42 | 4 | 6% |
| 24 | putative ribonucleoside-diphosphate reductase beta subunit | ORF127 | 40.62 | 4 | 8% |
| 25 | putative 3’-phosphatase | ORF123 | 35.43 | 3 | 9% |
| 26 | putative nictotinate phosphoribosyltransferase | ORF20 | 63.05 | 3 | 5% |
| 27 | unknown function protein | ORF14 | 14.86 | 3 | 6% |
| 28 | putative structural protein | ORF64 | 17.75 | 2 | 12% |
| 29 | putative methyltransferase | ORF53 | 17.27 | 2 | 14% |
| 30 | putative structural protein | ORF69 | 13.98 | 2 | 20% |
| 31 | unknown function protein | ORF140 | 37.78 | 2 | 4% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Chen, P.; Lin, Z.; Wang, T. Characterization of Two Pseudomonas aeruginosa Viruses vB_PaeM_SCUT-S1 and vB_PaeM_SCUT-S2. Viruses 2019, 11, 318. https://doi.org/10.3390/v11040318
Guo Y, Chen P, Lin Z, Wang T. Characterization of Two Pseudomonas aeruginosa Viruses vB_PaeM_SCUT-S1 and vB_PaeM_SCUT-S2. Viruses. 2019; 11(4):318. https://doi.org/10.3390/v11040318
Chicago/Turabian StyleGuo, Yangyijun, Ping Chen, Zhanglin Lin, and Tingting Wang. 2019. "Characterization of Two Pseudomonas aeruginosa Viruses vB_PaeM_SCUT-S1 and vB_PaeM_SCUT-S2" Viruses 11, no. 4: 318. https://doi.org/10.3390/v11040318
APA StyleGuo, Y., Chen, P., Lin, Z., & Wang, T. (2019). Characterization of Two Pseudomonas aeruginosa Viruses vB_PaeM_SCUT-S1 and vB_PaeM_SCUT-S2. Viruses, 11(4), 318. https://doi.org/10.3390/v11040318