Effects of Intranasal Pseudorabies Virus AH02LA Infection on Microbial Community and Immune Status in the Ileum and Colon of Piglets

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Design and Sample Collection
2.2. Pseudorabies Virus (PRV) DNA Detection in the Ileal and Colonic Mucosa
2.3. Gene Expression Analysis in Ileal and Colonic Mucosa
2.4. sIgA Concentration Analyses in the Ileal and Colonic Digesta
2.5. DNA Extraction, MiSeq Sequencing and Bioinformatics Analyses
2.6. SCFA and Lactate Concentrations Analysis in the Ileum and Colon
2.7. Statistical Analysis
3. Results
3.1. Pseudorabies Virus (PRV) DNA Detection in the Ileal and Colonic Mucosa
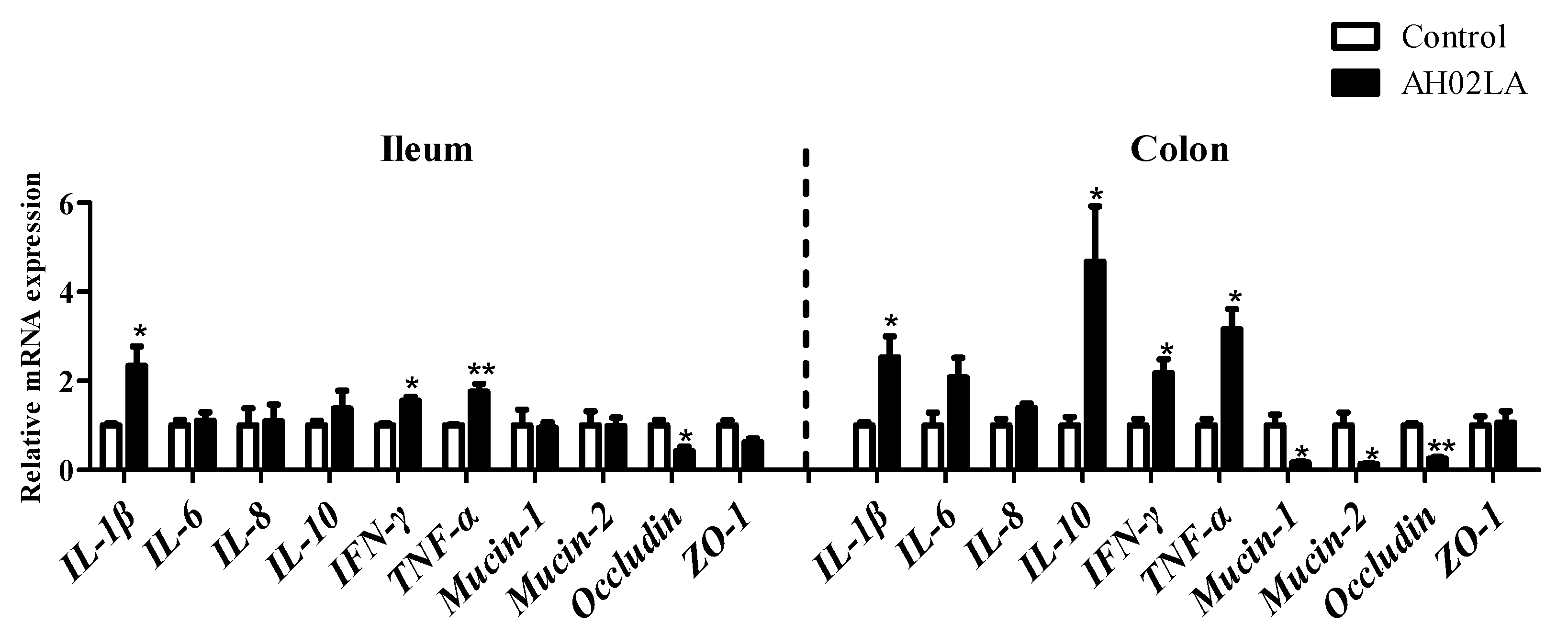
3.2. Gene Expression in Ileal and Colonic Mucosa
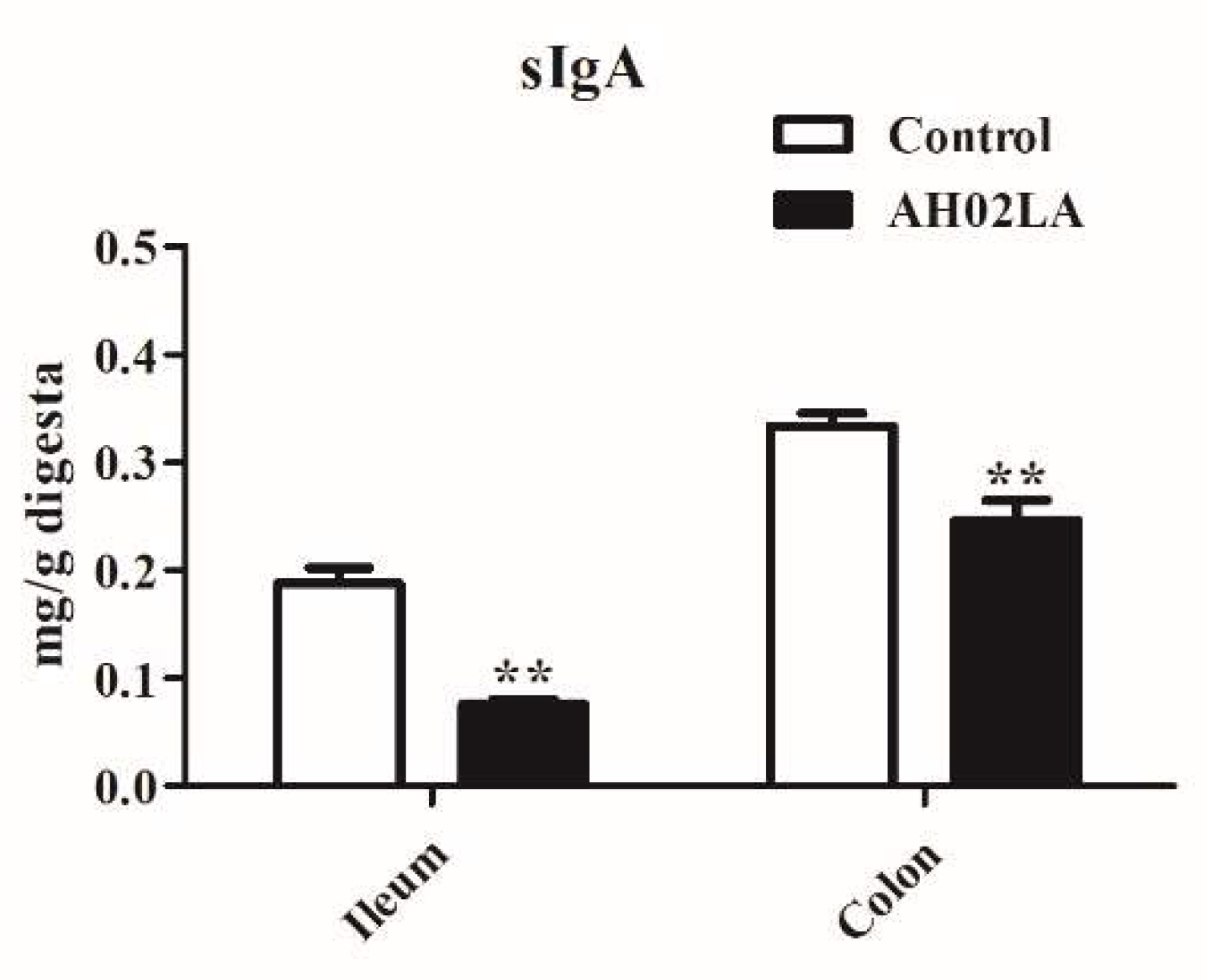
3.3. sIgA Concentration in the Ileum and Colon
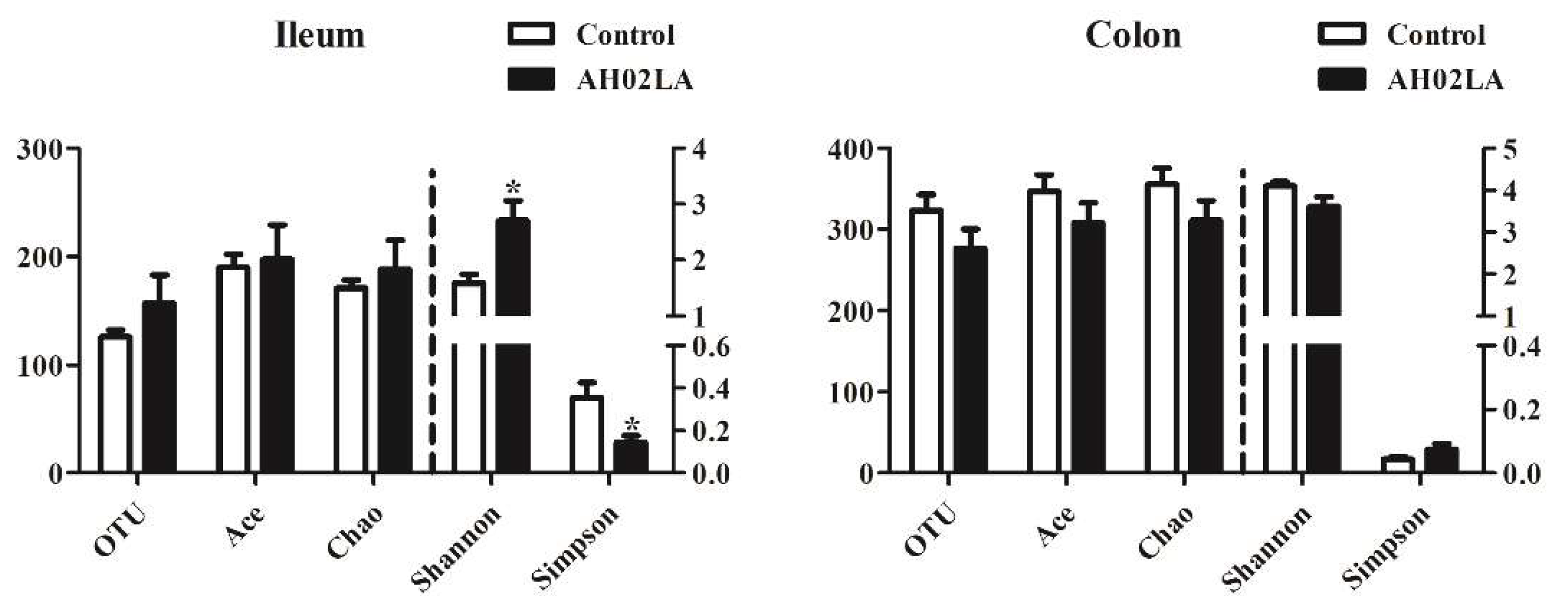
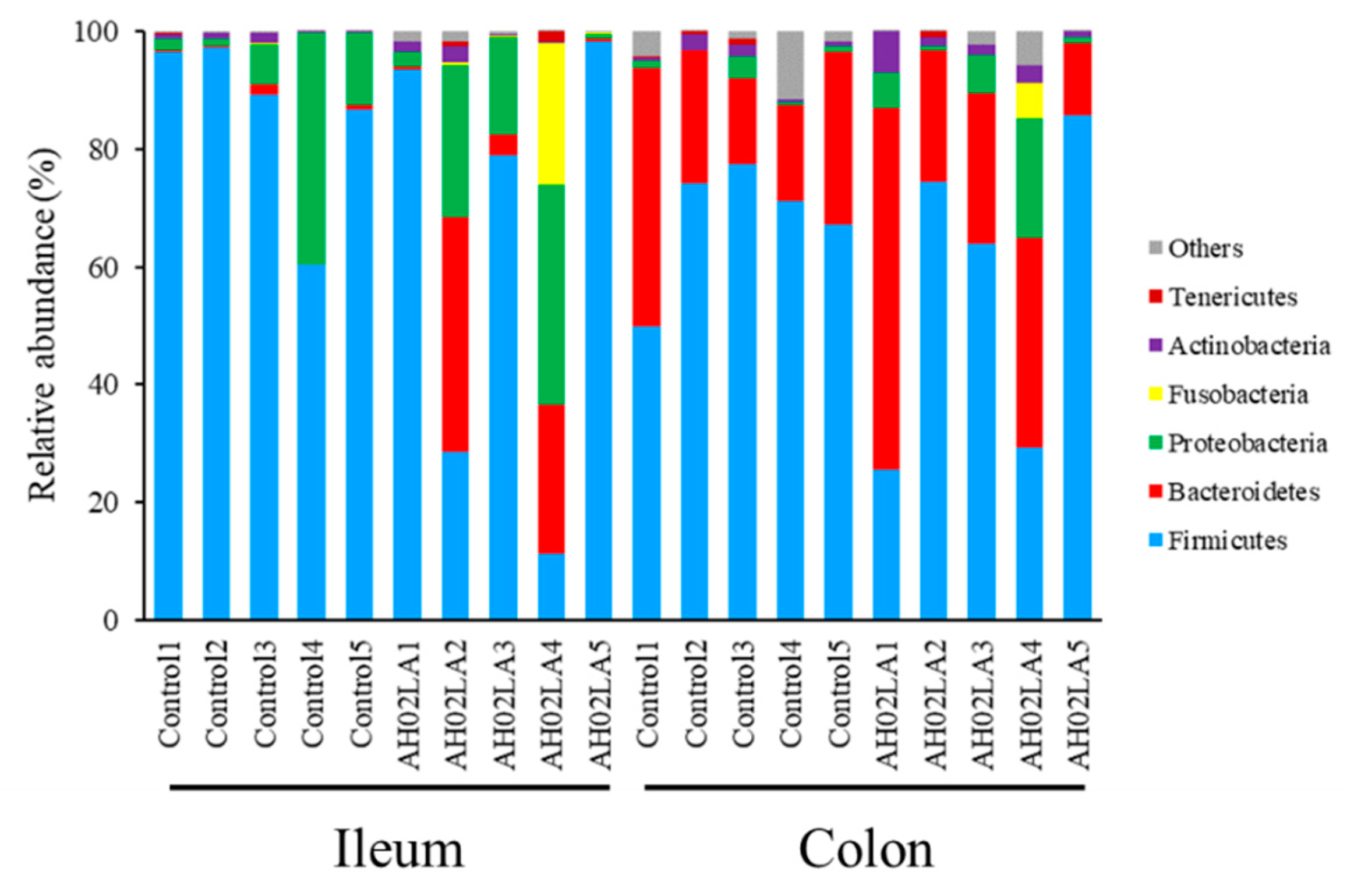
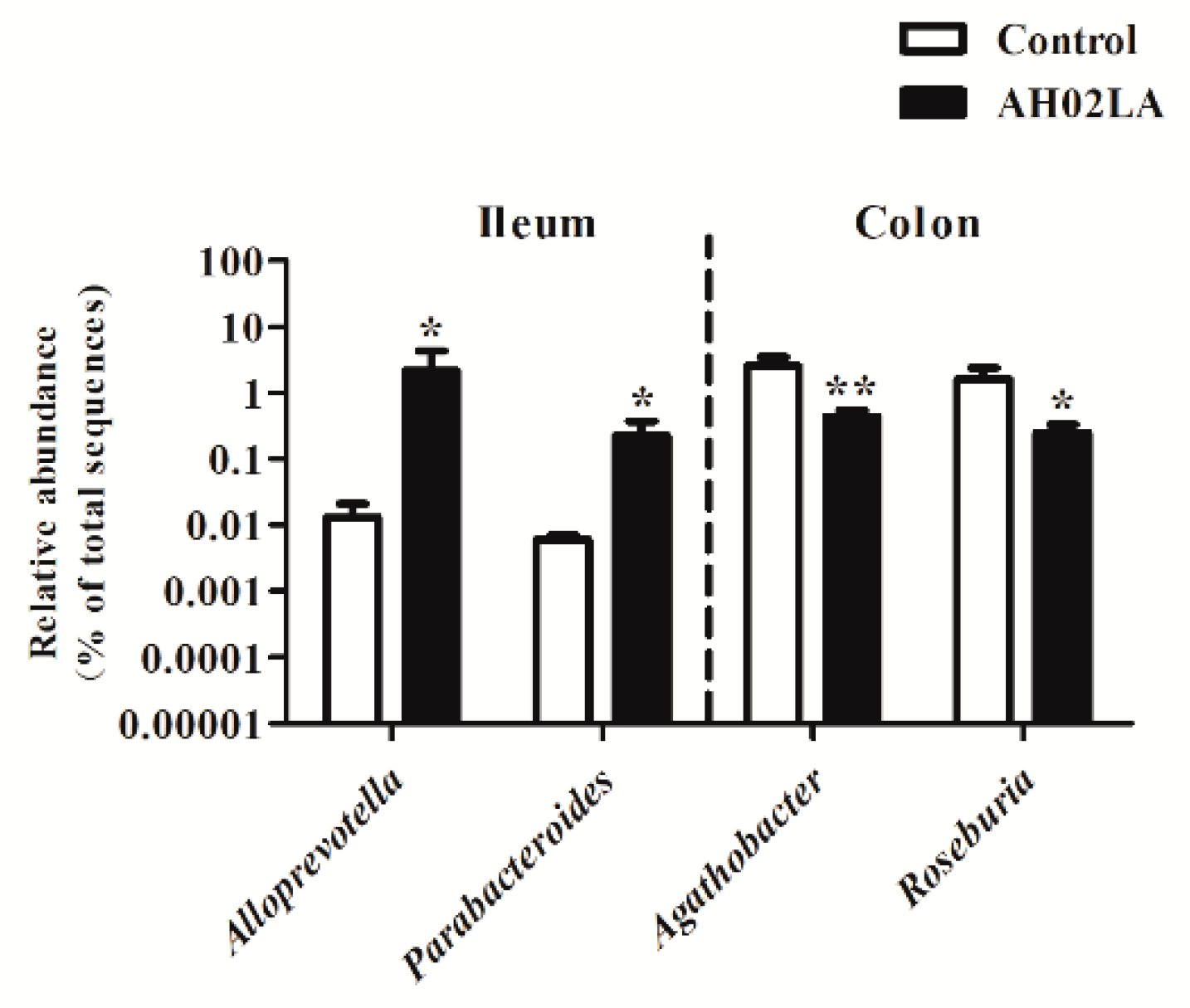
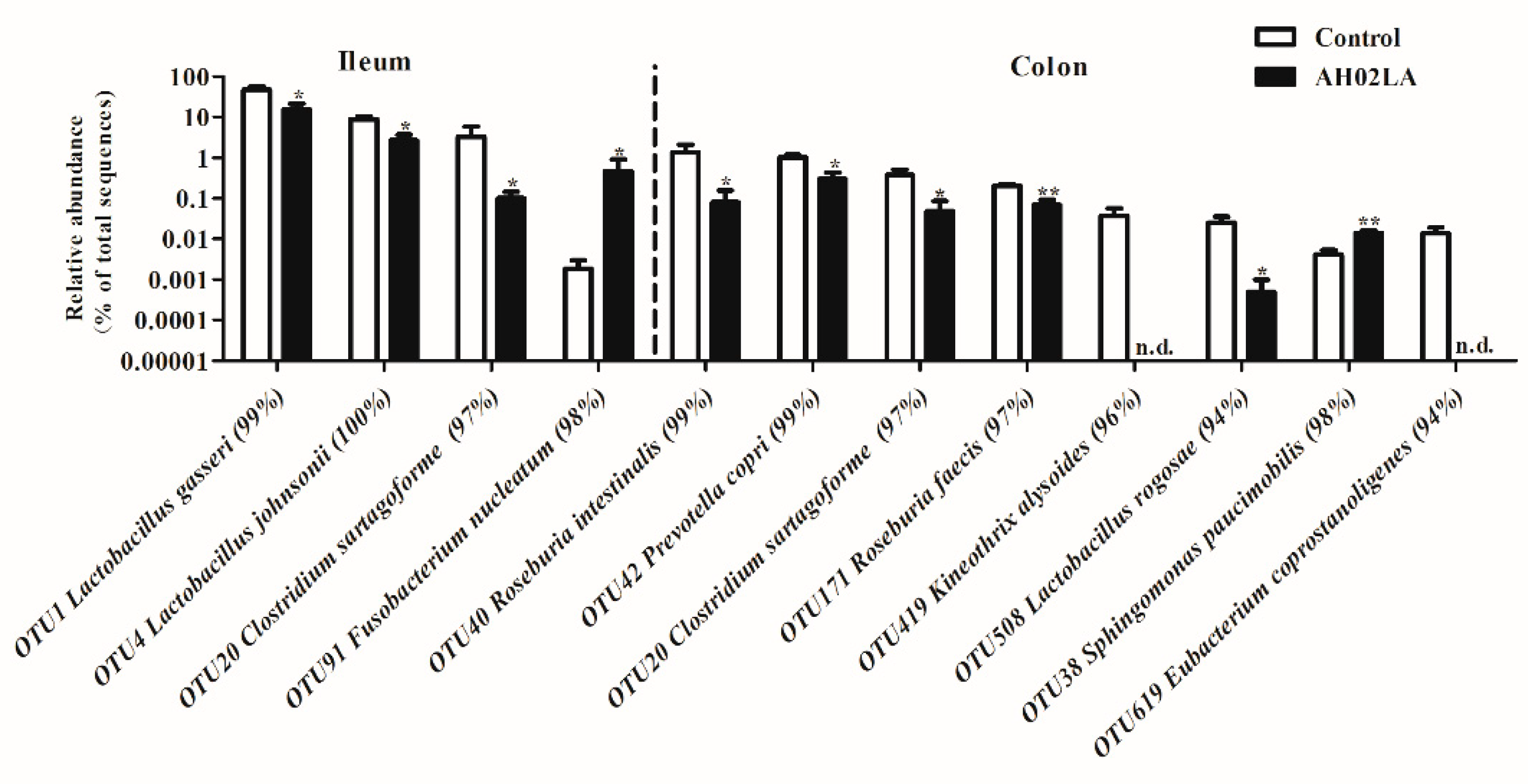
3.4. Bacterial Community in the Ileal and Colonic Digesta
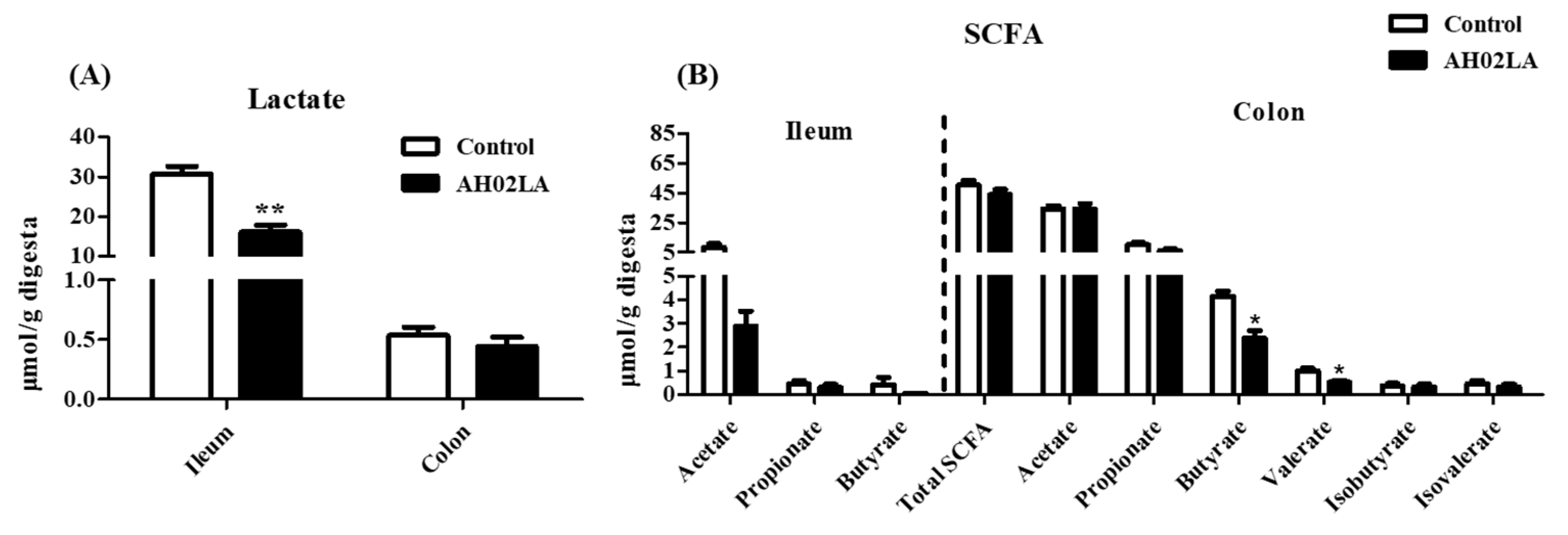
3.5. Short-Chain Fatty Acids (SCFA) and Lactate Concentrations in Ileal and Colonic Digesta
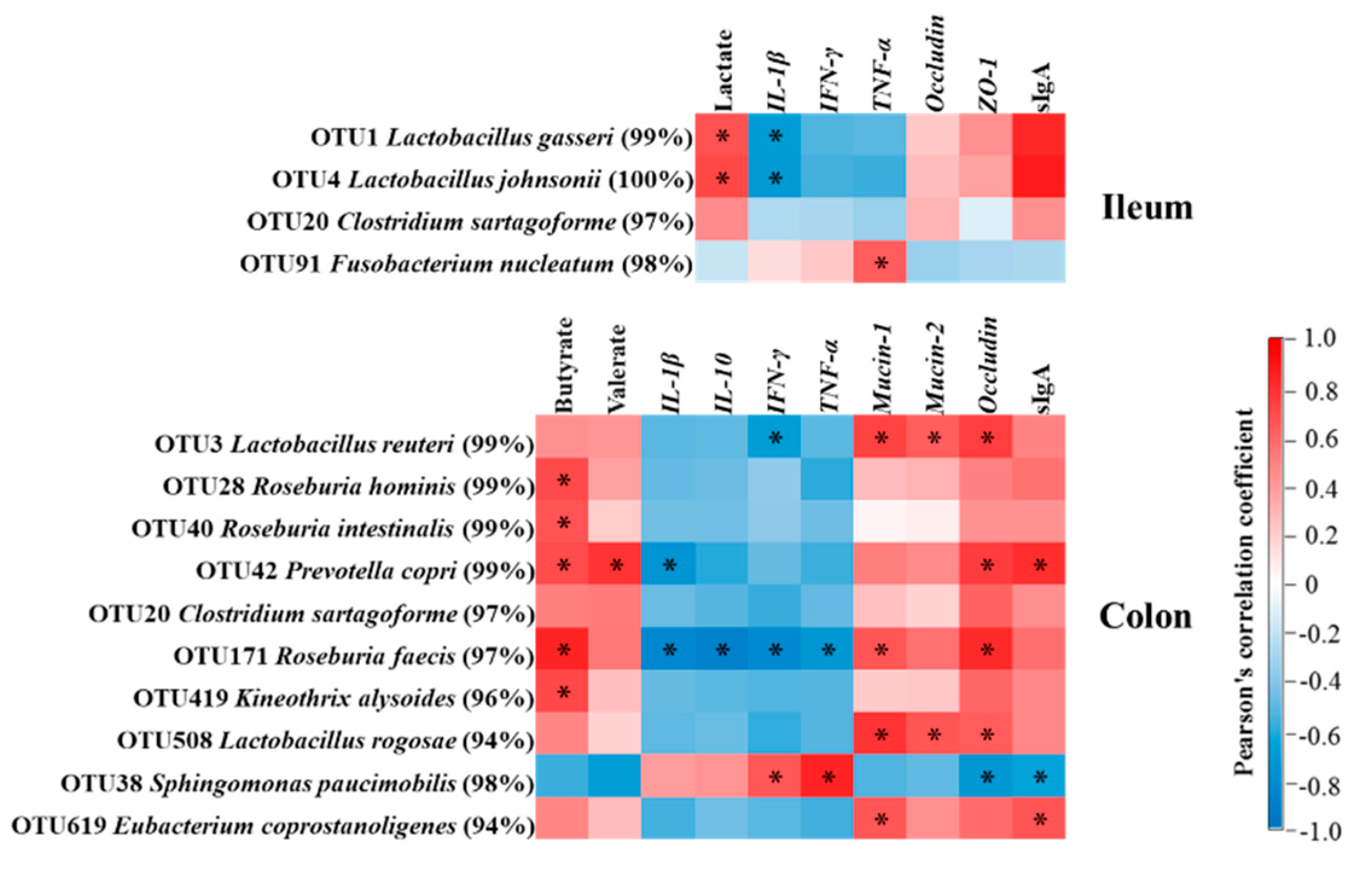
3.6. Correlation Analysis of Immune Markers, Bacteria, and Microbial Fermentation Products in the Ileum and Colon and Body Temperature
4. Discussion
4.1. Intranasal Pseudorabies Virus (PRV) AH02LA Infection Influences Some Immune Markers in the Ileum and Colon
4.2. Intranasal Pseudorabies Virus (PRV) AH02LA Infection Alters the Microbiota and Their Metabolites in the Ileum and Colon
4.3. Bacterial Composition Change Are Associated with Altered Immune Markers and Body Temperature
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wang, G.S.; Du, Y.; Wu, J.Q.; Tian, F.L.; Yu, X.J.; Wang, J.B. Vaccine resistant pseudorabies virus causes mink infection in China. BMC Vet. Res. 2018, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- An, T.Q.; Peng, J.M.; Tian, Z.J.; Zhao, H.Y.; Li, N.; Liu, Y.M.; Chen, J.Z.; Leng, C.L.; Sun, Y.; Chang, D.; et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012. Emerg. Infect. Dis. 2013, 19, 1749–1755. [Google Scholar] [CrossRef] [PubMed]
- Mettenleiter, T.C. Aujeszky’s disease (pseudorabies) virus: The virus and molecular pathogenesis--state of the art, June 1999. Vet. Res. 2000, 31, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Bai, C.; Sun, J.; Chang, S.; Zhang, X. Emergence of virulent pseudorabies virus infection in northern China. J. Vet. Sci. 2013, 14, 363–365. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, N.; Cong, X.; Wang, C.H.; Du, M.; Li, L.; Zhao, B.; Yuan, J.; Liu, D.D.; Li, S.; et al. Pathogenicity and genomic characterization of a pseudorabies virus variant isolated from Bartha-K61-vaccinated swine population in China. Vet. Microbiol. 2014, 174, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Verpoest, S.; Cay, B.; Favoreel, H.; De Regge, N. Age-dependent differences in pseudorabies virus neuropathogenesis and associated cytokine expression. J. Virol. 2017, 91, e02058-16. [Google Scholar] [CrossRef] [PubMed]
- Verpoest, S.; Redant, V.; Cay, A.B.; Favoreel, H.; De, N.R. Reduced virulence of a pseudorabies virus isolate from wild boar origin in domestic pigs correlates with hampered visceral spread and age-dependent reduced neuroinvasive capacity. Virulence 2018, 9, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Goto, Y.; Ivanov, I.I. Intestinal epithelial cells as mediators of the commensal-host immune crosstalk. Immunol. Cell Biol. 2013, 91, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Holmes, E.; Wilson, I.D. Gut microorganisms, mammalian metabolism and personalized health care. Nat. Rev. Microbiol. 2005, 3, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Dan, R.L.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef] [PubMed]
- Suiryanrayna, M.V.A.N.; Ramana, J.V. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2016, 6, 45. [Google Scholar] [CrossRef] [PubMed]
- Pflughoeft, K.J.; Versalovic, J. Human microbiome in health and disease. Annu. Rev. Pathol. 2012, 7, 99–122. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Seo, S.U.; Chen, G.Y.; Núñez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Koh, H.W.; Kim, M.S.; Lee, J.S.; Kim, H.; Park, S.J. Changes in the swine gut microbiota in response to porcine epidemic diarrhea infection. Microbes Environ. 2015, 30, 758–759. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Yang, Y.; Wang, J.; Jing, Y.; Yang, Q. Impact of TGEV infection on the pig small intestine. Virol. J. 2018, 15, 102. [Google Scholar] [CrossRef] [PubMed]
- Narita, M.; Kubo, M.; Fukusho, A.; Haritani, M.; Moriwaki, M. Necrotizing enteritis in piglets associated with Aujeszky’s disease virus infection. Vet. Pathol. 1984, 21, 450–452. [Google Scholar] [CrossRef]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667. [Google Scholar] [CrossRef]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felix, S.; Fredrik, B.C. The gut microbiota-masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar]
- Wang, J.; Guo, R.; Qiao, Y.; Xu, M.; Wang, Z.; Liu, Y.; Gu, Y.; Liu, C.; Hou, J. An inactivated gE-deleted pseudorabies vaccine provides complete clinical protection and reduces virus shedding against challenge by a Chinese pseudorabies variant. BMC Vet. Res. 2016, 12, 277. [Google Scholar] [CrossRef]
- Qiao, Y.; Gu, Y.; Liu, C.; Xu, M.; Wang, Z.; Liu, Y.; Hou, J.; Wang, J. Isolation and identification of one porcine pseudorabies virus strain AH02LA and its pathogenicity in piglets. Anim. Husb. Vet. Med. 2016, 48, 36–41. [Google Scholar]
- Joo, H.S.; Donaldson-Wood, C.R.; Johnson, R.H. A standardised haemagglutination inhibition test for porcine parvovirus antibody. Aust. Vet. J. 2010, 52, 422–424. [Google Scholar] [CrossRef]
- Liu, S.; Zhao, Y.; Hu, Q.; Lv, C.; Zhang, C.; Zhao, R.; Hu, F.; Lin, W.; Cui, S. A multiplex RT-PCR for rapid and simultaneous detection of porcine teschovirus, classical swine fever virus, and porcine reproductive and respiratory syndrome virus in clinical specimens. J. Virol. Methods 2011, 172, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Yue, F.; Cui, S.; Zhang, C.; Yoon, K. A multiplex PCR for rapid and simultaneous detection of porcine circovirus type 2, porcine parvovirus, porcine pseudorabies virus, and porcine reproductive and respiratory syndrome virus in clinical specimens. Virus Genes 2009, 38, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; Akkermans, A.D.; De Vos, W.M. Temperature gradient gel electrophoresis analysis of 16S rRNA from human fecal samples reveals stable and host-specific communities of active bacteria. Appl. Environ. Microbiol. 1998, 64, 3854. [Google Scholar]
- Mao, S.; Zhang, M.; Liu, J.; Zhu, W. Characterising the bacterial microbiota across the gastrointestinal tracts of dairy cattle: Membership and potential function. Sci. Rep. 2015, 5, 16116. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Pi, Y.; Peng, Y.; Mu, C.L.; Zhu, W.Y. Time-course responses of ileal and fecal microbiota and metabolite profiles to antibiotics in cannulated pigs. Appl. Microbiol. Biotechnol. 2018, 102, 2289–2299. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537. [Google Scholar] [CrossRef]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. Isme J. 2011, 5, 169–172. [Google Scholar] [CrossRef]
- Zhang, C.; Miao, Y.; Yang, Y.; Mu, C.; Yong, S.; Zhu, W. Differential effect of early antibiotic intervention on bacterial fermentation patterns and mucosal gene expression in the colon of pigs under diets with different protein levels. Appl. Microbiol. Biotechnol. 2016, 101, 1–13. [Google Scholar] [CrossRef]
- Kritas, S.K.; Pensaert, M.B.; Mettenleiter, T.C. Invasion and spread of single glycoprotein deleted mutants of Aujeszky’s disease virus (ADV) in the trigeminal nervous pathway of pigs after intranasal inoculation. Vet. Microbiol. 1994, 40, 323–334. [Google Scholar] [CrossRef]
- Yang, Q.Y.; Sun, Z.; Tan, F.F.; Guo, L.H.; Wang, Y.Z.; Wang, J.; Wang, Z.Y.; Wang, L.L.; Li, X.D.; Xiao, Y. Pathogenicity of a currently circulating Chinese variant pseudorabies virus in pigs. World J. Virol. 2016, 5, 23. [Google Scholar] [CrossRef]
- Praveena, P.E.; Periasamy, S.; Kumar, A.A.; Singh, N. Cytokine profiles, apoptosis and pathology of experimental Pasteurella multocida serotype A1 infection in mice. Res. Vet. Sci. 2010, 89, 332–339. [Google Scholar] [CrossRef]
- Boudry, G.; Péron, V.; HuërouLuron, I.L.; Lallès, J.P.; Sève, B. Weaning induces both transient and long-lasting modifications of absorptive, secretory, and barrier properties of piglet intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef]
- Phalipon, A.; Cardona, A.; Kraehenbuhl, J.P.; Edelman, L.; Sansonetti, P.J.; Corthésy, B. Secretory component: A new role in secretory IgA-mediated immune exclusion in vivo. Immunity 2002, 17, 107–115. [Google Scholar] [CrossRef]
- Jonefjäll, B.; Öhman, L.; Simrén, M.; Strid, H. IBS-like symptoms in patients with ulcerative colitis in deep remission are associated with increased levels of serum cytokines and poor psychological well-being. Inflamm. Bowel Dis. 2016, 22, 2630. [Google Scholar]
- Jian, W.; Fengqi, L.; Haiming, W.; Zhe-Xiong, L.; Rui, S.; Zhigang, T. Respiratory influenza virus infection induces intestinal immune injury via microbiota-mediated Th17 cell-dependent inflammation. J. Exp. Med. 2014, 211, 2397–2410. [Google Scholar]
- Zhao, N.; Wang, S.; Li, H.; Liu, S.; Li, M.; Luo, J.; Su, W.; He, H. Influence of novel highly pathogenic avian influenza A (H5N1) virus infection on migrating whooper swans fecal microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 46. [Google Scholar] [CrossRef]
- Palm, N.W.; de Zoete, M.R.; Flavell, R.A. Immune-microbiota interactions in health and disease. Clin. Immunol. 2015, 159, 122–127. [Google Scholar] [CrossRef]
- Kawase, M.; He, F.; Kubota, A.; Yoda, K.; Miyazawa, K.; Hiramatsu, M. Heat-killed Lactobacillus gasseri TMC0356 protects mice against influenza virus infection by stimulating gut and respiratory immune responses. Pathog. Dis. 2012, 64, 280–288. [Google Scholar] [CrossRef]
- Aiba, Y.; Nakano, Y.; Koga, Y.; Takahashi, K.; Komatsu, Y. A highly acid-resistant novel strain of Lactobacillus johnsonii No. 1088 has antibacterial activity, including that against Helicobacter pylori, and inhibits gastrin-mediated acid production in mice. Microbiologyopen 2015, 4, 465–474. [Google Scholar] [CrossRef]
- Hou, C.; Zeng, X.; Yang, F.; Liu, H.; Qiao, S. Study and use of the probiotic Lactobacillus reuteri in pigs: A review. J. Anim. Sci. Biotechnol. 2016, 6, 14. [Google Scholar] [CrossRef]
- Tamanaishacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnauremallet, M.; Jolivetgougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Flanagan, L.; Schmid, J.; Ebert, M.; Soucek, P.; Kunicka, T.; Liska, V.; Bruha, J.; Neary, P.; Dezeeuw, N.; Tommasino, M. Fusobacterium nucleatum associates with stages of colorectal neoplasia development, colorectal cancer and disease outcome. Eur. J. Clin. Microbiol. 2014, 33, 1381–1390. [Google Scholar] [CrossRef]
- Ryan, M.P.; Adley, C.C. Sphingomonas paucimobilis: A persistent Gram-negative nosocomial infectious organism. J. Hosp. Infect. 2010, 75, 153–157. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharm. Therap. 2007, 27, 104–119. [Google Scholar] [CrossRef]
- Rhee, S.H. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat. Rev. Gastro. Hepat. 2009, 6, 306–314. [Google Scholar] [CrossRef]
- Behnsen, J.; Jellbauer, S.; Wong, C.P.; Edwards, R.A.; George, M.D.; Ouyang, W.; Raffatellu, M. The cytokine IL-22 promotes pathogen colonization by suppressing related commensal bacteria. Immunity 2014, 40, 262–273. [Google Scholar] [CrossRef]
- Elisa, D.; Boxx, G.M.; He, X.; Pan, C.; David, B.S.; Cen, L.; Nora, R.; Shi, W.; Cheng, G. Influenza virus affects intestinal microbiota and secondary Salmonella infection in the gut through type I interferons. PLoS Pathog. 2016, 12, e1005572. [Google Scholar]
- Kato, L.M.; Kawamoto, S.; Maruya, M.; Fagarasan, S. The role of the adaptive immune system in regulation of gut microbiota. Immunol. Rev. 2014, 260, 67–75. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Liu, Y.; Chen, S.; Qiao, Y.; Zheng, Y.; Xu, M.; Wang, Z.; Hou, J.; Wang, J.; Fan, H. Effects of Intranasal Pseudorabies Virus AH02LA Infection on Microbial Community and Immune Status in the Ileum and Colon of Piglets. Viruses 2019, 11, 518. https://doi.org/10.3390/v11060518
Zhang C, Liu Y, Chen S, Qiao Y, Zheng Y, Xu M, Wang Z, Hou J, Wang J, Fan H. Effects of Intranasal Pseudorabies Virus AH02LA Infection on Microbial Community and Immune Status in the Ileum and Colon of Piglets. Viruses. 2019; 11(6):518. https://doi.org/10.3390/v11060518
Chicago/Turabian StyleZhang, Chuanjian, Yamei Liu, Saisai Chen, Yongfeng Qiao, Yating Zheng, Mengwei Xu, Zhisheng Wang, Jibo Hou, Jichun Wang, and Hongjie Fan. 2019. "Effects of Intranasal Pseudorabies Virus AH02LA Infection on Microbial Community and Immune Status in the Ileum and Colon of Piglets" Viruses 11, no. 6: 518. https://doi.org/10.3390/v11060518
APA StyleZhang, C., Liu, Y., Chen, S., Qiao, Y., Zheng, Y., Xu, M., Wang, Z., Hou, J., Wang, J., & Fan, H. (2019). Effects of Intranasal Pseudorabies Virus AH02LA Infection on Microbial Community and Immune Status in the Ileum and Colon of Piglets. Viruses, 11(6), 518. https://doi.org/10.3390/v11060518